
Targeting Enterococci with Antimicrobial Activity against Clostridium perfringens from Poultry


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Enterococcus Sampling and Identification
2.2. Screening of Enterococci for Antimicrobial, Specifically Anti-C. perfringens Activity
2.3. Effects of the Supernatants of BP+ Enterococci on C. perfringens Isolates
2.4. Phenotypic and Genotypic Characterization of the Selected BP+ Enterococci
2.4.1. Bacteriocinome
2.4.2. Antibiotic Resistance phenotype and resistome
2.4.3. Virulence
- Gelatinase activity and hemolysis
- Virulome
2.4.4. Plasmidome
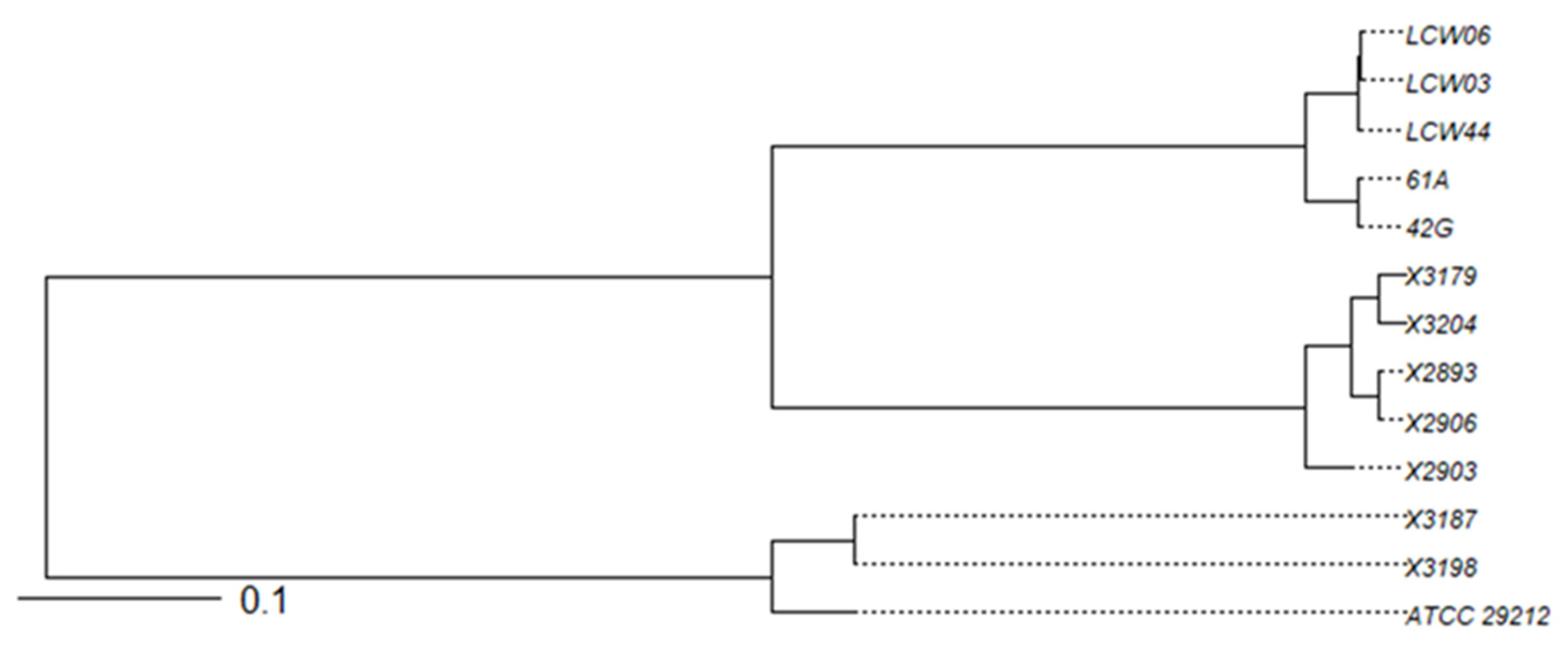
2.4.5. Genetic Lineages
3. Discussion
3.1. Screening for BP+ Enterococci
3.2. Phenotypic and Genotypic Characteristics of the BP+ Enterococci
3.2.1. Bacteriocinome
3.2.2. BP+ Enterococcus Resistance Phenotype and Resistome
3.2.3. Virulence of BP+ Enterococci
3.2.4. Plasmidome of the BP+ Enterococci
4. Materials and Methods
4.1. Enterococcus Sampling and Identification
4.2. Screening for Anti-C. perfringens Activity Using the “Spot on the Lawn” Method
4.3. Screening for Anti-C. perfringens Activity Using the Agar Diffusion Method
4.4. Anti-C. perfringens Activity Determination Using Microtitration Assay
4.5. Characterization of BP+ Enterococci
4.5.1. Susceptibility to Antibiotics
4.5.2. Gelatinase Activity and Hemolysis
4.5.3. Whole Genome Sequencing (WGS) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Wang, F.; Wang, Y.; Wan, Z.; Shao, H.; Qian, K.; Ye, J.; Qin, A. Generation of a recombinant chickenized monoclonal antibody against the neuraminidase of H9N2 avian influenza virus. AMB Express 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Gochez, D.; Moulin, G.; Erlacher-Vindel, E. OIE Annual Report on Antimicrobial Agents Intended for Use in Animals: Methos Used. Front. Vet. Sci. 2021, 6, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Mora, Z.V.-d.l.; Macías-Rodríguez, M.E.; Arratia-Quijada, J.; Gonzalez-Torres, Y.S.; Nuño, K.; Villarruel-López, A. Clostridium perfringens as foodborne pathogen in broiler production: Pathophysiology and potential strategies for controlling necrotic enteritis. Animals 2020, 10, 1718. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Shojadoost, B.; Boodhoo, N.; Astill, J.; Taha-Abdelaziz, K.; Hodgins, D.C.; Kulkarni, R.R.; Sharif, S. Necrotic enteritis in chickens: A review of pathogenesis, immune responses and prevention, focusing on probiotics and vaccination. Anim. health Res. Rev. 2021, 22, 147–162. [Google Scholar] [CrossRef]
- Immerseel, F.V.; Buck, J.D.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian. Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef]
- Agunos, A.; Pierson, F.W.; Lungu, B.; Dunn, P.A.; Tablante, N. Review of nonfoodborne zoonotic and potentially zoonotic poultry diseases. Avian. Dis. 2016, 60, 553–575. [Google Scholar] [CrossRef]
- Young, N.; O’sullivan, G. The influence of ingredients on product stability and shelf life. In Food and Beverage Stability and Shelf Life; Elsevier: Amsterdam, The Netherlands, 2011; pp. 132–183. [Google Scholar]
- Braiek, O.B.; Smaoui, S. Enterococci: Between emerging pathogens and potential probiotics. Biomed. Res. Int. 2019, 2019, 5938210. [Google Scholar]
- Franz, C.M.; Van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef]
- Silva, C.C.; Silva, S.P.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Hammami, R.; Fliss, I.; Corsetti, A. Application of protective cultures and bacteriocins for food biopreservation. Front. Microbiol. 2019, 10, 1561. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, M.S.; Clewell, D.B.; Ike, Y.; Shankar, N. Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Multiple bacteriocin production in lactic acid bacteria. J. Biosci. Bioeng. 2022, 134, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Cintas, L.M.; Casaus, P.; Håvarstein, L.S.; Hernandez, P.E.; Nes, I.F. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl. Environ. Microbiol. 1997, 63, 4321–4330. [Google Scholar] [CrossRef] [Green Version]
- Cintas, L.M.; Casaus, P.; Herranz, C.; Håvarstein, L.S.; Holo, H.; Hernández, P.E.; Nes, I.F. Biochemical and genetic evidence that Enterococcus faecium L50 produces enterocins L50A and L50B, the sec-dependent enterocin P, and a novel bacteriocin secreted without an N-terminal extension termed enterocin Q. J. Bacteriol. 2000, 182, 6806–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criado, R.; Gutiérrez, J.; Budin-Verneuil, A.; Hernandez, P.; Hartke, A.; Cintas, L.; Auffray, Y.; Benachour, A. Molecular analysis of the replication region of the pCIZ2 plasmid from the multiple bacteriocin producer strain Enterococcus faecium L50. Plasmid 2008, 60, 181–189. [Google Scholar] [CrossRef]
- Abriouel, H.; Ben Omar, N.; Lucas, R.; Martínez-Cañamero, M.; Galvez, A. Bacteriocin production, plasmid content and plasmid location of enterocin P structural gene in enterococci isolated from food sources. Lett. Appl. Microbiol. 2006, 42, 331–337. [Google Scholar] [CrossRef]
- Ness, I.F.; Diep, D.B.; Ike, Y. Enterococcal Bacteriocins and Antimicrobial Proteins that Contribute to Niche Control; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Cintas, L.M.; Casaus, P.; Holo, H.; Hernandez, P.E.; Nes, I.F.; Håvarstein, L.S. Enterocins L50A and L50B, two novel bacteriocins from Enterococcus faecium L50, are related to staphylococcal hemolysins. J. Bacteriol. 1998, 180, 1988–1994. [Google Scholar] [CrossRef] [Green Version]
- Criado, R.; Gutiérrez, J.; Martín, M.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Immunochemical characterization of temperature-regulated production of enterocin L50 (EntL50A and EntL50B), enterocin P, and enterocin Q by Enterococcus faecium L50. Appl. Environ. Microbiol. 2006, 72, 7634–7643. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Tomita, H.; Ike, Y. Bac 32, a novel bacteriocin widely disseminated among clinical isolates of Enterococcus faecium. Antimicrob. Agents Chemoter. 2006, 50, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, E.; Cai, Y.; Marchioni, E.; Ennahar, S. Genetic identification of the bacteriocins produced by Enterococcus faecium IT62 and evidence that bacteriocin 32 is identical to enterocin IT. Antimicrob. Agents Chemoter. 2009, 53, 1907–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanchi, H.; Hammami, R.; Fernandez, B.; Kourda, R.; Ben Hamida, J.; Fliss, I. Simultaneous production of formylated and nonformylated enterocins L50A and L50B as well as 61A, a new glycosylated durancin, by Enterococcus durans 61A, a strain isolated from artisanal fermented milk in Tunisia. J. Agric. Food Chem. 2016, 64, 3584–3590. [Google Scholar] [CrossRef]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Future Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Aymerich, T.; Holo, H.; Håvarstein, L.S.; Hugas, M.; Garriga, M.; Nes, I.F. Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a new antilisterial bacteriocin in the pediocin family of bacteriocins. Appl. Environ. Microbiol. 1996, 62, 1676–1682. [Google Scholar] [CrossRef] [Green Version]
- Casaus, P.; Nilsen, T.; Cintas, L.M.; Nes, I.F.; Hernández, P.E.; Holo, H. Enterocin B, a new bacteriocin from Enterococcus faecium T136 which can act synergistically with enterocin A. Microbiology 1997, 143, 2287–2294. [Google Scholar] [CrossRef] [Green Version]
- Ankaiah, D.; Palanichamy, E.; Antonyraj, C.B.; Ayyanna, R.; Perumal, V.; Ahamed, S.I.B.; Arul, V. Cloning, overexpression, purification of bacteriocin enterocin-B and structural analysis, interaction determination of enterocin-A, B against pathogenic bacteria and human cancer cells. Int. J. Biol. Macromol. 2018, 116, 502–512. [Google Scholar] [CrossRef]
- Ishibashi, N.; Himeno, K.; Fujita, K.; Masuda, Y.; Perez, R.H.; Zendo, T.; Wilaipun, P.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Purification and characterization of multiple bacteriocins and an inducing peptide produced by Enterococcus faecium NKR-5-3 from Thai fermented fish. Biosci. Biotechnol. Biochem. 2012, 76, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Himeno, K.; Rosengren, K.J.; Inoue, T.; Perez, R.H.; Colgrave, M.L.; Lee, H.S.; Chan, L.Y.; Henriques, S.n.T.; Fujita, K.; Ishibashi, N. Identification, characterization, and three-dimensional structure of the novel circular bacteriocin, enterocin NKR-5-3B, from Enterococcus faecium. Biochemistry 2015, 54, 4863–4876. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, N.; Matsumoto, N.; Perez, R.H.; Iwatani, S.; Sugino, H.; Zendo, T.; Wilaipun, P.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Molecular characterization of the possible regulation of multiple bacteriocin production through a three-component regulatory system in Enterococcus faecium NKR-5-3. J. Biosci. Bioeng. 2021, 131, 131–138. [Google Scholar] [CrossRef]
- Ishibashi, N.; Himeno, K.; Masuda, Y.; Perez, R.H.; Iwatani, S.; Zendo, T.; Wilaipun, P.; Leelawatcharamas, V.; Nakayama, J.; Sonomoto, K. Gene cluster responsible for secretion of and immunity to multiple bacteriocins, the NKR-5-3 enterocins. Appl. Environ. Microbiol. 2014, 80, 6647–6655. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Kaminaka, K.; Shima, J.; KAwAMoTo, S.; MoRI, K.; Choi, S.-H.; Doi, K.; OHMoMo, S.; Ogata, S. Isolation and characterization of enterocin SE-K4 produced by thermophilic enterococci, Enterococcus faecalis K-4. Biosci. Biotechnol. Biochem. 2001, 65, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Navaratna, M.A.; Sahl, H.-G.; Tagg, J.R. Two-component anti-Staphylococcus aureus lantibiotic activity produced by Staphylococcus aureus C55. Appl. Environ. Microbiol. 1998, 64, 4803–4808. [Google Scholar] [CrossRef] [Green Version]
- Kawada-Matsuo, M.; Shammi, F.; Oogai, Y.; Nakamura, N.; Sugai, M.; Komatsuzawa, H. C55 bacteriocin produced by ETB—plasmid positive Staphylococcus aureus strains is a key factor for competition with S. aureus strains. Microbiol. Immunol. 2016, 60, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Portillo, A.; Ruiz-Larrea, F.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide resistance genes in Enterococcus spp. Antimicrob. Agents Chemoter. 2000, 44, 967–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, R.; Galán, J.C.; Tenorio, C.; Ruiz-Garbajosa, P.; Zarazaga, M.; Torres, C.; Baquero, F. New aac(6′)-I genes in Enterococcus hirae and Enterococcus durans: Effect on β-lactam/aminoglycoside synergy. J. Antimicrob. Chemother. 2005, 55, 1053–1055. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert. Rev. Anti Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Fontana, R.; Ligozzi, M.; Pittaluga, F.; Satta, G. Intrinsic penicillin resistance in enterococci. Microb. Drug Resist. 1996, 2, 209–213. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in Enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [Green Version]
- Poeta, P.; Costa, D.; Igrejas, G.; Saenz, Y.; Zarazaga, M.; Rodrigues, J.; Torres, C. Polymorphisms of the pbp5 gene and correlation with ampicillin resistance in Enterococcus faecium isolates of animal origin. J. Med. Microbiol. 2007, 56, 236–240. [Google Scholar] [CrossRef]
- López, M.; Tenorio, C.; Del Campo, R.; Zarazaga, M.; Torres, C. Characterization of the mechanisms of fluoroquinolone resistance in vancomycin-resistant enterococci of different origins. J. Chemother. 2011, 23, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Cercenado, E. Enterococcus: Resistencias fenotípicas y genotípicas y epidemiología en España. Enferm. Infecc. Microbiol. Clin. 2011, 29, 59–65. [Google Scholar] [CrossRef]
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmore, M.S.; Clewell, D.B.; Courvalin, P.; Dunny, G.M.; Murray, B.E.; Rice, L.B. The enterococci: Pathogenesis, Molecular Biology, and Antibiotic Resistance; ASM Press: Washington, DC, USA, 2002; Volume 10. [Google Scholar]
- Nakayama, J.; Kariyama, R.; Kumon, H. Description of a 23.9-kilobase chromosomal deletion containing a region encoding fsr genes which mainly determines the gelatinase-negative phenotype of clinical isolates of Enterococcus faecalis in urine. Appl. Environ. Microbiol. 2002, 68, 3152–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Nallapareddy, S.R.; Singh, K.V.; Okhuysen, P.C.; Murray, B.E. A functional collagen adhesin gene, acm, in clinical isolates of Enterococcus faecium correlates with the recent success of this emerging nosocomial pathogen. Infect. Immun. 2008, 76, 4110–4119. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, B.; Wityk, P.; Gałęcka, M.; Michalik, M. The many faces of Enterococcus spp.—Commensal, probiotic and opportunistic pathogen. Microorganisms 2021, 9, 1900. [Google Scholar] [CrossRef]
- Poeta, P.; Costa, D.; Rojo-Bezares, B.; Zarazaga, M.; Klibi, N.; Rodrigues, J.; Torres, C. Detection of antimicrobial activities and bacteriocin structural genes in faecal enterococci of wild animals. Microbiol. Res. 2007, 162, 257–263. [Google Scholar] [CrossRef]
- Bennett, S.; Ben Said, L.; Lacasse, P.; Malouin, F.; Fliss, I. Susceptibility to nisin, bactofencin, pediocin and reuterin of multidrug resistant Staphylococcus aureus, Streptococcus dysgalactiae and Streptococcus uberis causing bovine mastitis. Antibiotics 2021, 10, 1418. [Google Scholar] [CrossRef]
- Lo Verso, L.; Lessard, M.; Talbot, G.; Fernandez, B.; Fliss, I. Isolation and selection of potential probiotic bacteria from the pig gastrointestinal tract. Probiotics Antimicrob. Proteins 2018, 10, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Biron, E.; Ben Said, L.; Subirade, M.; Fliss, I. Bacteriocin-Based Synergetic Consortia: A Promising Strategy to Enhance Antimicrobial Activity and Broaden the Spectrum of Inhibition. Microbiol. Spectr. 2022, 10, e00406–e00421. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of changes to the clinical and laboratory standards institute performance standards for antimicrobial susceptibility testing, M100. Clin. Microbiol. Newsl. 2021, 59, e00213–e00221. [Google Scholar] [CrossRef]
- Poeta, P.; Costa, D.; Klibi, N.; Rodrigues, J.; Torres, C. Phenotypic and Genotypic Study of Gelatinase and β--Haemolysis Activities in Faecal Enterococci of Poultry in Portugal. J. Vet. Med. 2006, 53, 203–208. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. Spec. Publ. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- van Heel, A.J.; de Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. Spec. Publ. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. Clin. Microbiol. Newsl. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malberg Tetzschner, A.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In silico genotyping of Escherichia coli isolates for extraintestinal virulence genes by use of whole-genome sequencing data. Clin. Microbiol. Newsl. 2020, 58, e01220–e01269. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemoter. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M. Multilocus sequence typing of total-genome-sequenced bacteria. Clin. Microbiol. Newsl. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.D.n.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. Clin. Microbiol. Newsl. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.W.; Fung, R.; Golubchik, T.; Harding, R.M.; Jeffery, K.J.; Jolley, K.A. Multilocus sequence typing of Clostridium difficile. Clin. Microbiol. Newsl. 2010, 48, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Lemee, L.; Dhalluin, A.; Pestel-Caron, M.; Lemeland, J.-F.; Pons, J.-L. Multilocus sequence typing analysis of human and animal Clostridium difficile isolates of various toxigenic types. Clin. Microbiol. Newsl. 2004, 42, 2609–2617. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed]
- Jaureguy, F.; Landraud, L.; Passet, V.; Diancourt, L.; Frapy, E.; Guigon, G.; Carbonnelle, E.; Lortholary, O.; Clermont, O.; Denamur, E. Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genom. 2008, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Number of BP+ Strains with Activity against the Indicator Strain | |||||||
|---|---|---|---|---|---|---|---|
| E. faecium (n = 11) | E. gallinarum (n = 9) | E. faecalis (n = 8) | E. durans (n = 3) | E. casseliflavus (n = 1) | Total | ||
| Indicator strains a | C. perfringens (X2967) | 11 | 9 | 8 | 3 | 1 | 32 |
| E. hirae (C1436) | 11 | 1 | 7 | 3 | 1 | 23 | |
| E. durans (C1433) | 11 | 5 | 2 | 3 | 1 | 22 | |
| E. casseliflavus (C1232) | 8 | 3 | 6 | 3 | - | 20 | |
| E. faecium (C2321) | 10 | 3 | 3 | 3 | - | 19 | |
| E. faecalis (C410) | 11 | 1 | 3 | 3 | - | 18 | |
| E. gallinarum (C2310) | 9 | 3 | 3 | 3 | - | 18 | |
| L. monocytogenes (C137) | 8 | 4 | 4 | 3 | - | 19 | |
| M. luteus (C157) | 1 | 1 | 7 | 3 | 1 | 13 | |
| S. suis (C2058) | 2 | - | 6 | 3 | 1 | 7 | |
| MRSA b (C411) | - | 1 | - | - | - | 1 | |
| MSSA b (C1570) | - | 1 | - | - | - | 1 | |
| BP+ Strain | Origin | Species | Number of C. perfringens (of 20 Tested) Inhibited by the Antimicrobial Activity of the Extracts of BP+ Strains | ||
|---|---|---|---|---|---|
| Non-Heated Supernatant | Heated Supernatant | Concentrated Supernatant | |||
| C1446 | Poultry | E. gallinarum | -a | - | 11 |
| X2829 | Poultry | E. faecium | 2 | 2 | 12 |
| X3036 | Poultry | E. gallinarum | - | - | 2 |
| X3179 | Poultry | E. faecium | 4 | 4 | 18 |
| X2903 | Poultry | E. faecium | - | - | 8 |
| X2947 | Poultry | E. faecium | - | - | 4 |
| X2956 | Poultry | E. faecium | - | - | 3 |
| X2960 | Poultry | E. faecium | - | - | 4 |
| X3187 | Poultry | E. faecalis | - | - | 1 |
| X3220 | Poultry | E. faecium | - | - | 1 |
| X3198 | Poultry | E. faecalis | - | - | 1 |
| X3204 | Poultry | E. faecium | 2 | 2 | 16 |
| X2906 | Poultry | E. faecium | 2 | 2 | 8 |
| 61A | Cow milk | E. durans | 4 | 3 | 20 |
| 42G | Cow milk | E. durans | 8 | 5 | 18 |
| LCW03 | Camel milk | E. durans | - | - | 18 |
| LCW44 | Camel milk | E. durans | - | - | 16 |
| LCW06 | Camel milk | E. durans | - | - | 16 |
| Strain | Species | Putative Enterocins |
|---|---|---|
| 42G | E. durans | Enterocin P, Enterocin L50 A/B |
| 61A | E. durans | Enterocin P, Enterocin L50 A/B |
| LCW03 | E. durans | Enterocin P, Enterocin L50 A/B, Bacteriocin 32 |
| LCW06 | E. durans | Enterocin P, Enterocin L50 A/B, Bacteriocin 32 |
| LCW44 | E. durans | Enterocin P, Enterocin L50 A/B, Bacteriocin 32 |
| X2893 | E. faecium | Enterocin A, Enterocin B |
| X2903 | E. faecium | Enterocins NKR-5-3A; Enterocin NKR-5-3D, Enterocin NKR-3-5-3-Z |
| X2906 | E. faecium | Enterocin A, Enterocin B |
| X3179 | E. faecium | Enterocin A, Enterocin B, Enterocin NKR-5-3A, Enterocin NKR-5-3D, Enterocin NKR-5-3-Z |
| X3204 | E. faecium | Enterocin A, Enterocin B |
| X3198 | E. faecalis | Ent SE-K4, Staphylococcin C55a/b |
| X3187 | E. faecalis | Ent SE-K4, Staphylococcin C55a/b |
| Strain | Species | Origin | Antibiotic Resistance Phenotype | Antibiotic Resistance Genotype | |
|---|---|---|---|---|---|
| Intrinsic Mechanisms | Acquired Mechanisms | ||||
| X2893 | E. faecium | Poultry | CIP | msr(C), aac(6′)-Ii | - |
| X3179 | E. faecium | CIP, E | msr(C), aac(6′)-Ii | erm(B) | |
| X2903 | E. faecium | CIP, E, P, C | msr(C), aac(6′)-Ii | erm(B), fexB, poxtA | |
| X3187 | E. faecalis | CIP, E, P, TE, CN, S | Isa(A) | erm(B), aac(6′)-aph(2″), tet(M) | |
| X3198 | E. faecalis | CIP, TE | Isa(A) | erm(B), aac(6′)-aph(2″), tet(M) | |
| X3204 | E. faecium | CIP, TE | msr(C), aac(6′)-Ii | str, tet(M), tet(L), cat | |
| X2906 | E. faecium | CIP | msr(C), aac(6′)-Ii | - | |
| 61A | E. durans | Other | Susceptible | aac(6′)-Iih | - |
| 42G | E. durans | Susceptible | aac(6′)-Iih | - | |
| LCW03 | E. durans | Susceptible | aac(6′)-Iih | - | |
| LCW44 | E. durans | Susceptible | aac(6′)-Iih | - | |
| LCW06 | E. durans | Susceptible | aac(6′)-Iih | - | |
| Strain | Species | Virulence Genes | Sequence Type |
|---|---|---|---|
| X3187 | E. faecalis | ElrA, SrtA, ace, cCF10, cOB1, cad, camE, ebpC, efaAfs, fsrB, gelE, hylA, hylB, and tpx | ST397 |
| X3198 | E. faecalis | ElrA, SrtA, ace, cCF10, cOB1, cad, camE, ebpC, efaAfs, fsrB, gelE, hylA, hylB, and tpx | ST397 |
| X2893 | E. faecium | acm | ST722 |
| X2903 | E. faecium | acm | New allelic combination: adk-1, atpA-2, ddl-7, gdh-57, gyd-1, pstS-80, purK-6 |
| X2906 | E. faecium | acm | ST722 |
| X3179 | E. faecium | acm | New allelic combination: adk-1, atpA-2, ddl-7, gdh-76, gdh-2, gyd-1, pstS-1, purK-3 |
| X3204 | E. faecium | acm | ST784 |
| Strain | Species | Type | Replicon Plasmid |
|---|---|---|---|
| X2893 | E. faecium | - | - |
| X3179 | E. faecium | Rep3 | rep29 |
| Inc18 | rep1, rep2 | ||
| RepA_N | repUS15 | ||
| X2903 | E. faecium | Rep3 | rep29 |
| Rep1 | repUS58 | ||
| Inc18 | rep1 | ||
| Rep_trans | rep14a | ||
| RepA_N | repUS15 | ||
| X3187 | E. faecalis | Rep_trans | repUS43 |
| X3198 | E. faecalis | Rep_trans | repUS43 |
| X3204 | E. faecium | RepA_N | repUS15 |
| Inc18 | rep1 | ||
| Rep1 | rep22 | ||
| X2906 | E. faecium | - | - |
| 61A | E. durans | RepA_N | repUS15 |
| Rep1 | repUS64 | ||
| Inc18 | rep1, rep2 | ||
| 42G | E. durans | RepA_N | repUS15 |
| Rep1 | repUS64 | ||
| Inc18 | rep1, rep2 | ||
| LCW03 | E. durans | Rep3 | rep18a |
| Inc18 | rep1, rep2 | ||
| RepA_N | repUS15 | ||
| LCW44 | E. durans | Inc18 | rep1, rep2 |
| Rep3 | rep18a | ||
| RepA_N | repUS15 | ||
| LCW06 | E. durans | Inc18 | rep1, rep2 |
| Rep3 | rep18a | ||
| RepA_N | repUS15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Vela, S.; Ben Said, L.; Soltani, S.; Guerbaa, R.; Fernández-Fernández, R.; Ben Yahia, H.; Ben Slama, K.; Torres, C.; Fliss, I. Targeting Enterococci with Antimicrobial Activity against Clostridium perfringens from Poultry. Antibiotics 2023, 12, 231. https://doi.org/10.3390/antibiotics12020231
García-Vela S, Ben Said L, Soltani S, Guerbaa R, Fernández-Fernández R, Ben Yahia H, Ben Slama K, Torres C, Fliss I. Targeting Enterococci with Antimicrobial Activity against Clostridium perfringens from Poultry. Antibiotics. 2023; 12(2):231. https://doi.org/10.3390/antibiotics12020231
Chicago/Turabian StyleGarcía-Vela, Sara, Laila Ben Said, Samira Soltani, Ramzi Guerbaa, Rosa Fernández-Fernández, Houssem Ben Yahia, Karim Ben Slama, Carmen Torres, and Ismail Fliss. 2023. "Targeting Enterococci with Antimicrobial Activity against Clostridium perfringens from Poultry" Antibiotics 12, no. 2: 231. https://doi.org/10.3390/antibiotics12020231
APA StyleGarcía-Vela, S., Ben Said, L., Soltani, S., Guerbaa, R., Fernández-Fernández, R., Ben Yahia, H., Ben Slama, K., Torres, C., & Fliss, I. (2023). Targeting Enterococci with Antimicrobial Activity against Clostridium perfringens from Poultry. Antibiotics, 12(2), 231. https://doi.org/10.3390/antibiotics12020231