
Diversity and Distribution of β-Lactamase Genes Circulating in Indian Isolates of Multidrug-Resistant Klebsiella pneumoniae

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
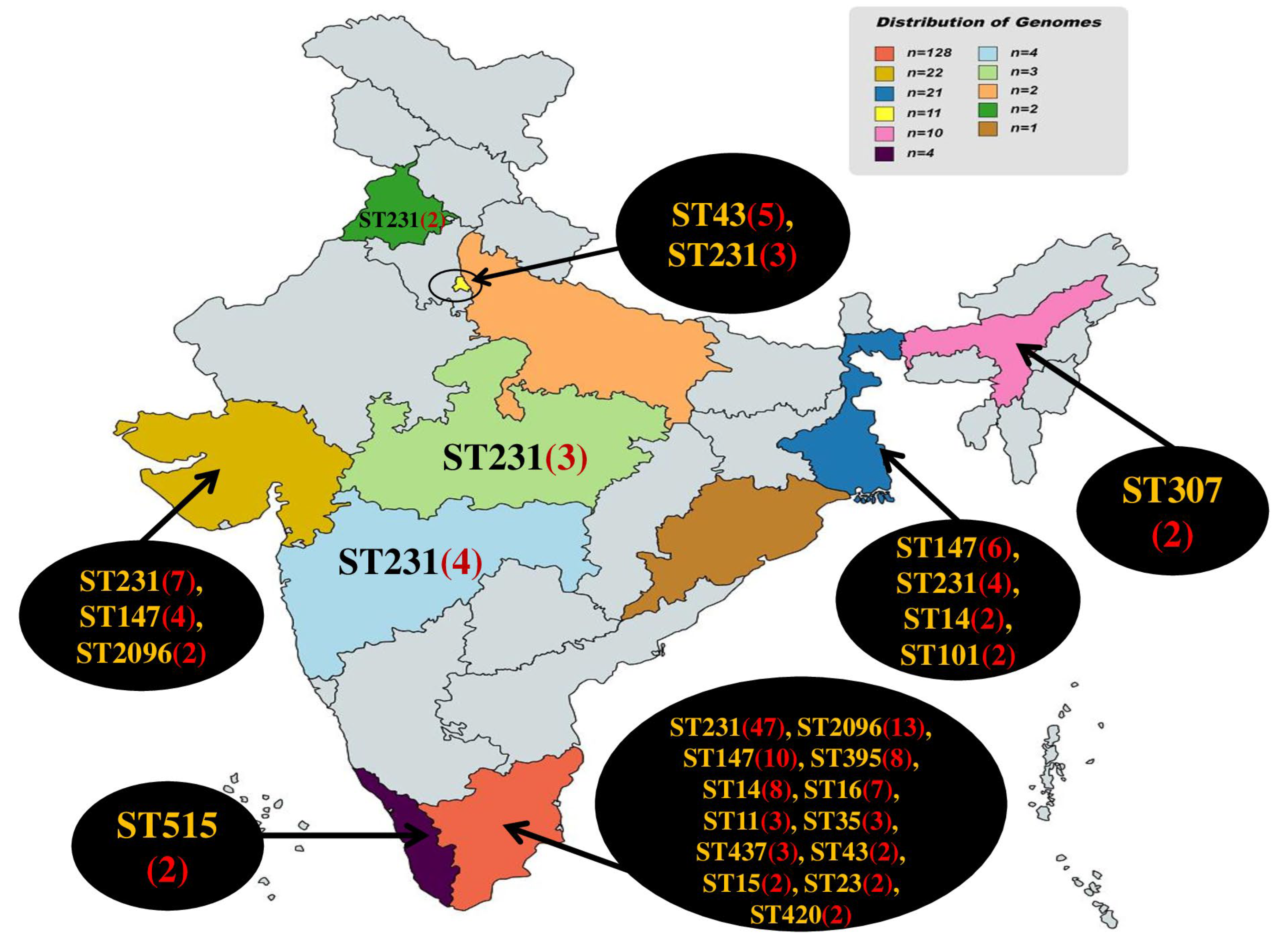
2.1. Distribution of Sequence Types (STs)
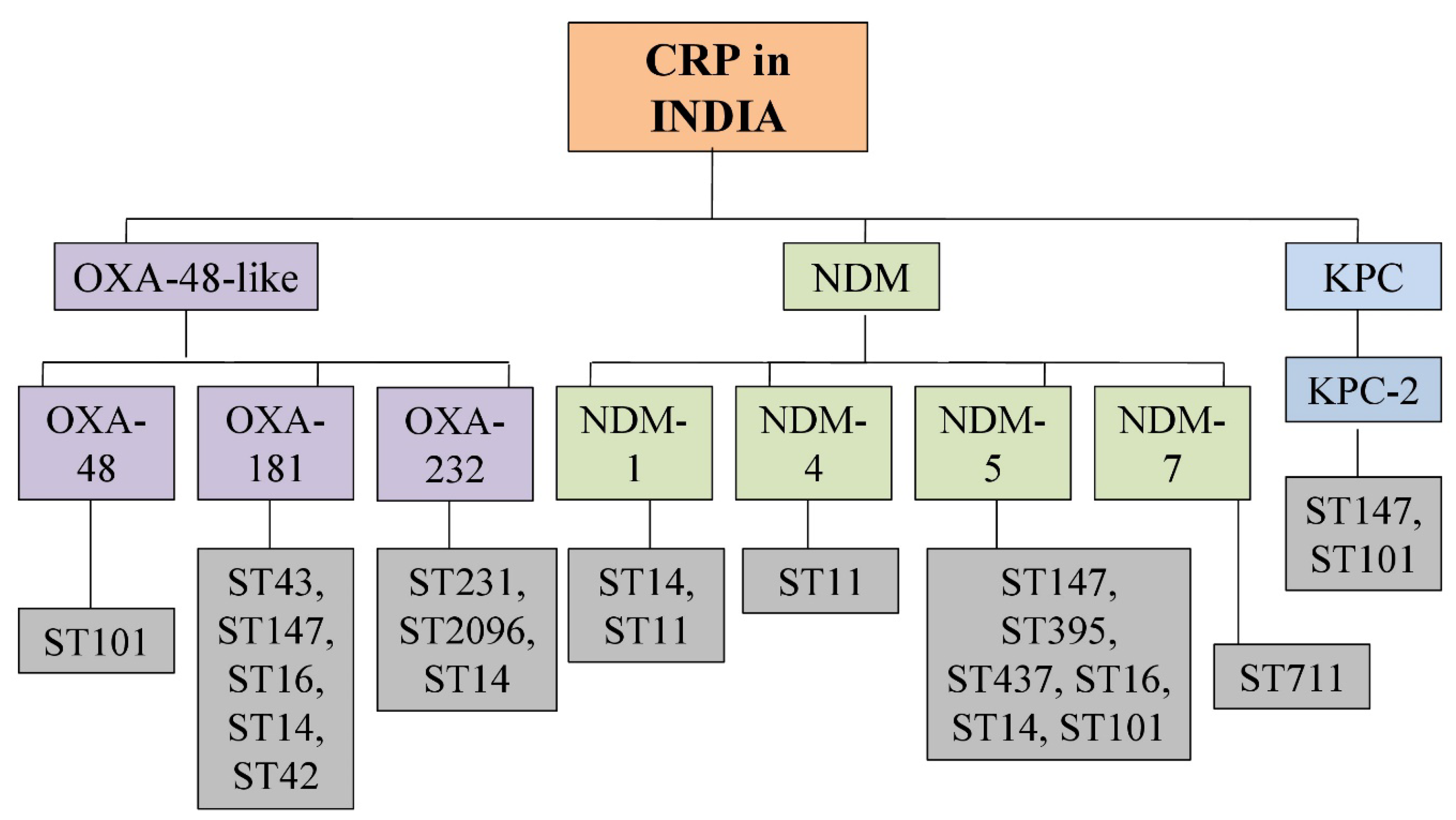
2.2. Distribution of Carbapenemases
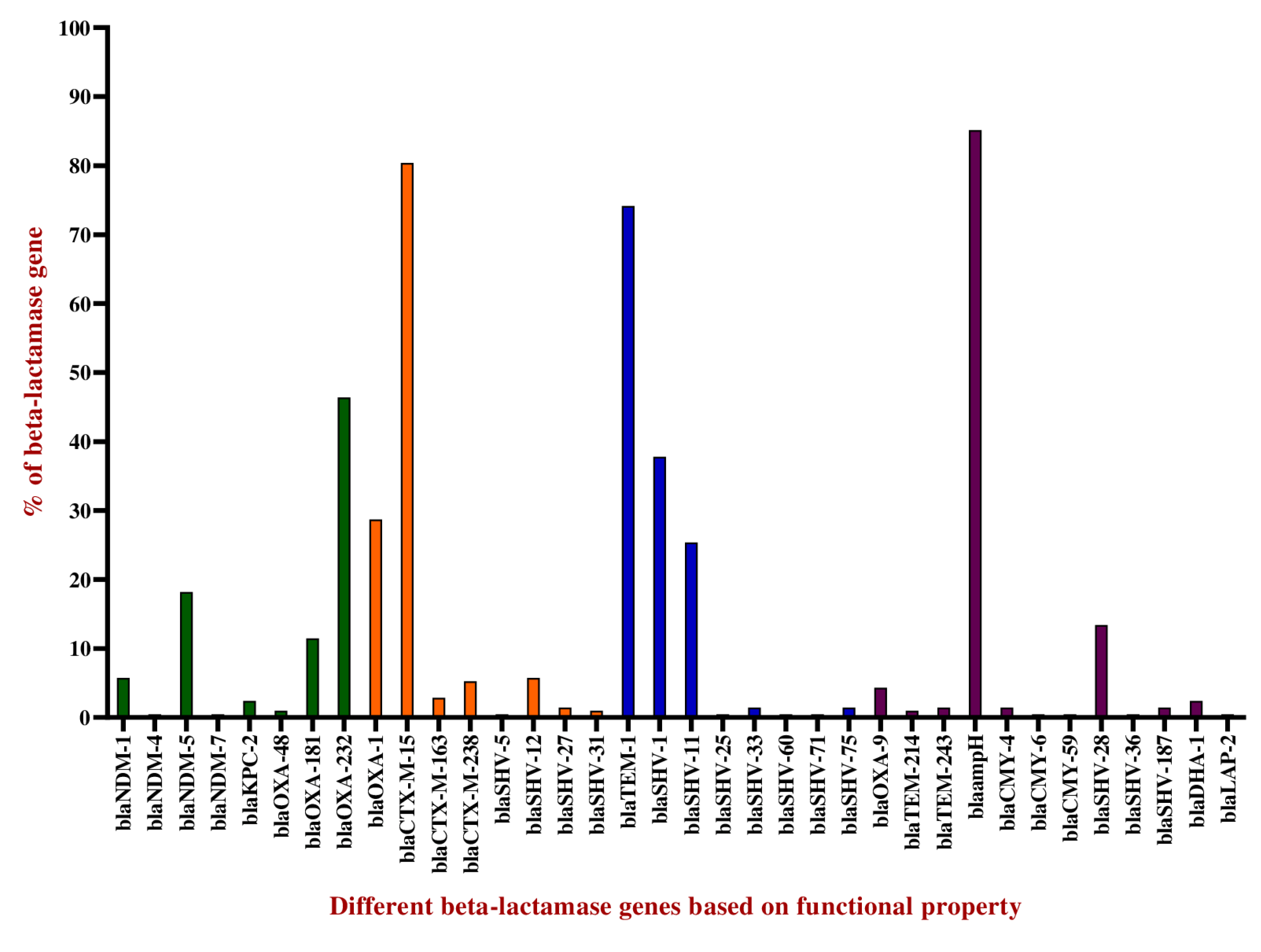
2.3. Distribution of Extended-Spectrum β-Lactamases
2.4. Distribution of Broad-Spectrum β-Lactamases
2.5. Other β-Lactamases
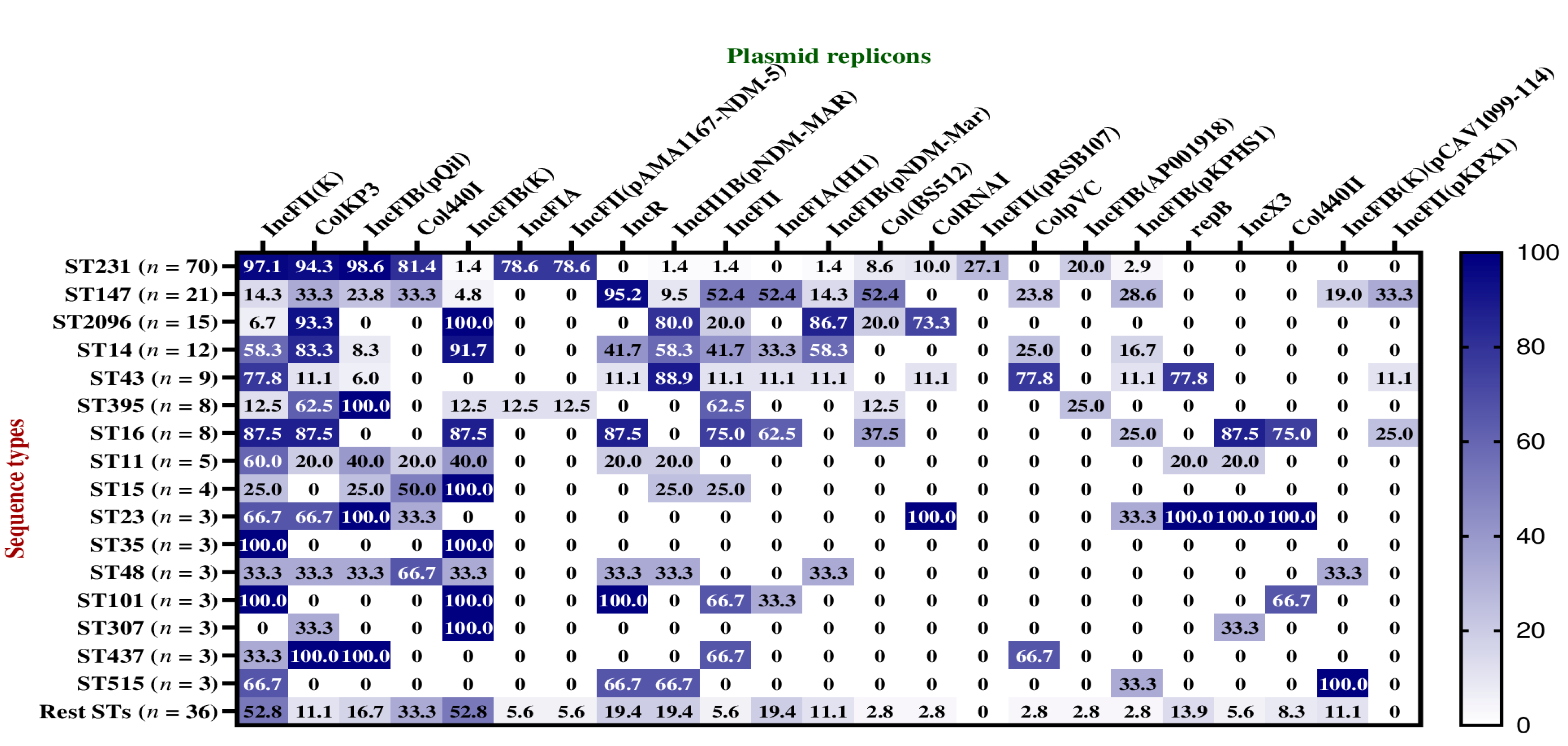
2.6. Plasmid Replicons among Dominant STs
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Identification, and Antibiotic Susceptibility Testing
4.2. Genomic DNA Extraction, NGS Library Preparation, and Whole Genome Sequencing
4.3. Analysis of Whole Genome Sequencing Data
4.4. Retrieval and Analysis of Publicly Available Genomes of Klebsiella Pneumoniae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A Major Worldwide Source and Shuttle for Antibiotic Resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Ho, P.-L.; Cheung, Y.-Y.; Lo, W.-U.; Li, Z.; Chow, K.-H.; Lin, C.-H.; Chan, J.F.-W.; Cheng, V.C.-C. Molecular Characterization of an Atypical IncX3 Plasmid PKPC-NY79 Carrying Bla KPC-2 in a Klebsiella pneumoniae. Curr. Microbiol. 2013, 67, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 13 February 2023).
- WHO Country Office for India, 2021, Indian Priority Pathogen List. Available online: https://cdn.who.int/media/docs/default-source/searo/india/antimicrobial-resistance/ippl_final_web.pdf?sfvrsn=9105c3d1_6 (accessed on 9 February 2023).
- Lee, C.-R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parveen, R.M.; Harish, B.N.; Parija, S.C. Emerging Carbapenem Resistance among Nosocomial Isolates of Klebsiella pneumoniae in South India. Int. J. Pharma Bio Sci. 2010, 1, 1–11. [Google Scholar]
- Shahid, M.; Malik, A.; Akram, M.; Agrawal, L.M.; Khan, A.U.; Agrawal, M. Prevalent Phenotypes and Antibiotic Resistance in Escherichia coli and Klebsiella pneumoniae at an Indian Tertiary Care Hospital: Plasmid-Mediated Cefoxitin Resistance. Int. J. Infect. Dis. 2008, 12, 256–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pragasam, A.K.; Shankar, C.; Veeraraghavan, B.; Biswas, I.; Nabarro, L.E.B.; Inbanathan, F.Y.; George, B.; Verghese, S. Molecular Mechanisms of Colistin Resistance in Klebsiella pneumoniae Causing Bacteremia from India—A First Report. Front. Microbiol. 2017, 7, 2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odsbu, I.; Khedkar, S.; Lind, F.; Khedkar, U.; Nerkar, S.; Orsini, N.; Tamhankar, A.; Stålsby Lundborg, C. Trends in Resistance to Extended-Spectrum Cephalosporins and Carbapenems among Escherichia coli and Klebsiella Spp. Isolates in a District in Western India during 2004–2014. Int. J. Environ. Res. Public Health 2018, 15, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggi, N.; Chatterjee, N.; Singh, V.; Giri, S.K.; Dwivedi, P.; Panwar, R.; Sharma, A.P. Carbapenem Resistance in Escherichia coli and Klebsiella pneumoniae among Indian and International Patients in North India. Acta Microbiol. Immunol. Hung. 2019, 66, 367–376. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Karki, D.; Dhungel, B.; Bhandari, S.; Kunwar, A.; Joshi, P.R.; Shrestha, B.; Rijal, K.R.; Ghimire, P.; Banjara, M.R. Antibiotic Resistance and Detection of Plasmid Mediated Colistin Resistance Mcr-1 Gene among Escherichia coli and Klebsiella pneumoniae Isolated from Clinical Samples. Gut Pathog. 2021, 13, 45. [Google Scholar] [CrossRef]
- Dey, S.; Gaur, M.; Sahoo, R.K.; Das, A.; Jain, B.; Pati, S.; Subudhi, E. Genomic Characterization of XDR Klebsiella pneumoniae ST147 Co-Resistant to Carbapenem and Colistin—The First Report in India. J. Glob. Antimicrob. Resist. 2020, 22, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Desai, S.; Passet, V.; Gajjar, D.; Brisse, S. Genomic Evolution of the Globally Disseminated Multidrug-Resistant Klebsiella pneumoniae Clonal Group 147. Microb. Genom. 2022, 8, 000737. [Google Scholar] [CrossRef] [PubMed]
- Shankar, C.; Jacob, J.J.; Sugumar, S.G.; Natarajan, L.; Rodrigues, C.; Mathur, P.; Mukherjee, D.N.; Sharma, A.; Chitnis, D.S.; Bharagava, A.; et al. Distinctive Mobile Genetic Elements Observed in the Clonal Expansion of Carbapenem-Resistant Klebsiella pneumoniae in India. Microb. Drug Resist. 2021, 27, 1096–1104. [Google Scholar] [CrossRef]
- Sundaresan, A.K.; Vincent, K.; Mohan, G.B.M.; Ramakrishnan, J. Association of Sequence Types, Antimicrobial Resistance and Virulence Genes in Indian Isolates of Klebsiella pneumoniae: A Comparative Genomics Study. J. Glob. Antimicrob. Resist. 2022, 30, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Naha, S.; Sands, K.; Mukherjee, S.; Saha, B.; Dutta, S.; Basu, S. OXA-181-Like Carbapenemases in Klebsiella pneumoniae ST14, ST15, ST23, ST48, and ST231 from Septicemic Neonates: Coexistence with NDM-5, Resistome, Transmissibility, and Genome Diversity. mSphere 2021, 6, e01156-20. [Google Scholar] [CrossRef]
- Shankar, C.; Mathur, P.; Venkatesan, M.; Pragasam, A.K.; Anandan, S.; Khurana, S.; Veeraraghavan, B. Rapidly Disseminating BlaOXA-232 Carrying Klebsiella pneumoniae Belonging to ST231 in India: Multiple and Varied Mobile Genetic Elements. BMC Microbiol. 2019, 19, 137. [Google Scholar] [CrossRef]
- Potron, A.; Rondinaud, E.; Poirel, L.; Belmonte, O.; Boyer, S.; Camiade, S.; Nordmann, P. Genetic and Biochemical Characterisation of OXA-232, a Carbapenem-Hydrolysing Class D β-Lactamase from Enterobacteriaceae. Int. J. Antimicrob. Agents 2013, 41, 325–329. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Ye, J.; Xu, W.; Xu, Y.; Zeng, W.; Liao, W.; Chen, T.; Cao, J.; Wu, Q.; et al. Outbreak of Multidrug-Resistant OXA-232-Producing ST15 Klebsiella pneumoniae in a Teaching Hospital in Wenzhou, China. Infect. Drug Resist. 2021, 14, 4395–4407. [Google Scholar] [CrossRef]
- Abe, R.; Akeda, Y.; Takeuchi, D.; Sakamoto, N.; Sugawara, Y.; Yamamoto, N.; Kerdsin, A.; Matsumoto, Y.; Motooka, D.; Leolerd, W.; et al. Clonal Dissemination of Carbapenem-Resistant Klebsiella pneumoniae ST16 Co-Producing NDM-1 and OXA-232 in Thailand. JAC-Antimicrob. Resist. 2022, 4, dlac084. [Google Scholar] [CrossRef]
- Xanthopoulou, K.; Carattoli, A.; Wille, J.; Biehl, L.M.; Rohde, H.; Farowski, F.; Krut, O.; Villa, L.; Feudi, C.; Seifert, H.; et al. Antibiotic Resistance and Mobile Genetic Elements in Extensively Drug-Resistant Klebsiella pneumoniae Sequence Type 147 Recovered from Germany. Antibiotics 2020, 9, 675. [Google Scholar] [CrossRef]
- Emeraud, C.; Birer, A.; Girlich, D.; Jousset, A.B.; Creton, E.; Naas, T.; Bonnin, R.A.; Dortet, L. Polyclonal Dissemination of OXA-232 Carbapenemase–Producing Klebsiella pneumoniae, France, 2013–2021. Emerg. Infect. Dis. 2022, 28, 2304–2307. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.A.; Landman, F.; de Haan, A.; Witteveen, S.; van Santen-Verheuvel, M.G.; Schouls, L.M.; the Dutch CPE surveillance Study Group. Bla OXA-48-like Genome Architecture among Carbapenemase-Producing Escherichia coli and Klebsiella pneumoniae in the Netherlands. Microb. Genom. 2021, 7, 000512. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.-A.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, e00102-19. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Huh, H.J.; Baek, J.Y.; Chung, N.Y.; Ryu, J.G.; Ki, C.-S.; Chung, D.R.; Lee, N.Y.; Song, J.-H. Klebsiella pneumoniae Co-Producing NDM-5 and OXA-181 Carbapenemases, South Korea. Emerg. Infect. Dis. 2015, 21, 1088–1089. [Google Scholar] [CrossRef] [PubMed]
- Avolio, M.; Vignaroli, C.; Crapis, M.; Camporese, A. Co-Production of NDM-1 and OXA-232 by ST16 Klebsiella pneumoniae, Italy, 2016. Future Microbiol. 2017, 12, 1119–1122. [Google Scholar] [CrossRef]
- Shibl, A.; Al-Agamy, M.; Memish, Z.; Senok, A.; Khader, S.A.; Assiri, A. The Emergence of OXA-48- and NDM-1-Positive Klebsiella pneumoniae in Riyadh, Saudi Arabia. Int. J. Infect. Dis. 2013, 17, e1130–e1133. [Google Scholar] [CrossRef] [Green Version]
- Solgi, H.; Nematzadeh, S.; Giske, C.G.; Badmasti, F.; Westerlund, F.; Lin, Y.-L.; Goyal, G.; Nikbin, V.S.; Nemati, A.H.; Shahcheraghi, F. Molecular Epidemiology of OXA-48 and NDM-1 Producing Enterobacterales Species at a University Hospital in Tehran, Iran, Between 2015 and 2016. Front. Microbiol. 2020, 11, 936. [Google Scholar] [CrossRef]
- Chaalal, N.; Touati, A.; Bakour, S.; Aissa, M.A.; Sotto, A.; Lavigne, J.-P.; Pantel, A. Spread of OXA-48 and NDM-1-Producing Klebsiella pneumoniae ST48 and ST101 in Chicken Meat in Western Algeria. Microb. Drug Resist. 2021, 27, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Rojas, L.J.; Hujer, A.M.; Rudin, S.D.; Wright, M.S.; Domitrovic, T.N.; Marshall, S.H.; Hujer, K.M.; Richter, S.S.; Cober, E.; Perez, F.; et al. NDM-5 and OXA-181 Beta-Lactamases, a Significant Threat Continues To Spread in the Americas. Antimicrob. Agents Chemother. 2017, 61, e00454-17. [Google Scholar] [CrossRef] [Green Version]
- Sherchan, J.B.; Tada, T.; Shrestha, S.; Uchida, H.; Hishinuma, T.; Morioka, S.; Shahi, R.K.; Bhandari, S.; Twi, R.T.; Kirikae, T.; et al. Emergence of Clinical Isolates of Highly Carbapenem-Resistant Klebsiella pneumoniae Co-Harboring BlaNDM-5 and BlaOXA-181 or -232 in Nepal. Int. J. Infect. Dis. 2020, 92, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safavi, M.; Bostanshirin, N.; Hajikhani, B.; Yaslianifard, S.; van Belkum, A.; Goudarzi, M.; Hashemi, A.; Darban-Sarokhalil, D.; Dadashi, M. Global Genotype Distribution of Human Clinical Isolates of New Delhi Metallo-β-Lactamase-Producing Klebsiella pneumoniae; A Systematic Review. J. Glob. Antimicrob. Resist. 2020, 23, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Giske, C.G.; Fröding, I.; Hasan, C.M.; Turlej-Rogacka, A.; Toleman, M.; Livermore, D.; Woodford, N.; Walsh, T.R. Diverse Sequence Types of Klebsiella pneumoniae Contribute to the Dissemination of BlaNDM-1 in India, Sweden, and the United Kingdom. Antimicrob. Agents Chemother. 2012, 56, 2735–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.; Hopkins, K.L.; Turton, J.; Doumith, M.; Hill, R.; Loy, R.; Meunier, D.; Pike, R.; Livermore, D.M.; Woodford, N. NDM Carbapenemases in the United Kingdom: An Analysis of the First 250 Cases. J. Antimicrob. Chemother. 2014, 69, 1777–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, E.-J.; Kang, D.Y.; Yang, J.W.; Kim, D.; Lee, H.; Lee, K.J.; Jeong, S.H. New Delhi Metallo-Beta-Lactamase-Producing Enterobacteriaceae in South Korea Between 2010 and 2015. Front. Microbiol. 2018, 9, 571. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Wei, Z.; Ji, S.; Du, X.; Shen, P.; Yu, Y. ST11, the Dominant Clone of KPC-Producing Klebsiella pneumoniae in China. J. Antimicrob. Chemother. 2011, 66, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Baraniak, A.; Izdebski, R.; Fiett, J.; Gawryszewska, I.; Bojarska, K.; Herda, M.; Literacka, E.; Żabicka, D.; Tomczak, H.; Pewińska, N.; et al. NDM-Producing Enterobacteriaceae in Poland, 2012–14: Inter-Regional Outbreak of Klebsiella pneumoniae ST11 and Sporadic Cases. J. Antimicrob. Chemother. 2016, 71, 85–91. [Google Scholar] [CrossRef] [Green Version]
- M100Ed32|Performance Standards for Antimicrobial Susceptibility Testing, 32nd Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 15 February 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Resistance Category | Genome Accession No. | Strain | Resistance Category | Genome Accession No. |
|---|---|---|---|---|---|
| SBS1 | XDR | JAFFRJ000000000 | DGL8 | PDR | JAJBAG000000000 |
| SBS3 | XDR | JAFFRI000000000 | DGL9 | XDR | JAJBAF000000000 |
| SBS4 | MDR | JAFFRH000000000 | DGL10 | XDR | JAJBAE000000000 |
| SBS5 | XDR | JAFFRG000000000 | DGL11 | XDR | JAJBAD000000000 |
| SBS11 | MDR | JAFFRA000000000 | DGL12 | XDR | JAJBAC000000000 |
| SBS12 | PDR | JAFFQZ000000000 | DGL13 | XDR | JAJBAB000000000 |
| DGL1 | MDR | JAJBAN000000000 | DGL14 | XDR | JAJBAA000000000 |
| DGL2 | MDR | JAJBAM000000000 | DGL15 | XDR | JAJAZZ000000000 |
| DGL5 | PDR | JAJBAJ000000000 | DGL16 | XDR | JAJAZY000000000 |
| DGL6 | XDR | JAJBAI000000000 | DGL17 | XDR | JAJAZX000000000 |
| DGL7 | XDR | JAJBAH000000000 | DGL18 | XDR | JAJAZW000000000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shukla, S.; Desai, S.; Bagchi, A.; Singh, P.; Joshi, M.; Joshi, C.; Patankar, J.; Maheshwari, G.; Rajni, E.; Shah, M.; et al. Diversity and Distribution of β-Lactamase Genes Circulating in Indian Isolates of Multidrug-Resistant Klebsiella pneumoniae. Antibiotics 2023, 12, 449. https://doi.org/10.3390/antibiotics12030449
Shukla S, Desai S, Bagchi A, Singh P, Joshi M, Joshi C, Patankar J, Maheshwari G, Rajni E, Shah M, et al. Diversity and Distribution of β-Lactamase Genes Circulating in Indian Isolates of Multidrug-Resistant Klebsiella pneumoniae. Antibiotics. 2023; 12(3):449. https://doi.org/10.3390/antibiotics12030449
Chicago/Turabian StyleShukla, Suraj, Siddhi Desai, Ashutosh Bagchi, Pushpendra Singh, Madhvi Joshi, Chaitanya Joshi, Jyoti Patankar, Geeti Maheshwari, Ekadashi Rajni, Manali Shah, and et al. 2023. "Diversity and Distribution of β-Lactamase Genes Circulating in Indian Isolates of Multidrug-Resistant Klebsiella pneumoniae" Antibiotics 12, no. 3: 449. https://doi.org/10.3390/antibiotics12030449
APA StyleShukla, S., Desai, S., Bagchi, A., Singh, P., Joshi, M., Joshi, C., Patankar, J., Maheshwari, G., Rajni, E., Shah, M., & Gajjar, D. (2023). Diversity and Distribution of β-Lactamase Genes Circulating in Indian Isolates of Multidrug-Resistant Klebsiella pneumoniae. Antibiotics, 12(3), 449. https://doi.org/10.3390/antibiotics12030449