

α-Pinene: Docking Study, Cytotoxicity, Mechanism of Action, and Anti-Biofilm Effect against Candida albicans
 , , , ,
, , , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
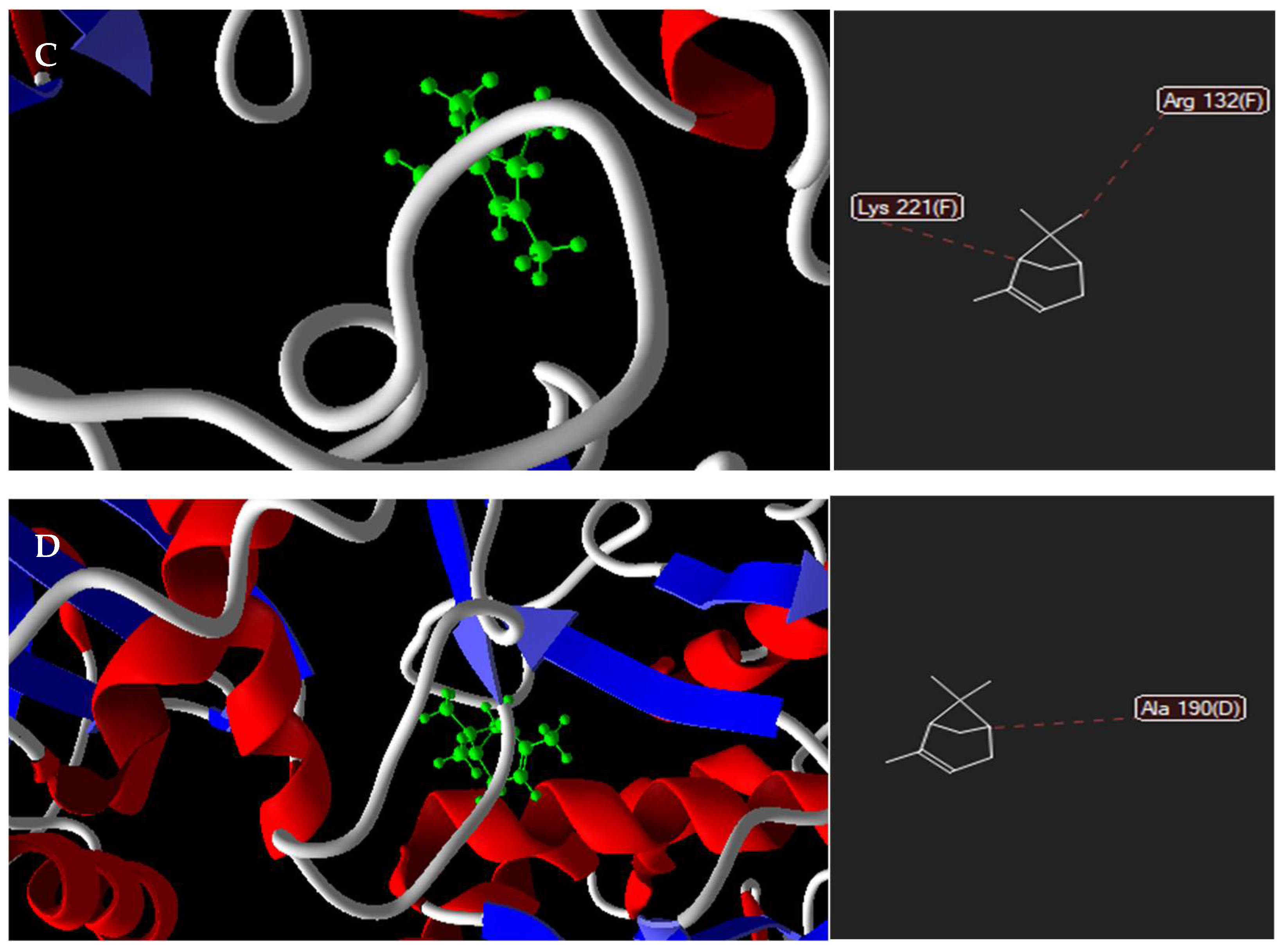
2.1. Docking Prediction
2.2. Sorbitol and Exogenous-Ergosterol Assays
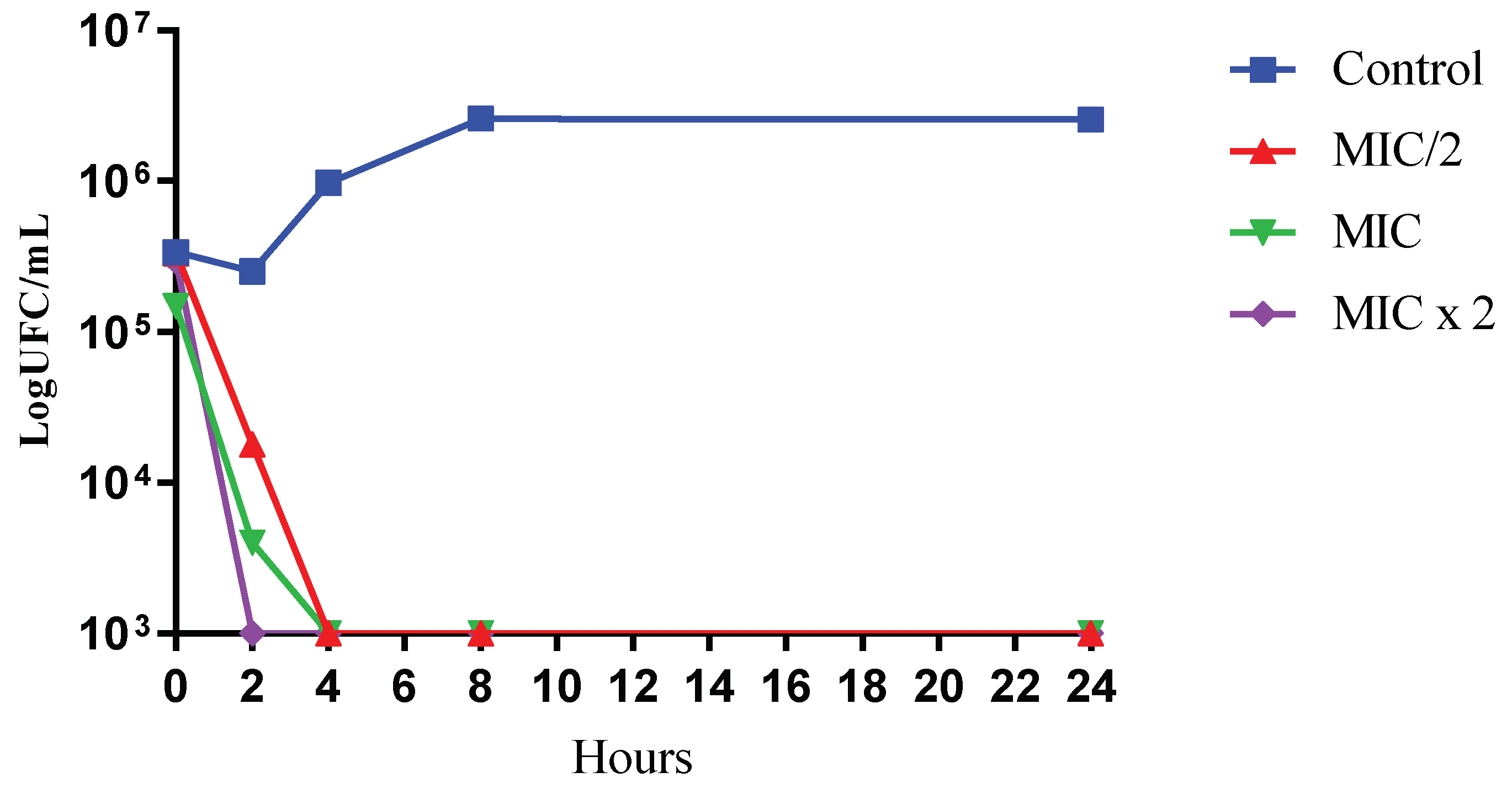
2.3. Time-Kill Assay
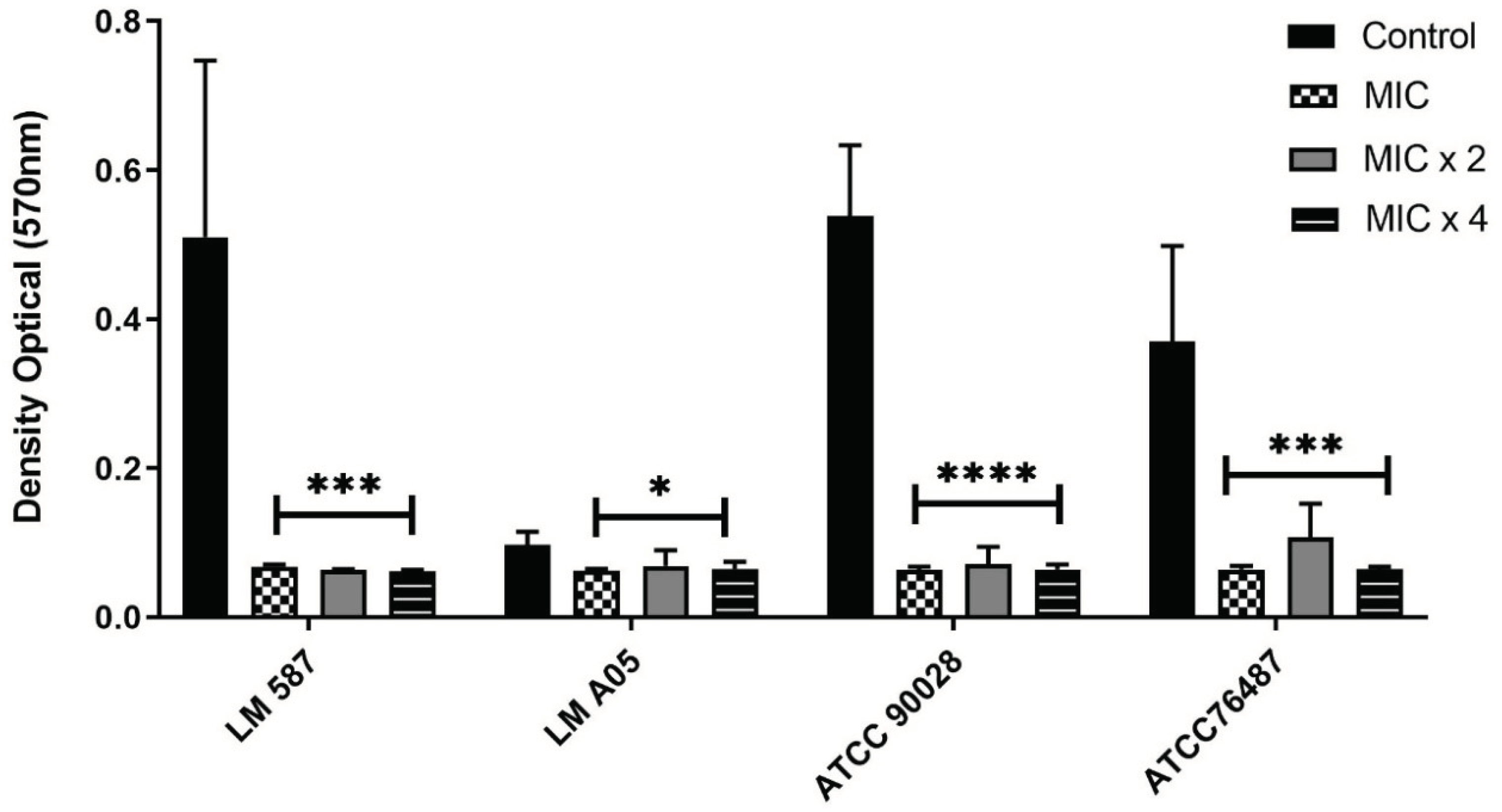
2.4. α-Pinene Anti-Biofilm Activity
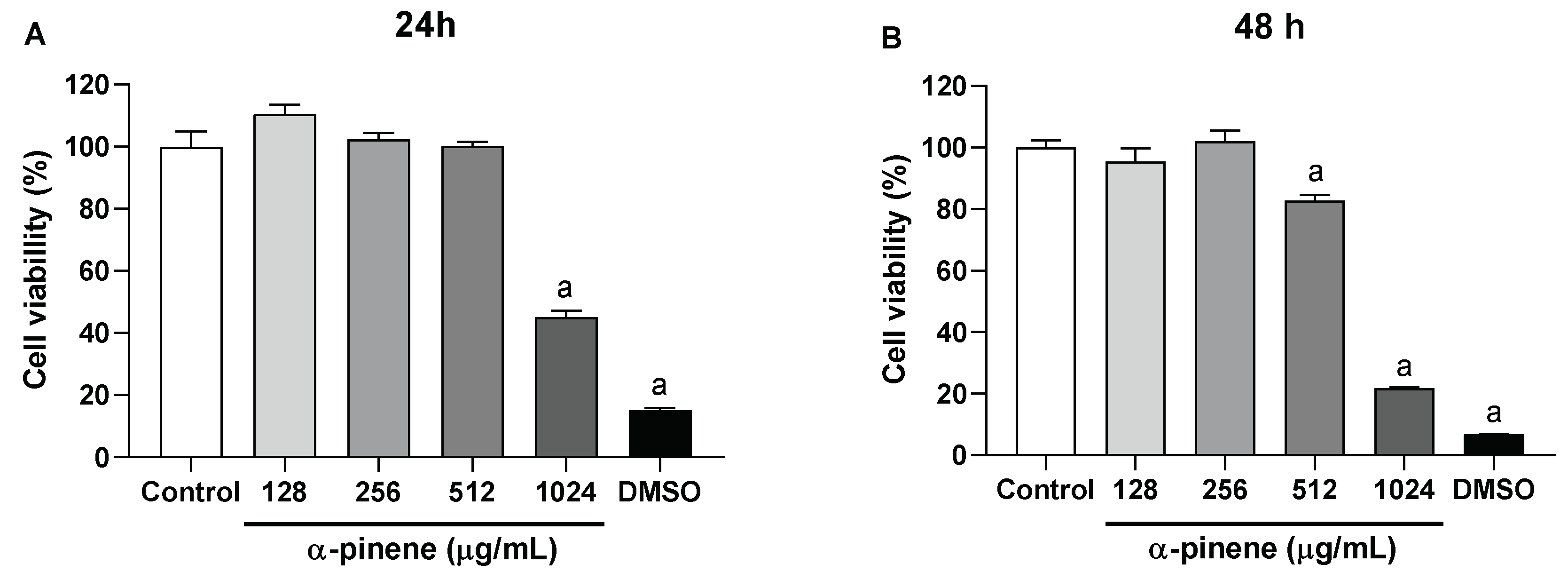
2.5. Cytotoxicity Assay against HaCaT Cells
3. Discussion
4. Materials and Methods
4.1. Test Substance, Antifungal Drugs, and Culture Media
4.2. Microorganisms
4.3. Molecular Docking
4.4. Mechanism of Action
4.4.1. Effect on the Fungal Cell Wall
4.4.2. Ergosterol Binding Assay
4.5. Time-Kill
4.6. Biofilm Eradication
4.7. Cytotoxicity Assay against HaCaT Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendonça, A.; Santos, H.; Franco-Duarte, R.; Sampaio, P. Fungal Infections Diagnosis—Past, Present and Future. Res. Microbiol. 2022, 173, 103915. [Google Scholar] [CrossRef] [PubMed]
- Nami, S.; Mohammadi, R.; Vakili, M.; Khezripour, K.; Mirzaei, H.; Morovati, H. Fungal Vaccines, Mechanism of Actions and Immunology: A Comprehensive Review. Biomed. Pharmacother. 2019, 109, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, T.; Kawano, T.; Yamashita, K.; Yamada, E.; Fujino, N.; Kaeriyama, M.; Fukuda, Y.; Nomura, N.; Mitsuyama, J.; Suematsu, H.; et al. Antifungal Susceptibility Trend and Analysis of Resistance Mechanism for Candida Species Isolated from Bloodstream at a Japanese University Hospital. J. Infect. Chemother. 2019, 25, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Pereira, R.; dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida Albicans: Formation, Regulation and Resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Armstrong-James, D.; Brown, G.D.; Netea, M.G.; Zelante, T.; Gresnigt, M.S.; van de Veerdonk, F.L.; Levitz, S.M. Immunotherapeutic Approaches to Treatment of Fungal Diseases. Lancet Infect. Dis. 2017, 17, e393–e402. [Google Scholar] [CrossRef]
- Shields, R.K.; Kline, E.G.; Healey, K.R.; Kordalewska, M.; Perlin, D.S.; Hong Nguyen, M.; Clancy, C.J. Spontaneous Mutational Frequency and Fks Mutation Rates Vary by Echinocandin Agent against Candida Glabrata. Antimicrob. Agents Chemother. 2019, 63, e01692-18. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; WHO: Geneva, Switzerland, 2022; ISBN 9789240060241. [Google Scholar]
- da Raimundo, J.S.; de Toledo, C.E.M. Plantas Com Atividade Antifúngica No Tratamento Da Candidíase: Uma Revisão Bibliográfica Plants With Antifungal Activity in the Treatment of Candidiasis. Rev. UNINGÁ Rev. 2017, 29, 75–80. [Google Scholar]
- Bohner, F.; Papp, C.; Gácser, A. The Effect of Antifungal Resistance Development on the Virulence of Candida Species. FEMS Yeast Res. 2022, 22, foac019. [Google Scholar] [CrossRef]
- Cortés, J.A.; Ruiz, J.F.; Melgarejo-Moreno, L.N. Candidemia En Colombia. Biomédica 2020, 40, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Tortorano, A.M.; Prigitano, A.; Morroni, G.; Brescini, L.; Barchiesi, F. Candidemia: Evolution of Drug Resistance and Novel Therapeutic Approaches. Infect. Drug Resist. 2021, 14, 5543–5553. [Google Scholar] [CrossRef]
- Qadri, H.; Haseeb Shah, A.; Mudasir Ahmad, S.; Alshehri, B.; Almilaibary, A.; Ahmad Mir, M. Natural Products and Their Semi-Synthetic Derivatives against Antimicrobial-Resistant Human Pathogenic Bacteria and Fungi. Saudi J. Biol. Sci. 2022, 29, 103376. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.C.R.; Lopes, P.M.; de Azevedo, M.M.B.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Activities of A-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, A.M.; Carlos, A.; Pignatari, C.; Edmond, M.B.; Marra, R.; Fernando, L.; Camargo, A.; Siqueira, R.A. Epidemiology and Microbiologic Characterization of Nosocomial Candidemia from a Brazilian National Surveillance Program. PLoS ONE 2016, 11, e0146909. [Google Scholar] [CrossRef]
- Viegas, C., Jr.; da Bolzani, V.S.; Barreiro, E.J. Os Produtos Naturais e a Química Medicinal Moderna. Quim. Nova 2006, 29, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Upadhyay, S.; Orhan, I.E.; Jugran, A.K.; Baghalpour, N.; Cho, W.C.; Sharifi-Rad, J. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Park, B.B.; An, J.Y.; Park, S.U. Recent Studies on Pinene and Its Biological and Pharmacological Activities. EXCLI J. 2021, 20, 812–818. [Google Scholar] [CrossRef]
- Allenspach, M.; Steuer, C. α-Pinene: A Never-Ending Story. Phytochemistry 2021, 190, 112857. [Google Scholar] [CrossRef]
- de Barros, D.B.; e Lima, L.D.O.; Silva, L.A.; Fonseca, M.C.; Diniz-Neto, H.; da Silva Rocha, W.P.; de Medeiros Beltrão, G.V.; Castellano, L.R.C.; Guerra, F.Q.S.; da Silva, M.V. Antifungal Effect of α-Pinene Alone and in Association with Antifungals against Candida Albicans Strains Efecto. Res. Soc. Dev. 2022, 11, e58711427748. [Google Scholar]
- da Alves, D.N.; Monteiro, A.F.M.; Andrade, P.N.; Lazarini, J.G.; Abílio, G.M.F.; Guerra, F.Q.S.; Scotti, M.T.; Scotti, L.; Rosalen, P.L.; de Castro, R.D. Docking Prediction, Antifungal Activity, Anti-Biofilm Effects on Candida Spp., and Toxicity against Human Cells of Cinnamaldehyde. Molecules 2020, 25, 5969. [Google Scholar] [CrossRef]
- Ferreira, E.S.; Cordeiro, L.V.; Silva, D.F.; Souza, H.D.S.; Lima, E.O.; Castro, R.D.D.E. Antifungal Activity and Mechanism of Action of 2-Chloro- N -Phenylacetamide: A New Molecule with Activity against Strains of Aspergillus Fl Avus. An. Acad. Bras. Ciências 2021, 93, 1–15. [Google Scholar] [CrossRef]
- Vieira, A.J.H.; Santos, J.I. Mecanismos de Resistência de Candida Albicans Aos Antifúngicos Anfotericina B, Fluconazol e Caspofungina. RBAC 2017, 49, 235–239. [Google Scholar] [CrossRef]
- Clerya, M.; Leite, A.; Parente, A.; Bezerra, D.B.; De Sousa, J.P.; Queiroga, F.; Guerra, S.; Lima, E.D.O. Evaluation of Antifungal Activity and Mechanism of Action of Citral against Candida Albicans. Evid. Based Complement. Altern. Med. 2014, 2014, 378280. [Google Scholar] [CrossRef] [Green Version]
- Saracino, I.M.; Foschi, C.; Pavoni, M.; Spigarelli, R.; Valerii, M.C.; Spisni, E. Antifungal Activity of Natural Compounds vs. Candida Spp.: A Mixture of Cinnamaldehyde and Eugenol Shows Promising In Vitro Results. Antibiotics 2022, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Mowat, E.; Jones, B.; Williams, C.; Lopez-ribot, J. Our Current Understanding of Fungal Biofilms. Crit. Rev. Microbiol. 2009, 35, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Rangel, L.; Luiz, P.; Lacet, G.; Ferreira, S.; Almeida, I.; Galbiatti, F.; Carvalho, D.; Roberto, L.; Dias, R.; Castro, D. Archives of Oral Biology Antifungal Activity, Mode of Action and Anti-Bio Fi Lm Effects of Laurus Nobilis Linnaeus Essential Oil against Candida spp. Arch. Oral Biol. 2017, 73, 179–185. [Google Scholar] [CrossRef]
- Seo, M.; Kang, T.J.; Lee, C.H.; Lee, A.; Noh, M. HaCaT Keratinocytes and Primary Epidermal Keratinocytes Have Different Transcriptional Profiles of Cornified Envelope- Associated Genes to T Helper Cell Cytokines. Biomol. Ther. 2012, 20, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikeyan, R.; Kanimozhi, G.; Prasad, N.R.; Agilan, B.; Ganesan, M.; Srithar, G. Alpha Pinene Modulates UVA-Induced Oxidative Stress, DNA Damage and Apoptosis in Human Skin Epidermal Keratinocytes; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 212, ISBN 9842305384. [Google Scholar]
- Karthikeyan, R.; Kanimozhi, G.; Madahavan, N.R.; Agilan, B.; Ganesan, M.; Prasad, N.R.; Rathinaraj, P. Alpha-Pinene Attenuates UVA-Induced Photoaging through Inhibition of Matrix Metalloproteinases Expression in Mouse Skin; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 217, ISBN 9842305384. [Google Scholar]
- Hadacek, F.; Greger, H. Testing of Antifungal Natural Products: Methodologies, Comparability of Results and Assay Choice. Phytochem. Anal. 2000, 11, 137–147. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 3rd ed.; Approved Standard; CLSI: Wayne, PA, USA, 2008; Volume 28, p. 13. [Google Scholar]
- Strushkevich, N.; Usanov, S.A.; Park, H.W. Structural Basis of Human CYP51 Inhibition by Antifungal Azoles. J. Mol. Biol. 2010, 397, 1067–1078. [Google Scholar] [CrossRef]
- Li, X.; Roberti, R.; Blobel, G. Structure of an Integral Membrane Sterol Reductase from Methylomicrobium Alcaliphilum. Nature 2015, 517, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Garlatti, V.; Belloy, N.; Martin, L.; Lacroix, M.; Matsushita, M.; Endo, Y.; Fujita, T.; Fontecilla-Camps, J.C.; Arlaud, G.J.; Thielens, N.M.; et al. Structural Insights into the Innate Immune Recognition Specificities of L- and H-Ficolins. EMBO J. 2007, 26, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Harshbarger, W. Crystal Structures of Binary and Ternary Complexes of Thymidylate Synthase (ThyA) from Mycobacterium Tuberculosis: Insights into Selectivity and Inhibition. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2012. [Google Scholar]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. NIH Public Access. J. Comput. Chem. 2010, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedretti, A.; Villa, L.; Vistoli, G. VEGA—An Open Platform to Develop Chemo-Bio-Informatics Applications, Using Plug-in Architecture and Script Programming. J. Comput. Aided Mol. Des. 2004, 18, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Frost, D.J.; Brandt, K.D.; Cugier, D.; Goldman, R. A Whole-Cell Candida Albicans Assay for the Detection of Inhibitors towards Fungal Cell Wall Synthesis and Assembly. J. Antibiot. 1995, 48, 306–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klepser, M.E.; Ernst, E.J.; Lewis, R.E.; Ernst, M.E.; Pfaller, M.A. Influence of Test Conditions on Antifungal Time-Kill Curve Results: Proposal for Standardized Methods. Antimicrob. Agents Chemother. 1998, 42, 1207–1212. [Google Scholar] [CrossRef] [Green Version]
- Klepser, M.E.; Malone, D.; Lewis, R.E.; Ernst, E.J. Evaluation of Voriconazole Pharmacodynamics Using Time-Kill Methodology. Antimicrob. Agents Chemother. 2000, 44, 1917–1920. [Google Scholar] [CrossRef] [Green Version]
- Rajasekharan, S.K.; Ramesh, S.; Satish, A.S.; Lee, J. Antibiofilm and Anti- β -Lactamase Activities of Burdock Root Extract and Chlorogenic Acid against Klebsiella Pneumoniae. J. Microbiol. Biotechnol. 2017, 27, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. lmmunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | ||||
|---|---|---|---|---|
| 14-α-Demethylase (3JUV) | δ-14-Sterol Reductase (4QUV) | 1,3-β-Glucan Synthase (2J0Y) | Thimidylate Synthase (3QJ7) | |
| Energy (kcal/mol) | −16 | −44 | −32 | −52 |
| Drugs | Sorbitol | Ergosterol | ||
|---|---|---|---|---|
| Absence | Presence | Absence | Presence | |
| α-pinene | 128 | 128 | 128 | >512 |
| Caspofungin | 0.062 | 2 | NA | NA |
| Amphotericin B | NA | NA | 1.25 | >512 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bomfim de Barros, D.; de Oliveira e Lima, L.; Alves da Silva, L.; Cavalcante Fonseca, M.; Ferreira, R.C.; Diniz Neto, H.; da Nóbrega Alves, D.; da Silva Rocha, W.P.; Scotti, L.; de Oliveira Lima, E.; et al. α-Pinene: Docking Study, Cytotoxicity, Mechanism of Action, and Anti-Biofilm Effect against Candida albicans. Antibiotics 2023, 12, 480. https://doi.org/10.3390/antibiotics12030480
Bomfim de Barros D, de Oliveira e Lima L, Alves da Silva L, Cavalcante Fonseca M, Ferreira RC, Diniz Neto H, da Nóbrega Alves D, da Silva Rocha WP, Scotti L, de Oliveira Lima E, et al. α-Pinene: Docking Study, Cytotoxicity, Mechanism of Action, and Anti-Biofilm Effect against Candida albicans. Antibiotics. 2023; 12(3):480. https://doi.org/10.3390/antibiotics12030480
Chicago/Turabian StyleBomfim de Barros, Daniela, Luanna de Oliveira e Lima, Larissa Alves da Silva, Mariana Cavalcante Fonseca, Rafael Carlos Ferreira, Hermes Diniz Neto, Danielle da Nóbrega Alves, Walicyranison Plinio da Silva Rocha, Luciana Scotti, Edeltrudes de Oliveira Lima, and et al. 2023. "α-Pinene: Docking Study, Cytotoxicity, Mechanism of Action, and Anti-Biofilm Effect against Candida albicans" Antibiotics 12, no. 3: 480. https://doi.org/10.3390/antibiotics12030480
APA StyleBomfim de Barros, D., de Oliveira e Lima, L., Alves da Silva, L., Cavalcante Fonseca, M., Ferreira, R. C., Diniz Neto, H., da Nóbrega Alves, D., da Silva Rocha, W. P., Scotti, L., de Oliveira Lima, E., Vieira Sobral, M., Cançado Castellano, L. R., Moura-Mendes, J., Queiroga Sarmento Guerra, F., & da Silva, M. V. (2023). α-Pinene: Docking Study, Cytotoxicity, Mechanism of Action, and Anti-Biofilm Effect against Candida albicans. Antibiotics, 12(3), 480. https://doi.org/10.3390/antibiotics12030480