


Inhibitory Activity of Essential Oils of Mentha spicata and Eucalyptus globulus on Biofilms of Streptococcus mutans in an In Vitro Model

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Chemical Analysis by GC-MS of the Essential Oils of M. spicata and E. globulus
2.2. Antibacterial Activity of the Essential Oil of M. spicata and E. globulus against S. mutans
2.3. Antibiofilm Activity Using an In Vitro Model on Dental Enamel Pieces of Bovine Origin
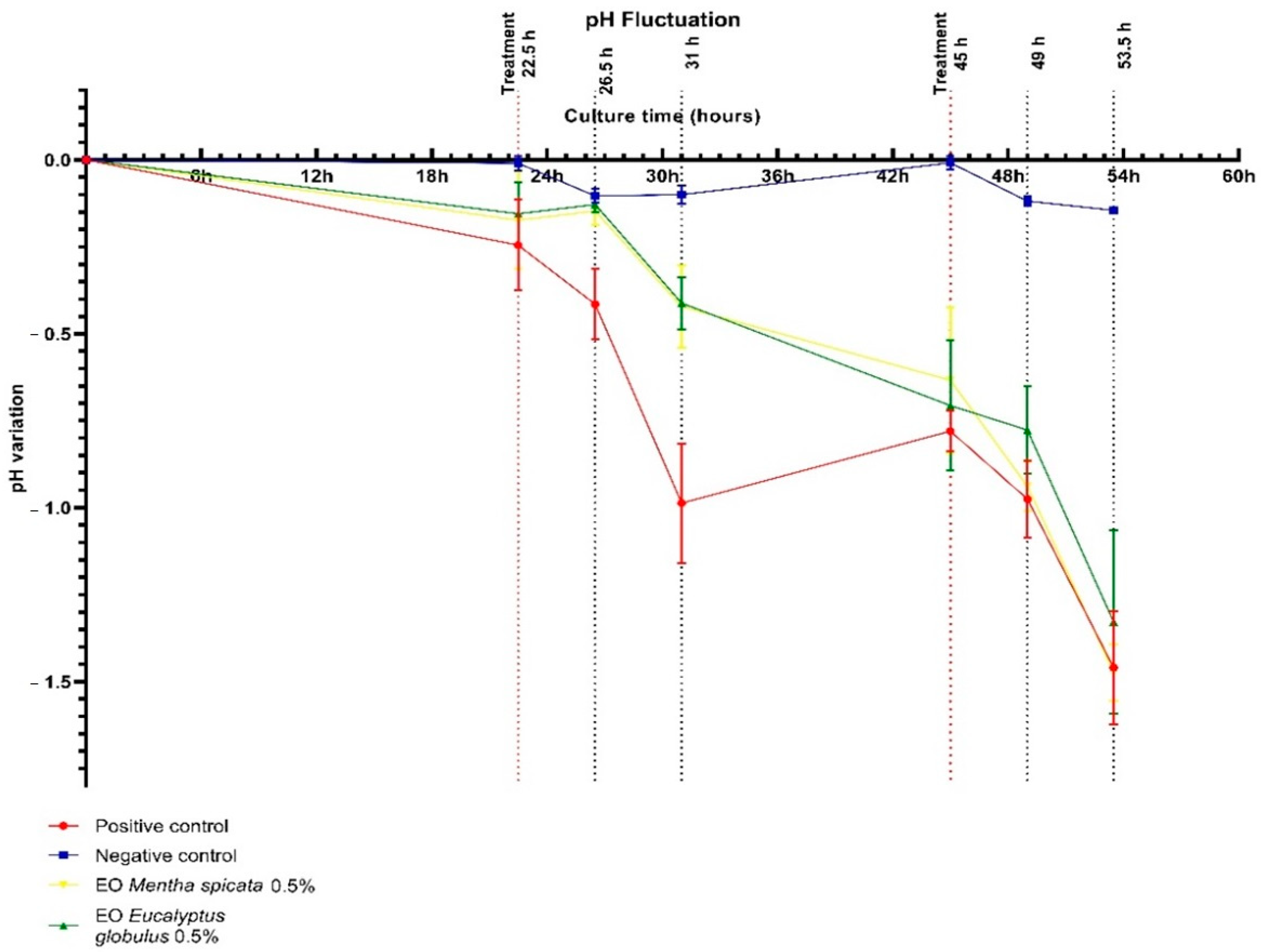
2.3.1. Culture Medium pH Fluctuation
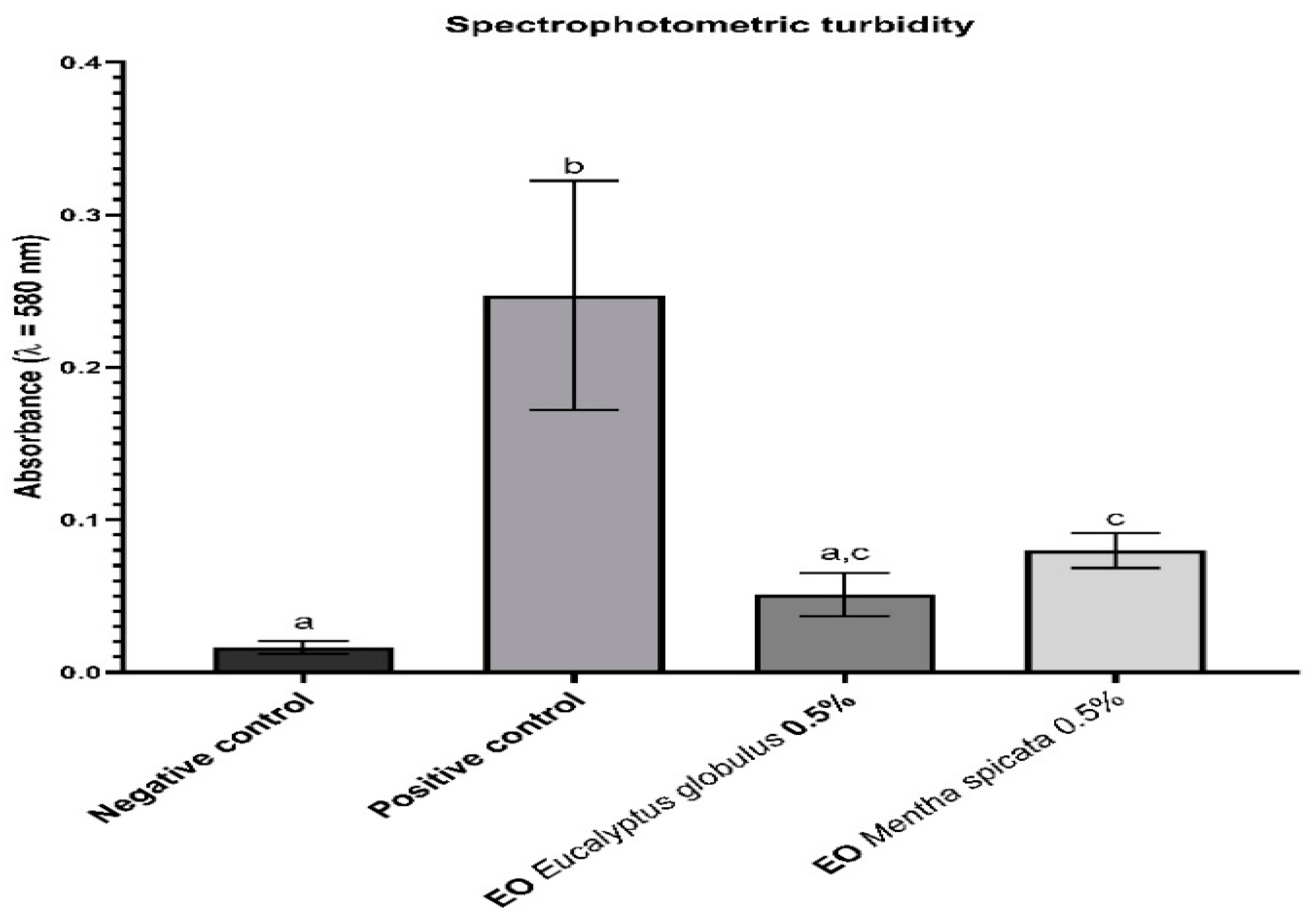
2.3.2. Spectrophotometric Turbidity
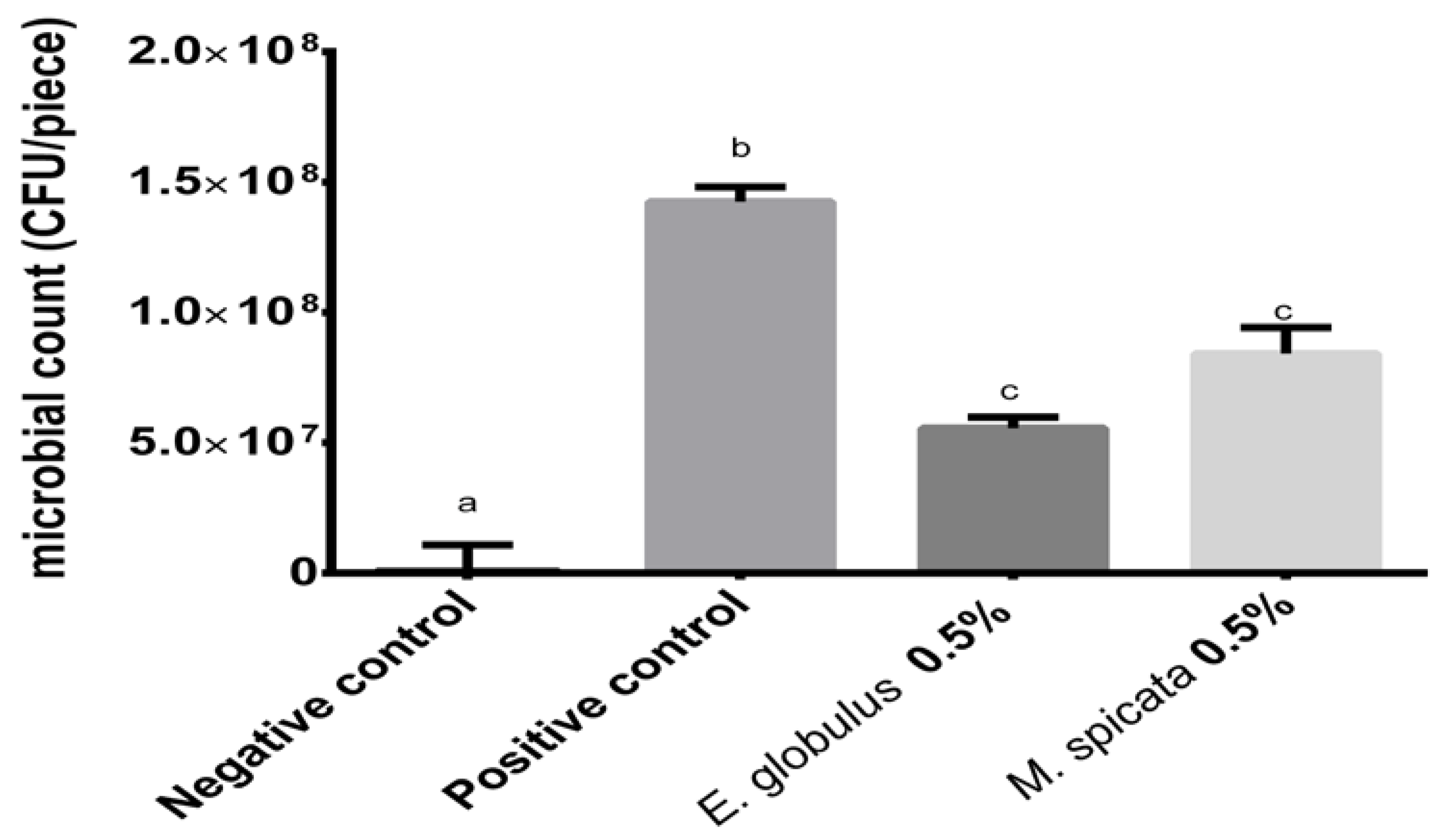
2.3.3. Recoverable Microorganism Count
3. Discussion
4. Materials and Methods
4.1. Obtention of the Essential Oil
4.2. Composition Analysis by Gas Chromatography-Mass Spectrometry (GC/MS) of Essential Oils
4.3. Inhibitory Activity of the Essential Oils M. spicata and E. globulus and Determination of the Minimum Inhibitory Concentration
4.3.1. Evaluation of the Ability to Form Biofilms by the Congo Red Method
4.3.2. Agar-Well Diffusion Method
4.3.3. Determination of the Minimum Inhibitory Concentration (MIC) by the Colorimetric Microdilution Method
4.4. In Vitro Model for the Formation of Biofilms on Dental Enamel Pieces of Bovine Origin
4.4.1. Culture Media
4.4.2. Activation of the Microorganism and Preparation of the Inoculum
4.4.3. Preparation of Enamel Slabs
4.4.4. Biofilm Cultivation and Treatment
4.4.5. Biofilm Collection and Evaluation of the Treatment
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- HOMD: Human Oral Microbiome Database. Available online: https://www.homd.org/ (accessed on 26 April 2022).
- Bertolini, M.; Costa, R.C.; Barão, V.A.R.; Cunha Villar, C.; Retamal-Valdes, B.; Feres, M.; Silva Souza, J.G. Oral Microorganisms and Biofilms: New Insights to Defeat the Main Etiologic Factor of Oral Diseases. Microorganisms 2022, 10, 2413. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.S.; Kranz, H.T.; Schneider, M.; Tietze, J.P.; Piwowarcyk, A.; Kuzius, T.; Arnold, W.; Naumova, E.A. Biofilm Formation on Different Dental Restorative Materials in the Oral Cavity. BMC Oral Health 2020, 20, 162. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Parida, S.; Mangwani, N.; Dash, H.R.; Das, S. Isolation and Characterization of Biofilm-Forming Bacteria and Associated Extracellular Polymeric Substances from Oral Cavity. Ann. Microbiol. 2013, 63, 1553–1562. [Google Scholar] [CrossRef]
- Marsh, P.D. Dental Plaque as a Biofilm and a Microbial Community—Implications for Health and Disease. BMC Oral Health 2006, 6, S14. [Google Scholar] [CrossRef]
- Motallaei, M.N.; Yazdanian, M.; Tebyanian, H.; Tahmasebi, E.; Alam, M.; Abbasi, K.; Seifalian, A.; Ranjbar, R.; Yazdanian, A. The Current Strategies in Controlling Oral Diseases by Herbal and Chemical Materials. Evid.-Based Complement. Altern. Med. 2021, 2021, 3423001. [Google Scholar] [CrossRef]
- Ghazal, T.S.; Levy, S.M.; Childers, N.K.; Carter, K.D.; Caplan, D.J.; Warren, J.J.; Cavanaugh, J.E.; Kolker, J. Mutans Streptococci and Dental Caries: A New Statistical Modeling Approach. Caries Res 2018, 52, 246–252. [Google Scholar] [CrossRef]
- Banas, J.A.; Drake, D.R. Are the Mutans Streptococci Still Considered Relevant to Understanding the Microbial Etiology of Dental Caries? BMC Oral Health 2018, 18, 129. [Google Scholar] [CrossRef]
- de Oliveira Carvalho, I.; Purgato, G.A.; Píccolo, M.S.; Pizziolo, V.R.; Coelho, R.R.; Diaz-Muñoz, G.; Alves Nogueira Diaz, M. In Vitro Anticariogenic and Antibiofilm Activities of Toothpastes Formulated with Essential Oils. Arch. Oral Biol. 2020, 117, 104834. [Google Scholar] [CrossRef]
- Maske, T.T.; van de Sande, F.H.; Arthur, R.A.; Huysmans, M.C.D.N.J.M.; Cenci, M.S. In Vitro Biofilm Models to Study Dental Caries: A Systematic Review. Biofouling 2017, 33, 661–675. [Google Scholar] [CrossRef]
- Matsumoto-Nakano, M. Role of Streptococcus Mutans Surface Proteins for Biofilm Formation. Jpn Dent. Sci. Rev. 2018, 54, 22. [Google Scholar] [CrossRef]
- Bardaweel, S.K.; Bakchiche, B.; ALSalamat, H.A.; Rezzoug, M.; Gherib, A.; Flamini, G. Chemical Composition, Antioxidant, Antimicrobial and Antiproliferative Activities of Essential Oil of Mentha Spicata L. (Lamiaceae) from Algerian Saharan Atlas. BMC Complement. Altern. Med. 2018, 18, 201. [Google Scholar] [CrossRef] [PubMed]
- Fitsiou, E.; Mitropoulou, G.; Spyridopoulou, K.; Tiptiri-Kourpeti, A.; Vamvakias, M.; Bardouki, H.; Panayiotidis, M.Ι.; Galanis, A.; Kourkoutas, Y.; Chlichlia, K.; et al. Phytochemical Profile and Evaluation of the Biological Activities of Essential Oils Derived from the Greek Aromatic Plant Species Ocimum basilicum, Mentha spicata, Pimpinella anisum and Fortunella margarita. Molecules 2016, 21, 1069. [Google Scholar] [CrossRef] [PubMed]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Abu-Zaitoun, S.Y.; Khasati, A.I.; Kalbouneh, S.R. Biological Properties and Bioactive Components of Mentha Spicata L. Essential Oil: Focus on Potential Benefits in the Treatment of Obesity, Alzheimer’s Disease, Dermatophytosis, and Drug-Resistant Infections. Evid.-Based Complement. Altern. Med. 2019, 2019, 3834265. [Google Scholar] [CrossRef]
- Immaroh, N.Z.; Kuliahsari, D.E.; Nugraheni, S.D. Review: Eucalyptus Globulus Essential Oil Extraction Method. IOP Conf. Ser. Earth Environ. Sci 2021, 733, 012103. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.M.; Elnesr, S.S.; Abd El-Hack, E.M. Using Essential Oils to Overcome Bacterial Biofilm Formation and Their Antimicrobial Resistance. Saudi J. Biol. Sci. 2021, 28, 5145. [Google Scholar] [CrossRef]
- Kwiatkowski, P.; Sienkiewicz, M.; Pruss, A.; Łopusiewicz, Ł.; Arszyńska, N.; Wojciechowska-Koszko, I.; Kilanowicz, A.; Kot, B.; Dołęgowska, B. Antibacterial and Anti-Biofilm Activities of Essential Oil Compounds against New Delhi Metallo-β-Lactamase-1-Producing Uropathogenic Klebsiella Pneumoniae Strains. Antibiotics 2022, 11, 147. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; de Feo, V. Mentha Spicata Essential Oil: Chemical Composition, Antioxidant and Antibacterial Activities against Planktonic and Biofilm Cultures of Vibrio Spp. Strains. Molecules 2015, 20, 14402–14424. [Google Scholar] [CrossRef]
- Aggarwal, K.K.; Khanuja, S.P.S.; Ahmad, A.; Gupta, V.K.; Kumar, S. Antimicrobial Activity Profiles of the Two Enantiomers of Limonene and Carvone Isolated from the Oils of Mentha Spicata and Anethum Sowa. Flavour Fragr. J. 2002, 17, 59–63. [Google Scholar] [CrossRef]
- Goldbeck, J.C.; do Nascimento, J.E.; Jacob, R.G.; Fiorentini, Â.M.; da Silva, W.P. Bioactivity of Essential Oils from Eucalyptus Globulus and Eucalyptus Urograndis against Planktonic Cells and Biofilms of Streptococcus Mutans. Ind. Crops. Prod. 2014, 60, 304–309. [Google Scholar] [CrossRef]
- Alghamdi, S. Isolation and Identification of the Oral Bacteria and Their Characterization for Bacteriocin Production in the Oral Cavity. Saudi J. Biol. Sci. 2022, 29, 318. [Google Scholar] [CrossRef] [PubMed]
- Mystkowska, J.; Niemirowicz-Laskowska, K.; Łysik, D.; Tokajuk, G.; Dąbrowski, J.R.; Bucki, R. The Role of Oral Cavity Biofilm on Metallic Biomaterial Surface Destruction–Corrosion and Friction Aspects. Int. J. Mol. Sci. 2018, 19, 743. [Google Scholar] [CrossRef] [PubMed]
- Rasooli, I.; Shayegh, S.; Astaneh, S.D.A. The Effect of Mentha Spicata and Eucalyptus Camaldulensis Essential Oils on Dental Biofilm. Int. J. Dent. Hyg. 2009, 7, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, L.K.D.; Jawale, B.A.; Sharma, S.; Sharma, H.; Kumar, H.S.C.M.; Kulkarni, P.A. Antimicrobial Activity of Commercially Available Essential Oils against Streptococcus Mutans. J. Contemp. Dent. Pract. 2012, 13, 71–74. [Google Scholar] [CrossRef]
- Dhifi, W.; Jelali, N.; Mnif, W.; Litaiem, M.; Hamdi, N. Chemical Composition of the Essential Oil of Mentha Spicata L. From Tunisia and Its Biological Activities. J. Food Biochem. 2013, 37, 362–368. [Google Scholar] [CrossRef]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial Adhesion and Biofilms on Surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Larsen, T.; Fiehn, N.E. Dental Biofilm Infections—An Update. APMIS 2017, 125, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Rosan, B.; Lamont, R.J. Dental Plaque Formation. Microbes Infect. 2000, 2, 1599–1607. [Google Scholar] [CrossRef]
- Leach, S.A. Release and Breakdown of Sialic Acid from Human Salivary Mucin and Its Role in the Formation of Dental Plaque. Nature 1963, 199, 486–487. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Li, M.; Gregory, R.L. Bacterial Interactions in Dental Biofilm. Virulence 2011, 2, 435. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Burne, R.A. Regulation of the GtfBC and Ftf Genes of Streptococcus Mutans in Biofilms in Response to PH and Carbohydrate. Microbiology 2001, 147, 2841–2848. [Google Scholar] [CrossRef]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef]
- van Steijn, G.J.; Nieuw Amerongen, A.V.; Veerman, E.C.I.; Kasanmoentalib, S.; Overdijk, B. Chitinase in Whole and Glandular Human Salivas and in Whole Saliva of Patients with Periodontal Inflammation. Eur. J. Oral Sci. 1999, 107, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Calderon, O.; Chacaltana-Ramos, L.J.; Huayanca-Gutiérrez, I.C.; Algarni, M.A.; Alqarni, M.; Batiha, G.E.S. Chemical Constituents, In Vitro Antioxidant Activity and In Silico Study on NADPH Oxidase of Allium Sativum L. (Garlic) Essential Oil. Antioxidants 2021, 10, 1844. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Armas, J.P.; Arroyo-Acevedo, J.L.; Palomino-Pacheco, M.; Herrera-Calderón, O.; Ortiz-Sánchez, J.M.; Rojas-Armas, A.; Calva, J.; Castro-Luna, A.; Hilario-Vargas, J. The Essential Oil of Cymbopogon Citratus Stapt and Carvacrol: An Approach of the Antitumor Effect on 7,12-Dimethylbenz-[α]-Anthracene (DMBA)-Induced Breast Cancer in Female Rats. Molecules 2020, 25, 3284. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New Method for Detecting Slime Production by Coagulase Negative Staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Rojas, R.; Bustamante, B.; Bauer, J.; Fernández, I.; Albán, J.; Lock, O. Antimicrobial Activity of Selected Peruvian Medicinal Plants. J. Ethnopharmacol. 2003, 88, 199–204. [Google Scholar] [CrossRef] [PubMed]
- CLSI Standard M07; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018.
- Liu, M.; Seidel, V.; Katerere, D.R.; Gray, A.I. Colorimetric Broth Microdilution Method for the Antifungal Screening of Plant Extracts against Yeasts. Methods 2007, 42, 325–329. [Google Scholar] [CrossRef]
- Wong, L.; Sissions, C.H. A Comparison of Human Dental Plaque Microcosm Biofilms Grown in an Undefined Medium and a Chemically Defined Artificial Saliva. Arch. Oral Biol. 2001, 46, 477–486. [Google Scholar] [CrossRef]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the Normal Bacterial Flora of the Oral Cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Compound | Retention Index | % of Relative Area |
|---|---|---|---|
| 1 | α-Pinene | 936 | 3.77 |
| 2 | Sabinene | 956 | 1.16 |
| 3 | α-Myrcene | 985 | 2.00 |
| 5 | 3-Carene | 1010 | 3.70 |
| 6 | Limonene | 1020 | 12.91 |
| 7 | 1,8-Cineol (eucalyptol) | 1033 | 1.20 |
| 8 | γ-Terpinene | 1062 | 0.33 |
| 9 | cis-α-Terpineol | 1078 | 0.20 |
| 10 | Terpinolene | 1086 | 0.41 |
| 11 | p-Menthan-3-one | 1092 | 0.70 |
| 12 | Menthone | 1125 | 0.37 |
| 13 | 1-menthol | 1136 | 1.88 |
| 14 | Carvone | 1142 | 57.93 |
| 15 | 2-isopropyl-5-methyl-3-cyclohexen-1-one | 1156 | 0.20 |
| 16 | p-Menthane | 1263 | 5.88 |
| 17 | Copaene | 1376 | 1.44 |
| 18 | α-Bourbonene | 1385 | 0.61 |
| 19 | Caryophyllene | 1420 | 1.03 |
| N | Compound. | Retention Index | % of Relative Areas |
|---|---|---|---|
| 1 | α-Pinene | 936 | 18.15 |
| 2 | β-Pinene | 983 | 1.78 |
| 3 | α–Myrcene | 985 | 1.56 |
| 4 | 1,8-Cineol (eucalyptol) | 1033 | 65.83 |
| 5 | 4-Terpineol | 1182 | 0.77 |
| 6 | α-Terpinol | 1242 | 2.02 |
| 7 | α-Terpineol acetate | 1330 | 5.14 |
| 8 | (+)-Aromadendrene | 1440 | 2.84 |
| 9 | Globulol | 1590 | 1.92 |
| Essential Oil | Concentration | Diameter ± SD (mm) |
|---|---|---|
| Mentha spicata | 100% | 18.30 ± 0.47 |
| 50% | 15.75 ± 0.41 | |
| 10% | 13.66 ± 0.47 | |
| Eucalyptus globulus | 100% | 27.00 ± 0.82 |
| 50% | 18.00 ± 0.82 | |
| 10% | 12.00 ± 0.82 | |
| Solvent (DMSO) | 0.1% | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landeo-Villanueva, G.E.; Salazar-Salvatierra, M.E.; Ruiz-Quiroz, J.R.; Zuta-Arriola, N.; Jarama-Soto, B.; Herrera-Calderon, O.; Pari-Olarte, J.B.; Loyola-Gonzales, E. Inhibitory Activity of Essential Oils of Mentha spicata and Eucalyptus globulus on Biofilms of Streptococcus mutans in an In Vitro Model. Antibiotics 2023, 12, 369. https://doi.org/10.3390/antibiotics12020369
Landeo-Villanueva GE, Salazar-Salvatierra ME, Ruiz-Quiroz JR, Zuta-Arriola N, Jarama-Soto B, Herrera-Calderon O, Pari-Olarte JB, Loyola-Gonzales E. Inhibitory Activity of Essential Oils of Mentha spicata and Eucalyptus globulus on Biofilms of Streptococcus mutans in an In Vitro Model. Antibiotics. 2023; 12(2):369. https://doi.org/10.3390/antibiotics12020369
Chicago/Turabian StyleLandeo-Villanueva, Guillermo Ernesto, María Elena Salazar-Salvatierra, Julio Reynaldo Ruiz-Quiroz, Noemi Zuta-Arriola, Benjamín Jarama-Soto, Oscar Herrera-Calderon, Josefa Bertha Pari-Olarte, and Eddie Loyola-Gonzales. 2023. "Inhibitory Activity of Essential Oils of Mentha spicata and Eucalyptus globulus on Biofilms of Streptococcus mutans in an In Vitro Model" Antibiotics 12, no. 2: 369. https://doi.org/10.3390/antibiotics12020369
APA StyleLandeo-Villanueva, G. E., Salazar-Salvatierra, M. E., Ruiz-Quiroz, J. R., Zuta-Arriola, N., Jarama-Soto, B., Herrera-Calderon, O., Pari-Olarte, J. B., & Loyola-Gonzales, E. (2023). Inhibitory Activity of Essential Oils of Mentha spicata and Eucalyptus globulus on Biofilms of Streptococcus mutans in an In Vitro Model. Antibiotics, 12(2), 369. https://doi.org/10.3390/antibiotics12020369