
Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates
 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Antibiotic Susceptibility Testing
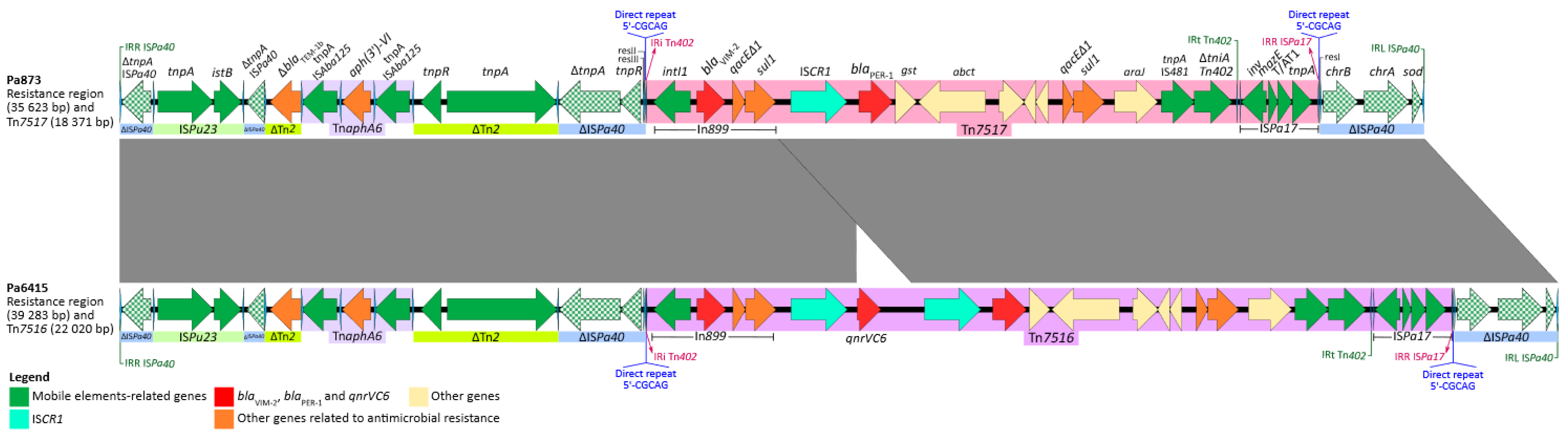
2.2. Genetic Features of Pa6415
2.3. Genetic features of Pa873
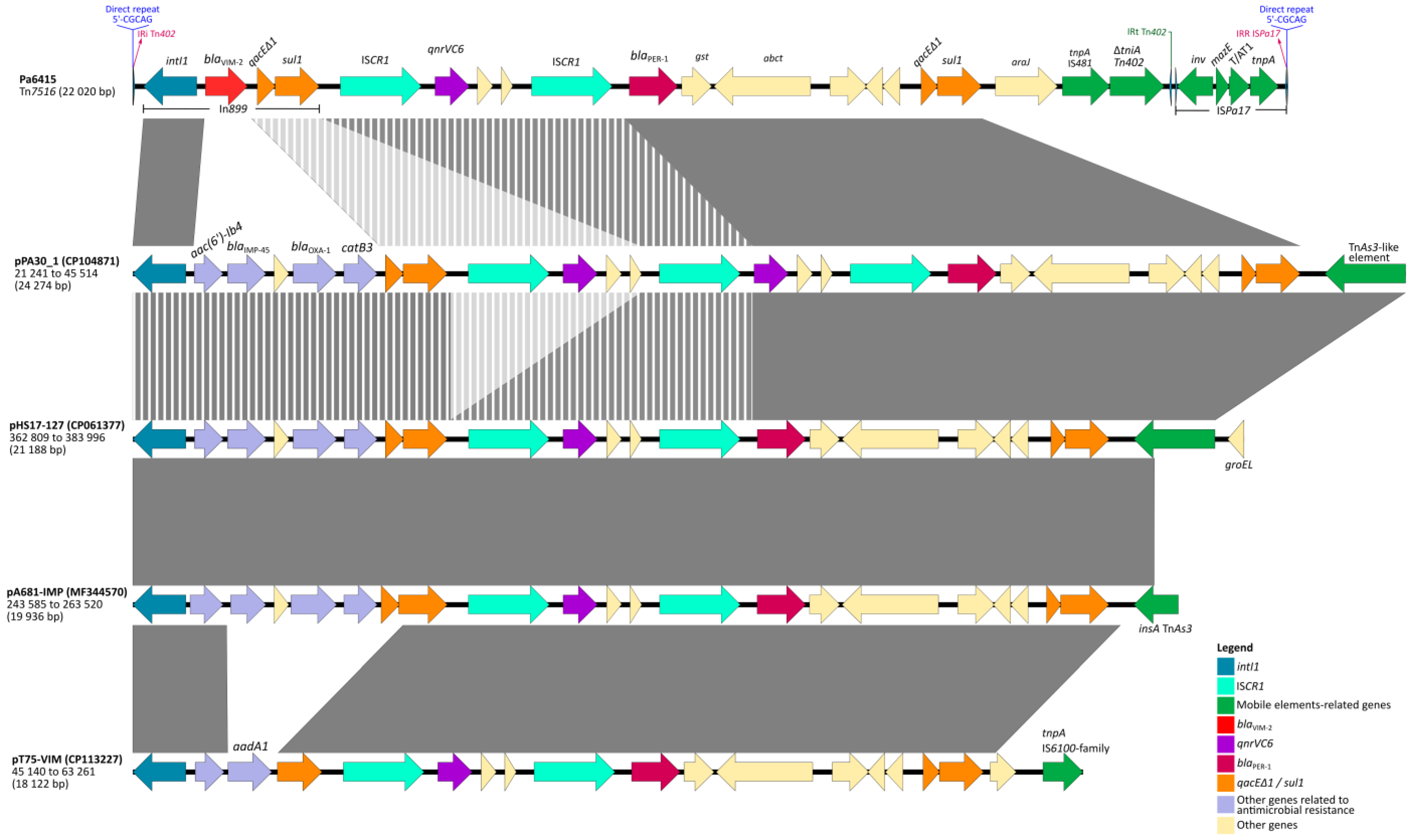
2.4. Resistance Regions and New Transposition Units (Tn7516 and Tn7517)
2.5. BLAST Analysis
3. Discussion
4. Materials and Methods
4.1. Strains, Identification, and Antibiotic Susceptibility Testing
4.2. Short- and Long-Read Genome Sequencing
4.3. Sequence Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. WHO Pathogens priority list working group. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Jian, J.; Yu, P.; Zheng-Li, C.; Hao, L.; Ze-Jing, W.; Shao-Shuo, Y.; Yu, S.; Guang-Yi, W.; Shi-Hui, Z.; Bing, M.; et al. Epidemiological retrospective analysis in major burn patients: Single centre medical records from 2009 to 2019. Updates Surg. 2022, 74, 1453–1459. [Google Scholar] [CrossRef]
- del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents. 2020, 56, 106196. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; Van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2022 guidance on the treatment of extended-spectrum β-lactamase producing Enterobacterales (ESBL-E), carbapenem-resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with difficult-to-treat resistance (DTR- P. aeruginosa). Clin. Infect. Dis. 2022, 75, 187–212. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Ortiz de la Rosa, J.M.; Sadek, M.; Nordmann, P. Impact of acquired broad-spectrum β-lactamases on susceptibility to cefiderocol and newly developed β-lactam/β-lactamase inhibitor combinations in Escherichia coli and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2022, 66, e0003922. [Google Scholar] [CrossRef]
- Yao, J.; Wang, J.; Chen, M.; Cai, Y. Cefiderocol: An overview of its in-vitro and in-vivo activity and underlying resistant mechanisms. Front. Med. 2021, 8, 741940. [Google Scholar] [CrossRef] [PubMed]
- Brovedan, M.A.; Marchiaro, P.M.; Díaz, M.S.; Faccone, D.; Corso, A.; Pasteran, F.; Viale, A.M.; Limansky, A.S. Psedomonas putida group species as reservoirs of mobilizable Tn402-like class 1 integrons carrying blaVIM-2 metallo-β-lactamase genes. Infect. Genet. Evol. 2021, 96, 105131. [Google Scholar] [CrossRef]
- Papa-Ezdra, R.; Bado, I.; Cordeiro, N.; Gutierrez, C.; Hitateguy, P.; Seija, V.; Vignoli, R. VIM-2-Producing Pseudomonas spp. in Uruguay: Sequence types, pulsotypes, and class 1 integrons including new variable regions featuring blaVIM-2 and blaGES-7. Antimicrob. Agents Chemother. 2016, 60, 5620–5622. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Acquired metallo-β-lactamases and their genetic association with class 1 integrons and ISCR elements in Gram-negative bacteria. Future Microbiol. 2015, 10, 873–887. [Google Scholar] [CrossRef]
- Zhan, Z.; Hu, L.; Jiang, X.; Zeng, L.; Feng, J.; Wu, W.; Chen, W.; Yang, H.; Yang, W.; Gao, B.; et al. Plasmid and chromosomal integration of four novel blaIMP-carrying transposons from Pseudomonas aeruginosa, Klebsiella pneumoniae and an Enterobacter sp. J. Antimicrob. Chemother. 2018, 73, 3005–3015. [Google Scholar] [CrossRef]
- Toleman, M.A.; Walsh, T.R. 2011. Combinatorial events of insertion sequences and ICE in Gram-negative bacteria. FEMS Microbiol. Rev. 2011, 35, 912–935. [Google Scholar] [CrossRef]
- Wu, J.; Xie, L.; Zhang, F.; Ni, Y.; Sun, J. Molecular characterization of ISCR1-mediated blaPER-1 in a non-O1, non-O139 Vibrio cholerae strain from China. Antimicrob. Agents Chemother. 2015, 59, 4293–4295. [Google Scholar] [CrossRef]
- Bush, K. Carbapenemases: Partners in crime. J. Glob. Antimicrob. Resist. 2013, 1, 7–16. [Google Scholar] [CrossRef]
- Ortiz de la Rosa, J.M.; Nordmann, P.; Poirel, L. ESBLs and resistance to ceftazidime/avibactam and ceftolozane/tazobactam combinations in Escherichia coli and Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2019, 74, 1934–1939. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Bado, I.; Caiata, L.; Vignoli, R.; Seija, V. First report of Pseudomonas aeruginosa co-harbouring blaVIM-2 and blaPER-1 in Latin America. J. Glob. Antimicrob. Resist. 2018, 15, 121–122. [Google Scholar] [CrossRef]
- Thacharodi, A.; Lamont, I.L. Aminoglycoside-modifying enzymes are sufficient to make Pseudomonas aeruginosa clinically resistant to key antibiotics. Antibiotics 2022, 11, 884. [Google Scholar] [CrossRef] [PubMed]
- Costello, S.E.; Deshpande, L.M.; Davis, A.P.; Mendes, R.E.; Castanheira, M. Aminoglycoside-modifying enzyme and 16S ribosomal RNA methyltransferase genes among a global collection of Gram-negative isolates. J. Glob. Antimicrob. Resist. 2019, 16, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Nigro, S.J.; Post, V.; Hall, R.M. Aminoglycoside resistance in multiply antibiotic-resistant Acinetobacter baumannii belonging to global clone 2 from Australian hospitals. J. Antimicrob. Chemother. 2011, 66, 1504–1509. [Google Scholar] [CrossRef]
- Tansirichaiya, S.; Rahman, M.D.A.; Roberts, A.P. The Transposon Registry. Mob. DNA 2019, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655, Erratum in Lancet 2022, 400, 1102. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, C.; Wang, Q.; Zeng, Y.; Sun, Q.; Shu, L.; Lu, J.; Cai, J.; Wang, S.; Zhang, R.; et al. Emergence and expansion of a carbapenem-resistant Pseudomonas aeruginosa clone are associated with plasmid-borne blaKPC-2 and virulence-related genes. mSystems 2021, 6, e00154-21. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Peng, W.; Wu, Y.; Li, H.; Wang, Q.; Yi, H.; Zhang, R.; Shao, B.; Zhu, K. A potential high-risk clone of Pseudomonas aeruginosa ST463. Front. Microbiol. 2021, 12, 670202. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, Y.; Zhang, P.; Wang, J.; Yuan, Q.; Shi, W.; Zhang, S.; Feng, H.; Chen, Y.; Yu, M.; et al. Bloodstream infections caused by Klebsiella pneumoniae carbapenemase-producing P. aeruginosa sequence type 463, associated with high mortality rates in China: A retrospective cohort study. Front. Cell. Infect. Microbiol. 2021, 11, 756782. [Google Scholar] [CrossRef]
- Bado, I.; Papa, R.; Delgado, J.F.; Gaudio, M.; Gutiérrez, C.; Cordeiro, N.F.; García-Fulgueiras, V.; Araújo, L.; Seija, V.; Medina, J.C.; et al. Molecular characterization of carbapenem-resistant Acinetobacter baumannii in the ICU of Uruguay’s University Hospital identifies the first rmtC gene in the species. Microb. Drug Resist. 2018, 24, 1012–1019. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, L.; Li, D.; Li, P.; Yuan, L.; Yang, F.; Guo, Q.; Wang, M. An IncP-2 plasmid sublineage associated with dissemination of blaIMP-45 among carbapenem-resistant Pseudomonas aeruginosa. Emerg. Microbes Infect. 2021, 10, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Cabanne, L.; Vahaboglu, H.; Nordmann, P. Genetic environment and expression of the extended-spectrum β-lactamase blaPER-1 gene in gram-negative bacteria. Antimicrob. Agents Chemother. 2005, 49, 1708–1713. [Google Scholar] [CrossRef]
- Torrens, G.; van der Schalk, T.E.; Cortes-Lara, S.; Timbermont, L.; Del Barrio-Tofiño, E.; Xavier, B.B.; Zamorano, L.; Lammens, C.; Ali, O.; Ruzin, A.; et al. Susceptibility profiles and resistance genomics of Pseudomonas aeruginosa isolates from European ICUs participating in the ASPIRE-ICU trial. J. Antimicrob. Chemother. 2022, 77, 1862–1872. [Google Scholar] [CrossRef]
- Pavez, M.; Troncoso, C.; Osses, I.; Salazar, R.; Illesca, V.; Reydet, P.; Rodríguez, C.; Chahin, C.; Concha, C.; Barrientos, L. High prevalence of CTX-M-1 group in ESBL-producing Enterobacteriaceae infection in intensive care units in southern Chile. Braz. J. Infect. Dis. 2019, 23, 102–110. [Google Scholar] [CrossRef]
- Bastidas-Caldes, C.; Romero-Alvarez, D.; Valdez-Vélez, V.; Morales, R.D.; Montalvo-Hernández, A.; Gomes-Dias, C.; Calvopiña, M. Extended-Spectrum Beta-Lactamases Producing Escherichia coli in South America: A Systematic Review with a One Health Perspective. Infect. Drug Resist. 2022, 15, 5759–5779. [Google Scholar] [CrossRef]
- Polotto, M.; Casella, T.; de Lucca Oliveira, M.G.; Rúbio, F.G.; Nogueira, M.L.; de Almeida, M.T.; Nogueira, M.C. Detection of P. aeruginosa harboring blaCTX-M-2, blaGES-1 and blaGES-5, blaIMP-1 and blaSPM-1 causing infections in Brazilian tertiary-care hospital. BMC Infect. Dis. 2012, 12, 176. [Google Scholar] [CrossRef] [PubMed]
- Pagani, L.; Mantengoli, E.; Migliavacca, R.; Núcleo, E.; Pollini, S.; Spalla, M.; Daturi, R.; Romero, E.; Rossolini, G.M. Multifocal detection of multidrug-resistant Pseudomonas aeruginosa producing the PER-1 extended-spectrum β-lactamase in Northern Italy. J. Clin. Microbiol. 2004, 42, 2523–2529. [Google Scholar] [CrossRef]
- de Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Carver, T.; Berriman, M.; Tivey, A.; Patel, C.; Böhme, U.; Barrell, B.G.; Parkhill, J.; Rajandream, M.A. Artemis and ACT: Viewing, annotating and comparing sequences stored in a relational database. Bioinformatics 2008, 24, 2672–2676. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Pa6415 | Pa873 | |
|---|---|---|
| Date of recovery | November 2016 | October 2021 |
| Sample origin | Cerebrospinal fluid | Tracheal aspirate |
| MLST | ST463 | ST395 |
| Antibiotics 1 | Susceptibility results * (Antimicrobial resistance genes detected) | |
| PTZ | 32 (blaPER-1/blaVIM-2) | 64 (blaPER-1/blaVIM-2) |
| CAZ | ≥256 (blaPER-1/blaVIM-2) | ≥256 (blaPER-1/blaVIM-2) |
| FEP | ≥64 (blaPER-1/blaVIM-2) | ≥64 (blaPER-1/blaVIM-2) |
| IPM | ≥32 (blaVIM-2) | 8 (blaVIM-2) |
| MEM | 1 | 4 (blaVIM-2) |
| GM | 4 | ≤1 |
| AK | ≥64 (aph(3′)-VIa) | ≥64 (aph(3′)-VIa) |
| CIP | 1(qnrVC6) | 0.125 |
| ATM | >256 (blaPER-1) | >256 (blaPER-1) |
| CZA | 4 | 24 (blaVIM-2) |
| CZA + ATM | 1 | 6 |
| FDC (mm) | 26 | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papa-Ezdra, R.; Cordeiro, N.F.; Outeda, M.; Garcia-Fulgueiras, V.; Araújo, L.; Seija, V.; Ayala, J.A.; Bado, I.; Vignoli, R. Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates. Antibiotics 2023, 12, 304. https://doi.org/10.3390/antibiotics12020304
Papa-Ezdra R, Cordeiro NF, Outeda M, Garcia-Fulgueiras V, Araújo L, Seija V, Ayala JA, Bado I, Vignoli R. Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates. Antibiotics. 2023; 12(2):304. https://doi.org/10.3390/antibiotics12020304
Chicago/Turabian StylePapa-Ezdra, Romina, Nicolás F. Cordeiro, Matilde Outeda, Virginia Garcia-Fulgueiras, Lucía Araújo, Verónica Seija, Juan A. Ayala, Inés Bado, and Rafael Vignoli. 2023. "Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates" Antibiotics 12, no. 2: 304. https://doi.org/10.3390/antibiotics12020304
APA StylePapa-Ezdra, R., Cordeiro, N. F., Outeda, M., Garcia-Fulgueiras, V., Araújo, L., Seija, V., Ayala, J. A., Bado, I., & Vignoli, R. (2023). Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates. Antibiotics, 12(2), 304. https://doi.org/10.3390/antibiotics12020304