
Olive Leaf as a Source of Antibacterial Compounds Active against Antibiotic-Resistant Strains of Campylobacter jejuni and Campylobacter coli

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Antibiotic Susceptibility of Campylobacter Strains
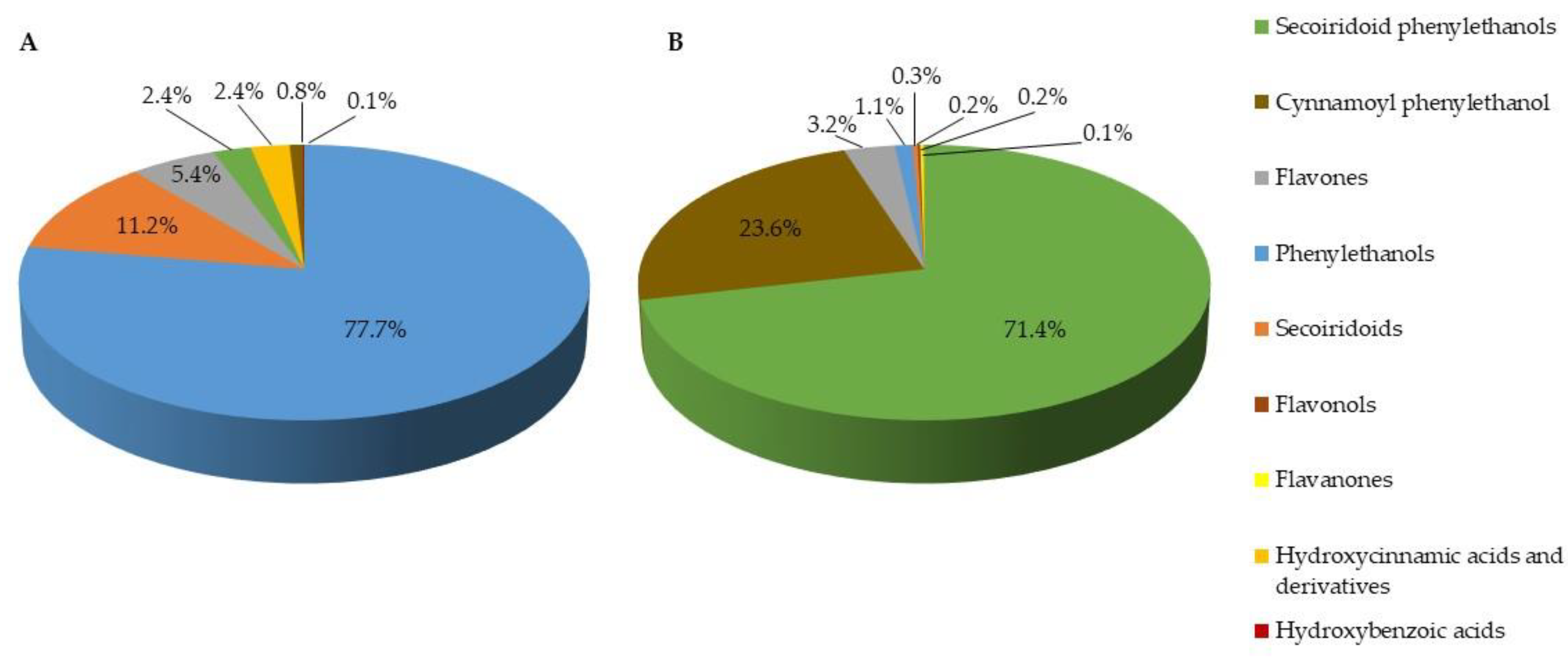
2.2. Chemical Characterization of Olive Leaf Extracts
2.3. Antibacterial Activity of Olive-Leaf Extracts against Campylobacter Strains
3. Materials and Methods
3.1. Olive Leaf Extracts, Reagents, and Reference Substances
3.2. Chemical Characterization
3.3. Campylobacter Strains, Growth Media, and Culture Conditions
3.4. Determination of Antibiotic Susceptibility of Campylobacter spp. Strains
3.5. Determination of Antibacterial Activity of Olive-Leaf Extracts against Campylobacter spp. Strains
3.6. Verification of Antibacterial Activity Using Pure Hydroxytyrosol
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union one health 2020 zoonoses report. EFSA J. 2021, 19, 6971. [Google Scholar]
- Silvan, J.M.; Martinez-Rodriguez, A.J. Food by-products as natural source of bioactive compounds against Campylobacter. In Encyclopedia of Food Security and Sustainability; Ferranti, P., Berry, E.M., Anderson, J.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 3, pp. 336–350. [Google Scholar]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human campylobacteriosis—A serious infectious threat in a one health perspective. In Fighting Campylobacter Infections. Current Topics in Microbiology and Immunology; Backert, S., Ed.; Springer: Cham, Switzerland, 2021; Volume 431, pp. 1–23. [Google Scholar]
- Wakerley, B.R.; Uncini, A.; Yuki, N.; Attarian, S.; Barreira, A.A.; Chan, Y.-C.; Créange, A.; Hsieh, S.-T.; Islam, B.; Kannan, M.A. Guillain-Barré and Miller Fisher syndromes—New diagnostic classification. Nat. Rev. Neurol. 2014, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- Ben Romdhane, R.; Merle, R. The data behind risk analysis of Campylobacter jejuni and Campylobacter coli infections. In Fighting Campylobacter Infections. Current Topics in Microbiology and Immunology; Backert, S., Ed.; Springer: Cham, Switzerland, 2021; Volume 431, pp. 25–58. [Google Scholar]
- Hakeem, M.J.; Lu, X. Survival and control of Campylobacter in poultry production environment. Front. Cell Infect. Microbiol. 2021, 10, 615049. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and potential treatments for reducing Campylobacter colonization in animal hosts and disease in humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union Summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar]
- Gahamanyi, N.; Song, D.G.; Cha, K.H.; Yoon, K.Y.; Mboera, L.E.G.; Matee, M.I.; Mutangana, D.; Amachawadi, R.G.; Komba, E.V.G.; Pan, C.H. Susceptibility of Campylobacter strains to selected natural products and frontline antibiotics. Antibiotics 2020, 9, 790. [Google Scholar] [CrossRef]
- Marotta, F.; Garofolo, G.; di Marcantonio, L.; Di Serafino, G.; Neri, D.; Romantini, R.; Sacchini, L.; Alessiani, A.; Di Donato, G.; Nuvoloni, R.; et al. Antimicrobial resistance genotypes and phenotypes of Campylobacter jejuni isolated in Italy from humans, birds from wild and urban habitats, and poultry. PLoS ONE 2019, 14, e0223804. [Google Scholar] [CrossRef] [Green Version]
- Klančnik, A.; Možina, S.S.; Zhang, Q. Anti-Campylobacter activities and resistance mechanisms of natural phenolic compounds in Campylobacter. PLoS ONE 2012, 7, e51800. [Google Scholar] [CrossRef] [Green Version]
- Silvan, J.M.; Mingo, E.; Hidalgo, M.; de Pascual-Teresa, S.; Carrascosa, A.V.; Martínez-Rodríguez, A.J. Antibacterial activity of a grape seed extract and its fractions against Campylobacter spp. Food Control 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Mingo, E.; Silvan, J.M.; Martínez-Rodríguez, A.J. Selective antibacterial effect on Campylobacter of a winemaking waste extract (WWE) as a source of active phenolic compounds. LWT Food Sci. Technol. 2016, 68, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Silvan, J.M.; Pinto-Bustillos, M.A.; Vasquez-Ponce, P.; Prodanov, M.; Martinez-Rodriguez, A.J. Olive mill wastewater as a potential source of antibacterial and anti-inflammatory compounds against the food-borne pathogen Campylobacter. Innov. Food Sci. Emerg. Technol. 2019, 51, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Možina, S.S.; Klančnik, A.; Kovac, J.; Jeršek, B.; Bucar, F. Antimicrobial natural products against Campylobacter. In Natural Antimicrobial Agents. Sustainable Development and Biodiversity; Mérillon, J.M., Riviere, C., Eds.; Springer: Cham, Switzerland, 2018; Volume 19, pp. 3–30. [Google Scholar]
- EFSA (European Food Safety Authority). Scientific opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 2105. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Update and review of control options for Campylobacter in broilers at primary production. EFSA J. 2020, 18, e06090. [Google Scholar]
- Baptista, E.; Borges, A.; Aymerich, T.; Alves, S.P.; Gama, L.T.d.; Fernandes, H.; Fernandes, M.J.; Fraqueza, M.J. Pulsed light application for Campylobacter control on poultry meat and its effect on colour and volatile profile. Foods 2022, 11, 2848. [Google Scholar] [CrossRef]
- Moudache, M.; Silva, F.; Nerín, C.; Zaidi, F. Olive cake and leaf extracts as valuable sources of antioxidant and antimicrobial compounds: A comparative study. Waste Biomass Valorization 2021, 12, 1431–1445. [Google Scholar] [CrossRef]
- Branciari, R.; Ranucci, D.; Ortenzi, R.; Roila, R.; Trabalza-Marinucci, M.; Servili, M.; Papa, P.; Galarini, R.; Valiani, A. Dietary administration of olive mill wastewater extract reduces Campylobacter spp. prevalence in broiler chickens. Sustainability 2016, 8, 837. [Google Scholar] [CrossRef] [Green Version]
- Romero-García, J.M.; Niño, L.; Martínez-Patiño, C.; Álvarez, C.; Castro, E.; Negro, M.J. Biorefinery based on olive biomass. State of the art and future trends. Bioresour. Technol. 2014, 159, 421–432. [Google Scholar] [CrossRef]
- Sudjana, A.N.; D’Orazio, C.; Ryan, V.; Rasool, N.; Ng, J.; Islam, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of commercial Olea europaea (olive) leaf extract. Int. J. Antimicrob. Agents 2009, 33, 461–463. [Google Scholar] [CrossRef]
- Šimat, V.; Skroza, D.; Tabanelli, G.; Čagalj, M.; Pasini, F.; Gómez-Caravaca, A.M.; Fernández-Fernández, C.; Sterniša, M.; Možina, S.; Ozogul, Y.; et al. Antioxidant and antimicrobial activity of hydroethanolic leaf extracts from six mediterranean olive cultivars. Antioxidants 2022, 11, 1656. [Google Scholar] [CrossRef]
- Dunai, A.; Spohn, R.; Farkas, Z.; Lázár, V.; Györkei, Á.; Apjok, G.; Boross, G.; Szappanos, B.; Grézal, G.; Faragó, A.; et al. Rapid decline of bacterial drug-resistance in an antibiotic-free environment through phenotypic reversion. Elife 2019, 8, e47088. [Google Scholar] [CrossRef]
- Sproston, E.L.; Wimalarathna, H.M.L.; Sheppard, S.K. Trends in fluoroquinolone resistance in Campylobacter. Microb. Genom. 2018, 4, e000198. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhang, M.; Zhou, J.; Pang, L.; Wang, G.; Hou, F. The molecular mechanisms of ciprofloxacin resistance in clinical Campylobacter jejuni and their genotyping characteristics in Beijing, China. Foodborne Pathog. Dis. 2017, 14, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Kovač, J.; Čadež, N.; Stessl, B.; Stingl, K.; Gruntar, I.; Ocepek, M.; Trkov, M.; Wagner, M.; Smole Možina, S. High genetic similarity of ciprofloxacin-resistant Campylobacter jejuni in central Europe. Front. Microbiol. 2015, 6, 1169. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.E.; Barton, M.D.; Blair, I.S.; Corcoran, D.; Dooley, J.S.; Fanning, S.; Kempf, I.; Lastovica, A.J.; Lowery, C.J.; Matsuda, M.; et al. The epidemiology of antibiotic resistance in Campylobacter. Microbes Infect. 2006, 8, 1955–1966. [Google Scholar] [CrossRef]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Kielsznia, A.; Korzekwa, K.; Tobiasz, A.; Korzeniowska-Kowal, A.; Wieliczko, A. High prevalence of resistance to fluoroquinolones and tetracycline Campylobacter spp. isolated from poultry in Poland. Microb. Drug Resist. 2018, 24, 314–322. [Google Scholar] [CrossRef]
- Metreveli, M.; Bulia, S.; Tevzadze, L.; Tsanava, S.; Zarske, M.; Goenaga, J.C.; Preuß, S.; Lomidze, G.; Koulouris, S.; Imnadze, P.; et al. Comparison of antimicrobial susceptibility profiles of thermotolerant Campylobacter spp. isolated from human and poultry samples in Georgia (Caucasus). Antibiotics 2022, 11, 1419. [Google Scholar] [CrossRef] [PubMed]
- CDC (Center for Disease Control and Prevention). Antibiotic resistance threats in the United States, 2019. In AR Threats Report; Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019; pp. 79–80. [Google Scholar]
- Wang, J.; Wang, Z.; Zhang, J.M.; Ding, Y.; Ma, Z.H.; Jiang, F.F.; Nie, X.; Tang, S.J.; Chen, M.T.; Wu, S.; et al. Prevalence, antibiotic susceptibility and genetic diversity of Campylobacter jejuni isolated from retail food in China. LWT Food Sci. Technol. 2021, 143, 111098. [Google Scholar] [CrossRef]
- Pergola, S.; Franciosini, M.P.; Comitini, F.; Ciani, M.; De Luca, S.; Bellucci, S.; Menchetti, L.; Casagrande Proietti, P. Genetic diversity and antimicrobial resistance profiles of Campylobacter coli and Campylobacter jejuni isolated from broiler chicken in farms and at time of slaughter in central Italy. J. Appl. Microbiol. 2017, 122, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Post, A.; Martiny, D.; van Waterschoot, N.; Hallin, M.; Maniewski, U.; Bottieau, E.; Van Esbroeck, M.; Vlieghe, E.; Ombelet, S.; Vandenberg, O.; et al. Antibiotic susceptibility profiles among Campylobacter isolates obtained from international travelers between 2007 and 2014. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2101–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, F.; Colston, J.M.; Paredes-Olortegui, M.; François, R.; Pisanic, N.; Burga, R.; Peñataro-Yori, P.; Kosek, M.N. Antibiotic resistance of Campylobacter species in a pediatric cohort study. Antimicrob. Agents Chemother. 2019, 63, e01911–e01918. [Google Scholar] [CrossRef] [Green Version]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, N.S.A.; Bradford, J.M. Recovery and stability of oleuropein and other phenolic compounds during extraction and processing of olive (Olea europaea L.) leaves. J. Food Agric. Environ. 2008, 6, 8–13. [Google Scholar]
- Medina, E.; Romero, C.; García, P.; Brenes, M. Characterization of bioactive compounds in commercial olive leaf extracts, and olive leaves and their infusions. Food Funct. 2019, 10, 4716–4724. [Google Scholar] [CrossRef]
- Erbay, Z.; Icier, F. The importance and potential uses of olive leaves. Food Rev. Int. 2010, 26, 319–334. [Google Scholar] [CrossRef]
- Borjan, D.; Leitgeb, M.; Knez, Ž.; Hrnčič, M.K. Microbiological and antioxidant activity of phenolic compounds in olive leaf extract. Molecules 2020, 25, 5946. [Google Scholar] [CrossRef]
- Chatzikonstantinou, A.V.; Giannakopoulou, A.; Spyrou, S.; Simos, Y.V.; Kontogianni, V.G.; Peschos, D.; Katapodis, P.; Polydera, A.C.; Stamatis, H. Production of hydroxytyrosol rich extract from Olea europaea leaf with enhanced biological activity using immobilized enzyme reactors. Environ. Sci. Pollut. Res. Int. 2022, 29, 29624–29637. [Google Scholar] [CrossRef]
- Wei, J.; Wang, S.; Pei, D.; Qu, L.; Li, Y.; Chen, J.; Di, D.; Gao, K. Antibacterial activity of hydroxytyrosol acetate from olive leaves (Olea Europaea L.). Nat. Prod. Res. 2018, 32, 1967–1970. [Google Scholar] [CrossRef]
- Belaqziz, M.; Tan, S.P.; El-Abbassi, A.; Kiai, H.; Hafidi, A.; O’Donovan, O.; McLoughlin, P. Assessment of the antioxidant and antibacterial activities of different olive processing wastewaters. PLoS ONE 2017, 12, e0182622. [Google Scholar] [CrossRef] [Green Version]
- Yakhlef, W.; Arhab, R.; Romero, C.; Brenes, M.; de Castro, A.; Medina, E. Phenolic composition and antimicrobial activity of Algerian olive products and by-products. LWT Food Sci. Technol. 2018, 93, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Brenes, M.; García, A.; de los Santos, B.; Medina, E.; Romero, C.; de Castro, A.; Romero, F. Olive glutaraldehyde-like compounds against plant pathogenic bacteria and fungi. Food Chem. 2011, 125, 1262–1266. [Google Scholar] [CrossRef]
- Medina, E.; Brenes, M.; García, A.; Romero, C.; de Castro, A. Bactericidal activity of glutaraldehyde-like compounds from olive products. J. Food Prot. 2009, 72, 2611–2614. [Google Scholar] [CrossRef]
- Medina-Martínez, M.S.; Truchado, P.; Castro-Ibáñez, I.; Allende, A. Antimicrobial activity of hydroxytyrosol: A current controversy. Biosci. Biotechnol. Biochem. 2016, 80, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.J.; Tu, J.L.; Qin, F.G.F.; Xu, Y.J.; Li, B. Phenolic composition of oleuropein extract after enzymatic process by HPLC-MS and their antioxidant and antibacterial activities. J. Food Biochem. 2018, 42, e12517. [Google Scholar] [CrossRef]
- Musella, E.; Ouazzani, I.C.e.; Mendes, A.R.; Rovera, C.; Farris, S.; Mena, C.; Teixeira, P.; Poças, F. Preparation and characterization of bioactive chitosan-based films incorporated with olive leaves extract for food packaging applications. Coatings 2021, 11, 1339. [Google Scholar] [CrossRef]
- Silvan, J.M.; Guerrero-Hurtado, E.; Gutiérrez-Docio, A.; Alarcon-Cavero, T.; Prodanov, M.; Martinez-Rodriguez, A.J. Olive leaf extracts modulate inflammation and oxidative stress associated with human H. pylori infection. Antioxidants 2021, 10, 2030. [Google Scholar] [CrossRef]
- Medina, E.; Brenes, M.; Romero, C.; García, A.; de Castro, A. Main antimicrobial compounds in table olives. J. Agric. Food Chem. 2007, 55, 9817–9823. [Google Scholar] [CrossRef]
- Perestrelo, R.; Lu, Y.; Santos, S.A.O.; Silvestre, A.J.D.; Neto, C.P.; Câmara, J.S.; Rocha, S.M. Phenolic profile of sercial and tinta negra Vitis vinifera L. grape skins by HPLC–DAD–ESI-MSn: Novel phenolic compounds in Vitis vinifera L. grape. Food Chem. 2012, 135, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Michel, T.; Khlif, I.; Kanakis, P.; Termentzi, A.; Allouche, N.; Halabalaki, M.; Skaltsounis, A.L. UHPLC-DAD-FLD and UHPLCHRMS/MS based metabolic profiling and characterization of different Olea europaea organs of Koroneiki and Chetoui varieties. Phytochem. Let. 2015, 11, 424–439. [Google Scholar] [CrossRef]
- García-Sánchez, L.; Melero, B.; Jaime, I.; Hänninen, M.L.; Rossi, M.; Rovira, J. Campylobacter jejuni survival in a poultry processing plant environment. Food Microbiol. 2017, 65, 185–192. [Google Scholar] [CrossRef]
- Persson, S.; Olsen, K.E.P. Multiplex PCR for identification of Campylobacter coli and Campylobacter jejuni from pure cultures and directly on stool samples. J. Med. Microbiol. 2005, 54, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.; Gatica, M.A.; Riquelme, V.; Vergara, C.; Yañez, J.M.; San Martín, B.; Sáenz, L.; Vidal, M.; Martínez, M.C.; Araya, P.; et al. Characterization of antimicrobial susceptibility and its association with virulence genes related to adherence, invasion, and cytotoxicity in Campylobacter jejuni and Campylobacter coli isolates from animals, meat, and humans. Microb. Drug Resist. 2016, 22, 432–444. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). EU Protocol for Harmonised Monitoring of Antimicrobial Resistance in Human Salmonella and Campylobacter Isolates—June 2016; ECDC: Stockholm, Sweeden, 2016. [Google Scholar]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 10 October 2022).
- Société Française de Microbiologie (SFM). Tableaux des concentrations critiques pour l’interpretation des CMI et des diamètres critiques des zones d’inhibition. Campylobacter spp. In CA-SFM/EUCAST; Société Française de Microbiologie: Paris, France, 2022; pp. 119–120. [Google Scholar]
- Villalva, M.; Silvan, J.M.; Guerrero-Hurtado, E.; Gutierrez-Docio, A.; Navarro del Hierro, J.; Alarcón-Cavero, T.; Prodanov, M.; Martin, D.; Martinez-Rodriguez, A.J. Influence of in vitro gastric digestion of olive leaf extracts on their bioactive properties against H. pylori. Foods 2022, 11, 1832. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef]


{kind=link}
| Specie | Strain | CIP | TET | ERY | GEN | AMC | AMX | Antibiotic Resistant Rate |
|---|---|---|---|---|---|---|---|---|
| C. jejuni | JS1 | R (>32) | R (32) | S (0.5) | S (0.25) | S (0.5) | I (16) | 2/6 |
| JR1 | R (>32) | R (32) | S (0.5) | S (0.38) | S (0.5) | R (32) | 3/6 | |
| JR2 | R (>32) | R (>256) | S (0.5) | S (0.19) | S (0.38) | R (>256) | 3/6 | |
| JP1 | R (>32) | R (24) | S (0.38) | S (0.125) | S (0.25) | R (24) | 3/6 | |
| 700819 | I (0.032) | S (0.032) | S (0.25) | S (0.5) | S (0.19) | S (3) | 0/6 | |
| C. coli | CR1 | R (>32) | R (32) | S (0.75) | S (0.75) | S (1) | S (3) | 2/6 |
| CP1 | R (>32) | R (>256) | S (1) | S (0.5) | S (1) | I (6) | 2/6 | |
| CR2 | R (>32) | R (3) | S (0.5) | S (0.5) | S (0.5) | S (2) | 2/6 | |
| CP2 | R (>32) | R (>256) | S (2) | S (0.5) | S (0.75) | I (8) | 2/6 | |
| 43478 | I (0.016) | S (0.032) | S (0.125) | S (0.75) | S (2) | I (8) | 0/6 | |
| Strain resistance rate | 8/10 | 8/10 | 0/10 | 0/10 | 0/10 | 3/10 | ||
| Compounds | Extract E1 | Extract E2 |
|---|---|---|
| Phenylethanols | ||
| 3,4-DHPE (Hydroxytyrosol) + 3,4-DHPE glucoside 1 | 13,743 ± 1659 * | 182 ± 4 * |
| 3,4-DHPE glucoside 2 + 3 | 965 ± 13 * | 123 ± 1 * |
| 4-HPE (Tyrosol) | 250 ± 6 * | 9.1 ± 0.1 * |
| 3,4-DHPG | 20.1 ± 0.4 * | 9.4 ± 0.5 * |
| Secoiridoids | ||
| EA 2-glucoside (Oleoside 11-methyl ester) | 1352 ± 49 * | 84.4 ± 4.6 * |
| EMA 2-glucoside (Secoxyloganin) | 657 ± 158 | ND |
| EA (Elenolic acid) | 155 ± 14 | ND |
| Flavones | ||
| Luteolin 7-O-glucoside | 655 ± 22 * | 513 ± 46 * |
| Luteolin 4′-methyl ether 7-O-glucoside (Diosmin) | 123 ± 11 | 111 ± 12 |
| Apigenin 7-O-rutinoside (Isorhoifolin) | 109 ± 1 * | 122 ± 5 * |
| Apigenin 7-O-glucuronide | 76.4 ± 7.1 | 64.0 ± 6.0 |
| Luteolin 3′,7-di-O-glucoside | 39.9 ± 1.4 * | 69.6 ± 2.3 * |
| Apigenin 6,8-di-C-glucoside | 39.3 ± 1.4 * | 24.2 ± 0.2 * |
| Luteolin | ND | 17.1 ± 1.2 |
| Secoiridoid phenylethanols | ||
| 3,4-DHPE-EA glucoside (Oleuropein) | 355 ± 57 * | 20,471 ± 1061 * |
| 4-HPE-EA-glucoside (Ligustroside) | 99.3 ± 9.3 * | 360 ± 16 * |
| Hydroxycinnamic acids and derivatives | ||
| trans-4-HCA (trans-4-coumaric acid) | 209 ± 45 * | 1.2 ± 0.1 * |
| trans-3,4-DHCA (trans-caffeic acid) | 140 ± 4 * | 4.5 ± 0.1 * |
| trans-3-M,4-HCA (trans-ferulic acid) | 116 ± 4 * | 5.0 ± 0.5 * |
| trans-4,5-DCQA (trans-4,5-dicaffeoylquinic acid) | ND | 16.5 ± 0.2 |
| Cynnamoyl phenylethanol | ||
| 3,4-DHPE caffeoyl glucoside (Verbascoside) | 161 ± 11 * | 6872 ± 230 * |
| Hydroxybenzoic acids | ||
| 3,4-DHBA (Protocatechuic acid) | 7.9 ± 0.2 | ND |
| 3,4-DHBA glucoside | 6.4 ± 0.7 | ND |
| Flavonols | ||
| Quercetin 3-O-glucoside (Isoquercitrin) | ND | 9.1 ± 0.6 |
| Quercetin 3-rhamnoside (Quercitrin) | ND | 10.8 ± 0.2 |
| Quercetin | ND | 31.4 ± 0.5 |
| Flavanones | ||
| Eriodictyol 7-O-rutinoside | ND | 22.3 ± 3.1 |
| Eriodictyol 7-O-glucoside | ND | 22.8 ± 1.0 |
| Total phenolic and secoiridoid compounds | 19,279 | 29,155 |
| Species | Strains | Control Growth | Extract E1 | Extract E2 | ||||
|---|---|---|---|---|---|---|---|---|
| log CFU/mL | log Reduction | MIC (mg/mL) | log CFU/mL | log Reduction | MIC (mg/mL) | |||
| C. jejuni | JS1 | 9.62 ± 0.04 c | <1.48 a | >8.14 | 1.0 | 8.13 ± 0.09 b | 1.49 | 2.0 |
| JR1 | 8.63 ± 0.03 c | <1.48 a | >7.15 | 1.0 | 6.98 ± 0.09 b | 1.65 | 2.0 | |
| JR2 | 8.50 ± 0.06 c | 2.67 ± 0.14 a | 5.83 | 2.0 | 6.86 ± 0.10 b | 1.64 | 2.0 | |
| JP1 | 9.31 ± 0.06 c | 3.99 ± 0.04 a | 5.32 | 2.0 | 8.08 ± 0.04 b | 1.23 | 2.0 | |
| 700819 | 9.34 ± 0.05 c | 5.22 ± 0.04 a | 4.12 | 2.0 | 8.82 ± 0.01 b | 0.52 | 2.0 | |
| C. coli | CR1 | 8.64 ± 0.03 c | <1.48 a | >7.16 | 0.1 | 6.69 ± 0.02 b | 1.95 | 2.0 |
| CP1 | 9.32 ± 0.05 c | <1.48 a | >7.84 | 0.5 | 6.83 ± 0.06 b | 2.49 | 2.0 | |
| CR2 | 9.47 ± 0.04 b | <1.48 a | >7.99 | 0.1 | 9.78 ± 0.03 b | - | - | |
| CP2 | 9.16 ± 0.09 c | <1.48 a | >7.68 | 1.5 | 8.01 ± 0.07 b | 1.15 | 2.0 | |
| 43478 | 8.47 ± 0.05 c | <1.48 a | >6.99 | 0.1 | 6.22 ± 0.05 b | 2.25 | 2.0 | |
| Strains | Control Growth | Hydroxytyrosol | log Reduction | MIC (mg/mL) |
|---|---|---|---|---|
| C. jejuni JS1 | 9.49 ± 0.05 | <1.48 | >9.49 | 0.25 |
| C. coli CR1 | 8.76 ± 0.03 | <1.48 | >8.76 | 0.1 |
| Strain Designation | Specie | Isolation Source |
|---|---|---|
| JS1 | C. jejuni | Slaughterhouse |
| JR1 | C. jejuni | Chicken breast retail |
| JR2 | C. jejuni | Chicken drumstick retail |
| JP1 | C. jejuni | Carcass in production chain |
| 700819 | C. jejuni | Reference strain * |
| CR1 | C. coli | Chicken drumstick retail |
| CP1 | C. coli | Machine in production chain |
| CR2 | C. coli | Hamburger retail |
| CP2 | C. coli | Carcass in production chain |
| 43478 | C. coli | Reference strain * |
| Antibiotic Group | Antibiotic Agent | Concentration Range (µg/mL) | Breakpoint (S/R) MIC (µg/mL) | |
|---|---|---|---|---|
| C. jejuni | C. coli | |||
| Macrolides | Erythromycin (ERY) | 0.016–256 | ≤4/>4 1 | ≤8/>8 1 |
| Tetracyclines | Tetracycline (TET) | 0.016–256 | ≤2/>2 1 | ≤2/>2 1 |
| Aminoglycosides | Gentamicin (GEN) | 0.016–256 | ≤2/>2 2 | ≤2/>2 2 |
| Penicillins | Amoxicillin (AMX) | 0.016–256 | ≤4/>16 2 | ≤4/>16 2 |
| Amoxicillin/Clavulanic acid (AMC) | 0.016–256 | ≤4/>16 2 | ≤4/>16 2 | |
| Fluoroquinolones | Ciprofloxacin (CIP) | 0.002–32 | ≤0.001/>0.5 1 | ≤0.001/>0.5 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvan, J.M.; Guerrero-Hurtado, E.; Gutierrez-Docio, A.; Prodanov, M.; Martinez-Rodriguez, A.J. Olive Leaf as a Source of Antibacterial Compounds Active against Antibiotic-Resistant Strains of Campylobacter jejuni and Campylobacter coli. Antibiotics 2023, 12, 26. https://doi.org/10.3390/antibiotics12010026
Silvan JM, Guerrero-Hurtado E, Gutierrez-Docio A, Prodanov M, Martinez-Rodriguez AJ. Olive Leaf as a Source of Antibacterial Compounds Active against Antibiotic-Resistant Strains of Campylobacter jejuni and Campylobacter coli. Antibiotics. 2023; 12(1):26. https://doi.org/10.3390/antibiotics12010026
Chicago/Turabian StyleSilvan, Jose Manuel, Esperanza Guerrero-Hurtado, Alba Gutierrez-Docio, Marin Prodanov, and Adolfo J. Martinez-Rodriguez. 2023. "Olive Leaf as a Source of Antibacterial Compounds Active against Antibiotic-Resistant Strains of Campylobacter jejuni and Campylobacter coli" Antibiotics 12, no. 1: 26. https://doi.org/10.3390/antibiotics12010026
APA StyleSilvan, J. M., Guerrero-Hurtado, E., Gutierrez-Docio, A., Prodanov, M., & Martinez-Rodriguez, A. J. (2023). Olive Leaf as a Source of Antibacterial Compounds Active against Antibiotic-Resistant Strains of Campylobacter jejuni and Campylobacter coli. Antibiotics, 12(1), 26. https://doi.org/10.3390/antibiotics12010026