
The Characterization and Beta-Lactam Resistance of Staphylococcal Community Recovered from Raw Bovine Milk

Abstract
:1. Introduction
2. Results
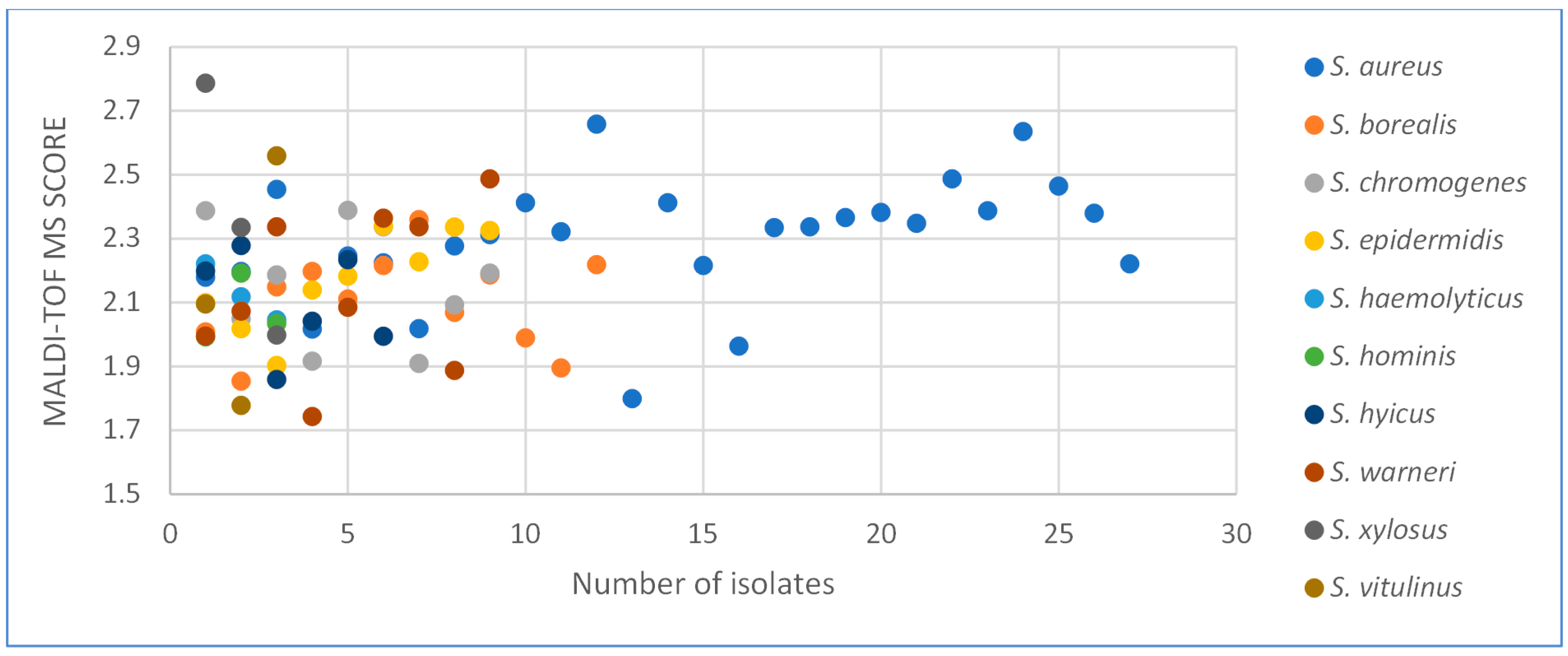
2.1. MALDI-TOF MS and Sequence Analysis
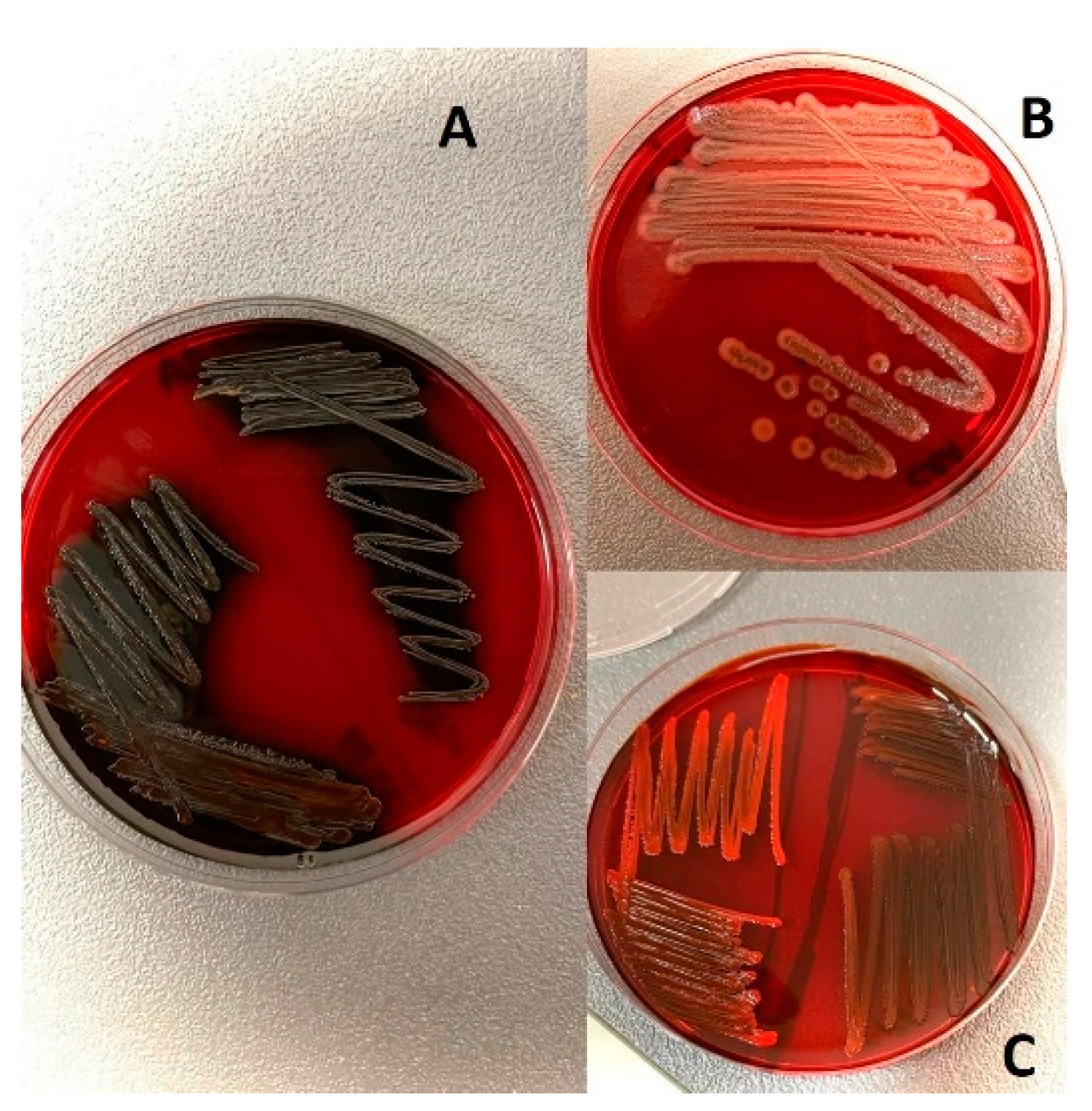
2.2. Evaluation of Hemolysis, Biofilm, and Coagulase Ability
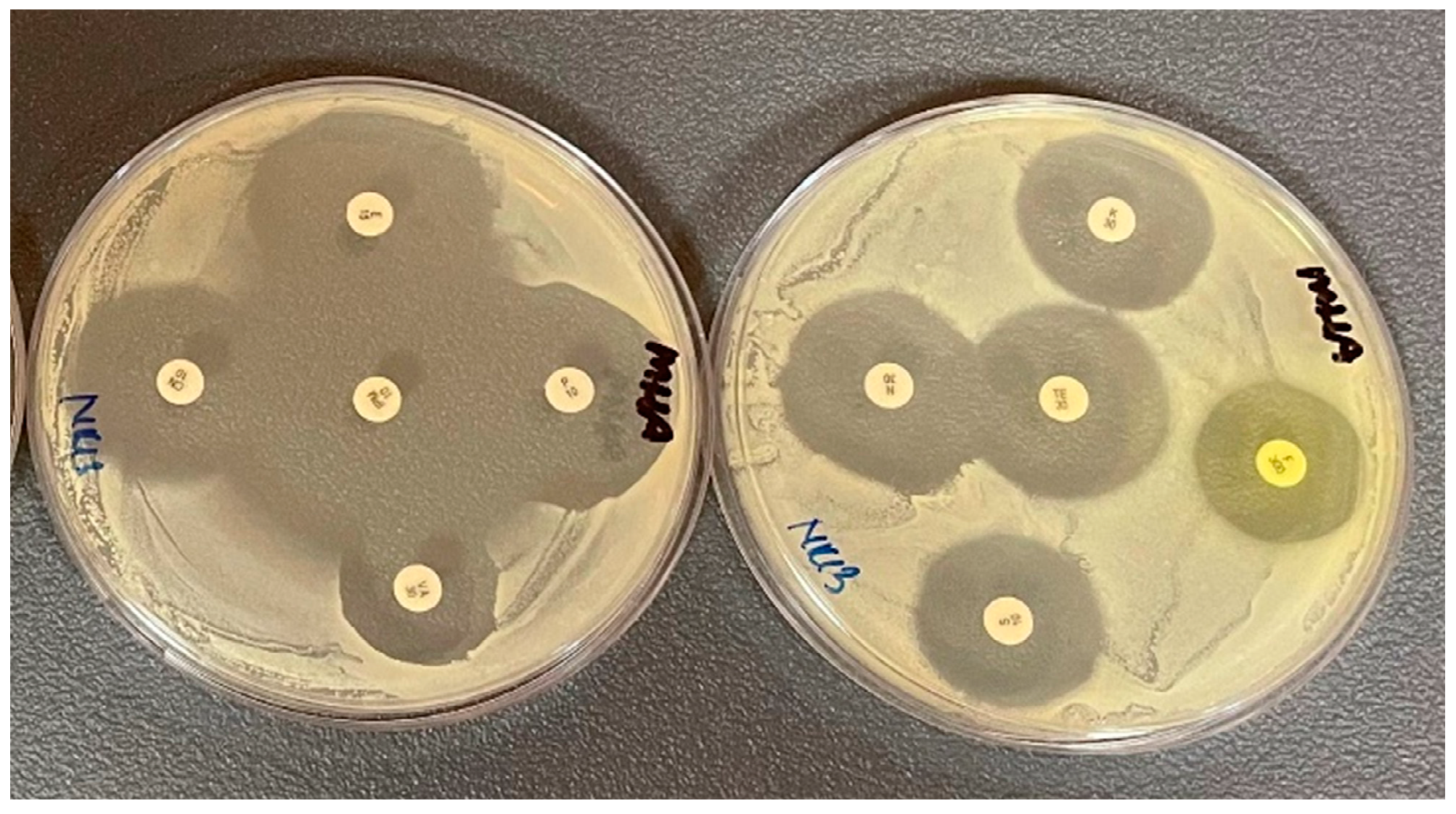
2.3. Antibiotic Resistance Profiles
2.4. Beta-Lactamase Screening
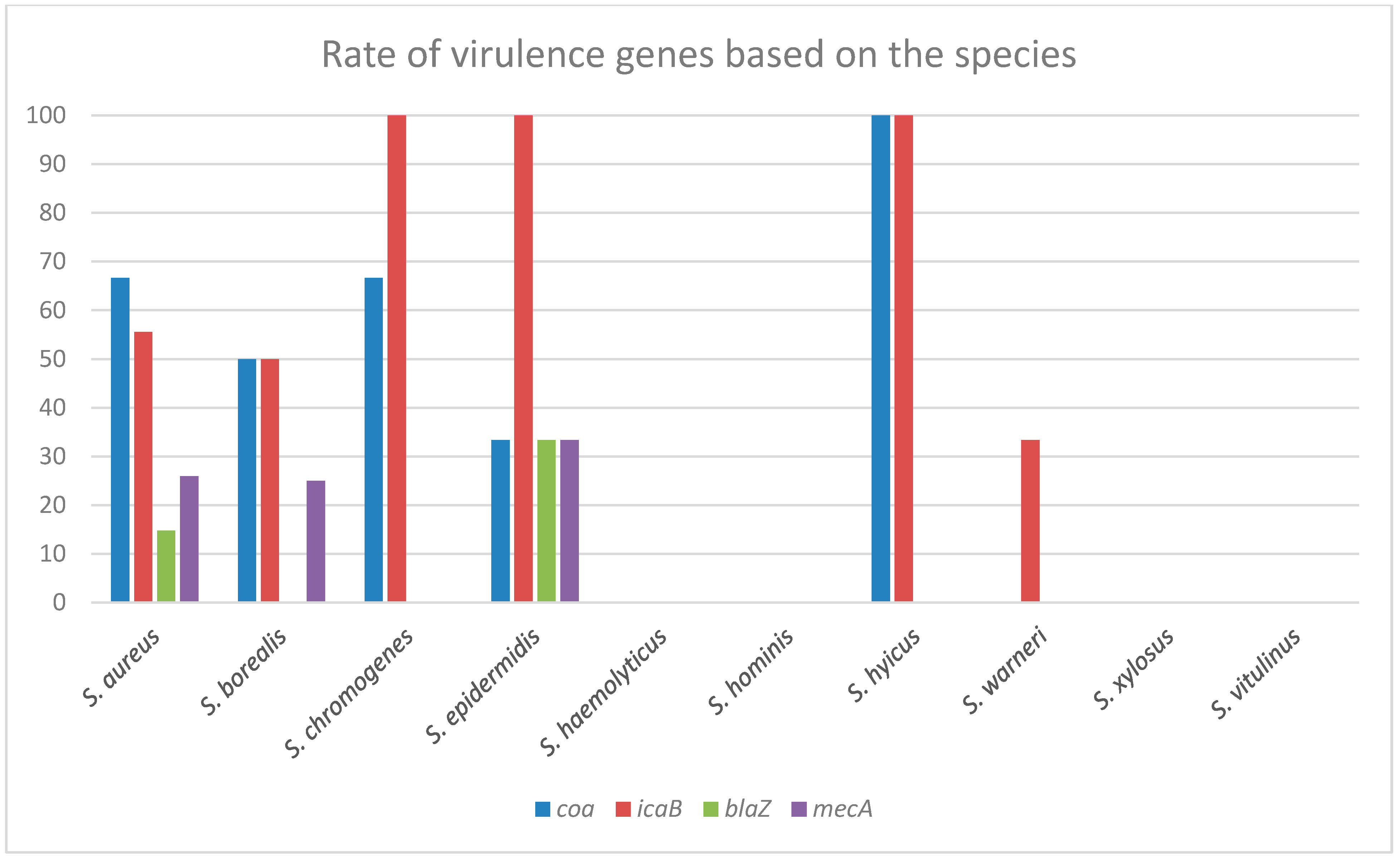
2.5. PCR Results
2.6. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation of Bacteria and MALDI-TOF MS Identificaiton
4.3. Molecular Characterization
4.4. Hemolysis Ability of Isolates
4.5. Congo Red Agar Assay
4.6. Biofilm Assay
4.7. Phenotypic Presence of Coagulase Enzyme
4.8. Multiple Antibiotic Resistance
4.9. Chromogenic Nitrocefin Disk Method
4.10. Determination of coa, blaZ, mecA, and icaB genes
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Rodríguez, F.; Mercanoglu Taban, B. A state-of-art review on multi-drug resistant pathogens in foods of animal origin: Risk factors and mitigation strategies. Front. Microbiol. 2019, 10, 2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loayza, F.; Graham, J.P.; Trueba, G. Factors obscuring the role of E. coli from domestic animals in the global antimicrobial resistance crisis: An evidence-based review. Int. J. Environ. Res. Public Health 2020, 17, 3061. [Google Scholar] [CrossRef]
- Cheng, W.N.; Han, S.G. Bovine mastitis: Risk factors, therapeutic strategies, and alternative treatments—A review. Asian-Australas. J. Anim. Sci. 2020, 33, 1699–1713. [Google Scholar] [CrossRef] [PubMed]
- Kabelitz, T.; Aubry, E.; van Vorst, K.; Amon, T.; Fulde, M. The role of Streptococcus spp. in bovine mastitis. Microorganisms 2021, 9, 1497. [Google Scholar] [CrossRef]
- Sezener, M.G.; Fındık, A.; Ergüden, V.E.; Akgöz, S.; Gülhan, T.; Çiftci, A. The determination of antibiotic resistances and some virulence genes of Staphylococcus aureus isolated from bovine mastitis. JAES 2019, 4, 182–187. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Yatoo, M.I.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef]
- Pekana, A.; Green, E. Antimicrobial resistance profiles of Staphylococcus aureus isolated from meat carcasses and bovine milk in abattoirs and dairy farms of the Eastern Cape, South Africa. Int. J. Environ. Res. Public Health 2018, 15, 2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costanzo, N.; Ceniti, C.; Santoro, A.; Clausi, M.T.; Casalinuovo, F. Foodborne pathogen assessment in raw milk cheeses. Int. J. Food Sci. 2020, 2020, 3616713. [Google Scholar] [CrossRef]
- González-Martín, M.; Corbera, J.A.; Suárez-Bonnet, A.; Tejedor-Junco, M.T. Virulence factors in coagulase-positive staphylococci of veterinary interest other than Staphylococcus aureus. Vet. Q. 2020, 40, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Skočková, A.; Bogdanovıčová, K.; Koláčková, I.; Karpíšková, R. Antimicrobial-resistant and extended-spectrum β-Lactamase–producing Escherichia coli in raw cow’s milk. J. Food Prot. 2015, 78, 72–77. [Google Scholar] [CrossRef]
- Ahmed, I.M. Detection of CTX-M gene in extended spectrum β-lactamases producing Enterobacteriaceae isolated from bovine milk. Iraqi J. Vet. Sci. 2021, 35, 397–402. [Google Scholar] [CrossRef]
- Read, A.F.; Woods, R.J. Antibiotic resistance management. Evol. Med. Public Health 2014, 2014, 147. [Google Scholar] [CrossRef]
- Andersson, D.I.; Balaban, N.Q.; Baquero, F. Antibiotic resistance: Turning evolutionary principles into clinical reality. FEMS Microbiol. Rev. 2020, 44, 171–188. [Google Scholar] [CrossRef]
- Sipahi, N.; Karakaya, E.; Ikiz, S. Phenotypic and genotypic investigation of the heavy metal resistance in Escherichia coli isolates recovered from cattle stool samples. Turk. J. Vet. Anim. Sci. 2019, 43, 684–691. [Google Scholar] [CrossRef]
- Melo, D.A.D.; Coelho, I.D.S.; Motta, C.C.D.; Rojas, A.C.C.M.; Dubenczuk, F.C.; Coelho, S.D.M.D.O.; Souza, M.M.S.D. Impairments of mecA gene detection in bovine Staphylococcus spp. Braz. J. Microbiol. 2014, 45, 1075–1082. [Google Scholar] [CrossRef] [Green Version]
- Gueimonde, M.; Sánchez, B.; G de los Reyes-Gavilán, C.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Sedky, D.; Ghazy, A.A.; Soliman, N.A.; Shaapan, R.M. Comparative diagnosis of infectious bacteria in bovine milk. J. Anim. Health Prod. 2020, 8, 171–182. [Google Scholar] [CrossRef]
- Taye, Y.; Degu, T.; Fesseha, H.; Mathewos, M. Isolation and identification of lactic acid bacteria from cow milk and milk products. Sci. World J. 2021, 2021, 4697445. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Hur, Y.K.; Kim, E.J.; Ahn, Y.T.; Kim, J.G.; Choi, Y.J.; Huh, C.S. Comparative analysis of the microbial communities in raw milk produced in different regions of Korea. Asian-Australas J. Anim. Sci. 2017, 30, 1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; In Lee, S.; Rackerby, B.; Frojen, R.; Goddik, L.; Ha, S.D.; Park, S.H. Assessment of overall microbial community shift during Cheddar cheese production from raw milk to aging. Appl. Microbiol. Biotechnol. 2020, 104, 6249–6260. [Google Scholar] [CrossRef] [PubMed]
- Kılınçel, Ö.; Çelik, O.E.; Aytan, A.; Alkan, İ.; Ayvaz, İ.B.; Öztürk, C.E. Düzce İlinde çiğ süt örneklerinde Mycobacterium bovis aranması. Düzce Üniversitesi Sağlık Bilim. Enstitüsü Derg. 2018, 8, 112–114. [Google Scholar]
- Wang, H.; Shen, J.; Zhu, C.; Ma, K.; Fang, M.; Li, B.; Wang, W.; Xue, T. Antibiotics Resistance and Virulence of Staphylococcus aureus Isolates Isolated from Raw Milk from Handmade Dairy Retail Stores in Hefei City, China. Foods 2022, 11, 2185. [Google Scholar] [CrossRef] [PubMed]
- Fischer, E.A.J.; Dierikx, C.M.; van Essen-Zandbergen, A.; Mevius, D.; Stegeman, A.; Velkers, F.C.; Klinkenberg, D. Competition between Escherichia coli Populations with and without Plasmids Carrying a Gene Encoding Extended-Spectrum Beta-Lactamase in the Broiler Chicken Gut. Appl. Environ. Microbiol. 2019, 85, e00892-19. [Google Scholar] [CrossRef] [Green Version]
- Rusenova, N.; Gebreyes, W.; Koleva, M.; Mitev, J.; Penev, T.; Vasilev, N.; Miteva, T. Comparison of three methods for routine detection of Staphylococcus aureus isolated from bovine mastitis. Kafkas Univ. Vet. Fak. Derg. 2013, 19, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Savaşan, S.; Kırkan, Ş.; Erbaş, G.; Parın, U.; Ciftci, A. Determination of virulence factors of staphylococci isolated from bovine mastitis. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 947–952. [Google Scholar] [CrossRef]
- Özenç, E.; Şeker, E.; Yılmaz, M. Abattoir-Based survey of mastitis in cattle in Afyonkarahisar Province. Kocatepe Vet. J. 2019, 12, 437–442. [Google Scholar] [CrossRef]
- Clark, C.M.; Costa, M.S.; Sanchez, L.M.; Murphy, B.T. Coupling MALDI-TOF mass spectrometry protein and specialized metabolite analyses to rapidly discriminate bacterial function. Proc. Natl. Acad. Sci. USA 2018, 115, 4981–4986. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Chui, H.; Domish, L.; Hernandez, D.; Wang, G. Recent development of mass spectrometry and proteomics applications in identification and typing of bacteria. Proteom-Clin. Appl. 2016, 10, 346–357. [Google Scholar] [CrossRef]
- Javed, M.U.; Ijaz, M.; Fatima, Z.; Anjum, A.A.; Aqib, A.I.; Ali, M.M.; Rehman, A.; Ahmed, A.; Ghaffar, A. Frequency and antimicrobial susceptibility of methicillin and vancomycin-resistant Staphylococcus aureus from bovine milk. Pak. Vet. J. 2021, 41, 463–468. [Google Scholar] [CrossRef]
- Javid, F.; Taku, A.; Bhat, M.A.; Badroo, G.A.; Mudasir, M.; Sofi, T.A. Molecular typing of Staphylococcus aureus based on coagulase gene. Vet. World 2018, 11, 423–430. [Google Scholar] [CrossRef]
- Effendi, M.H.; Hisyam, M.A.M.; Hastutiek, P.; Tyasningsih, W. Detection of coagulase gene in Staphylococcus aureus from several dairy farms in East Java, Indonesia, by polymerase chain reaction. Vet. World. 2019, 12, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Pashangeh, S.; Shekarforoush, S.S.; Aminlari, M.; Hosseinzadeh, S.; Nizet, V.; Dahesh, S.; Rahmdel, S. Inhibition of histamine accumulation by novel histamine-degrading species of Staphylococcus sp. isolated from goats and sheep milk. Food Sci. Nutr. 2022, 10, 354–362. [Google Scholar] [CrossRef]
- Otto, M. Coagulase-negative staphylococci as reservoirs of genes facilitating MRSA infection: Staphylococcal commensal species such as Staphylococcus epidermidis are being recognized as important sources of genes promoting MRSA colonization and virulence. Bioessays 2013, 35, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, B.; Rosalino, L.M.; Palmeira, J.D.; Torres, R.T.; Cunha, M.V. Antimicrobial resistance in commensal Staphylococcus aureus from wild ungulates is driven by agricultural land cover and livestock farming. Environ. Pollut. 2022, 303, 119116. [Google Scholar] [CrossRef] [PubMed]
- Zedan, A.; Alatfeehy, N.; Marouf, S. Isolation and antibiogram profiles of Staphylococcus aureus isolates from cow milk and dog samples. J. Appl. Vet. Sci. 2022, 8, 38–44. [Google Scholar] [CrossRef]
- Meroni, G.; Soares Filipe, J.F.; Drago, L.; Martino, P.A. Investigation on antibiotic-resistance, biofilm formation and virulence factors in multi drug resistant and non multi drug resistant Staphylococcus pseudintermedius. Microorganisms 2019, 7, 702. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, P.; Sachivkina, N.; Vatnikov, Y.; Shabunin, S.; Engashev, S.; Kontsevaya, S.; Karamyan, A.; Bokov, D.; Kuznetsova, O.; Vasilieva, E. Role of microorganisms isolated from cows with mastitis in Moscow region in biofilm formation. Vet. World 2021, 14, 40–48. [Google Scholar] [CrossRef]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus biofilm: Morphology, genetics, pathogenesis and treatment strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef]
- Azara, E.; Longheu, C.; Sanna, G.; Tola, S. Biofilm formation and virulence factor analysis of Staphylococcus aureus isolates collected from ovine mastitis. J. Appl. Microbiol. 2017, 123, 372–379. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, S.; Lou, X.; Cheng, S.; Liu, X.; Zheng, W.; Wang, H. Biofilm formation and prevalence of adhesion genes among Staphylococcus aureus isolates from different food sources. Microbiologyopen 2020, 9, e00946. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, L.V. Microbial biofilm in food processing. LWT-Food Sci. Technol. 1999, 32, 321–326. [Google Scholar] [CrossRef]
- Gün, İ.; Ekİncİ, F.Y. Biofilms: Microbial life on surfaces. J. Food 2009, 34, 165–173. [Google Scholar]
- Wu, Y.; Li, J.; Qiao, M.; Meng, D.; Meng, Q.; Qiao, J.; Cai, X. Characteristic profiles of biofilm, enterotoxins and virulence of Staphylococcus aureus isolates from dairy cows in Xinjiang Province, China. J. Vet. Med. 2019, 20, e74. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Shang, Y.; Wu, Y.; Wu, J.; Chen, J.; Wang, Z.; Yu, Z. Diclazuril inhibits biofilm formation and hemolysis of Staphylococcus aureus. ACS Infect. Dis. 2021, 7, 1690–1701. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.T.; da Silva, J.G.; Aragão, B.B.; Peixoto, R.M.; Mota, R.A. Occurrence of β-lactam-resistant Staphylococcus aureus in milk from primiparous dairy cows in the northeastern region of Brazil. Trop. Anim. Health Prod. 2020, 52, 2303–2307. [Google Scholar] [CrossRef] [PubMed]
- Cremonesi, P.; Severgnini, M.; Romanò, A.; Sala, L.; Luini, M.; Castiglioni, B. Bovine milk microbiota: Comparison among three different DNA extraction protocols to identify a better approach for bacterial analysis. Microbiol. Spectr. 2021, 9, e00374-21. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Meng, L.; Liu, H.; Wu, H.; Schroyen, M.; Zheng, N.; Wang, J. Effect of cephalosporin treatment on the microbiota and antibiotic resistance genes in feces of dairy cows with clinical mastitis. Antibiotics 2022, 11, 117. [Google Scholar] [CrossRef]
- Bidya, S.; Suman, R.S. Comparative study of three β lactamase test methods in Staphylococcus aureus isolated from two Nepalese hospitals. Open J. Clin. Diag. 2014, 4, 44037. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.M.; Martins, K.B.; Silva, V.R.D.; Mondelli, A.L.; Cunha, M.D.L.R.D.S.D. Correlation of phenotypic tests with the presence of the blaZ gene for detection of beta-lactamase. Braz. J. Microbiol. 2017, 48, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, A.G.; Csabai, I.; Krikó, E.; Tőzsér, D.; Maróti, G.; Patai, Á.V.; Solymosi, N. Antimicrobial resistance genes in raw milk for human consumption. Sci. Rep. 2020, 10, 7464. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.; Li, X.; Wang, X.; Li, H. Detection of antibiotic resistance, virulence gene, and drug resistance gene of Staphylococcus aureus ısolates from bovine mastitis. Microbiol. Spectr. 2022, 10, e00471-22. [Google Scholar] [CrossRef] [PubMed]
- El-Zamkan, M.A.; Mohamed, H.M. Antimicrobial resistance, virulence genes and biofilm formation in Enterococcus species isolated from milk of sheep and goat with subclinical mastitis. PloS ONE 2021, 16, e0259584. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, E.E.; Mousa, W.S.; Abdelsalam, S.Y.; Heikal, H.S.; Shawish, R.R.; Nooruzzaman, M.; Abdeen, A. Prevalence and characterization of coagulase positive Staphylococci from food products and human specimens in Egypt. Antibiotics 2021, 10, 75. [Google Scholar] [CrossRef]
- Haran, K.P.; Godden, S.M.; Boxrud, D.; Jawahir, S.; Bender, J.B.; Sreevatsan, S. Prevalence and characterization of Staphylococcus aureus, including methicillin-resistant Staphylococcus aureus, isolated from bulk tank milk from Minnesota dairy farms. J. Clin. Microbiol. 2012, 50, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Larsen, J.; Kjeldgaard, J.; Andersen, P.S.; Skov, R.; Ingmer, H. Methicillin-resistant and -susceptible Staphylococcus aureus from retail meat in Denmark. Int. J. Food Microbiol. 2017, 249, 72–76. [Google Scholar] [CrossRef]
- Delesalle, L.; Sadoine, M.L.; Mediouni, S.; Denis-Robichaud, J.; Zinszer, K.; Zarowsky, C.; Aenishaenslin, C.; Carabin, H. How are large-scale One Health initiatives targeting infectious diseases and antimicrobial resistance evaluated? A scoping review. One Health 2022, 14, 100380. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial antibiotic resistance: The most critical pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Tian, M.; He, X.; Feng, Y.; Wang, W.; Chen, H.; Gong, M.; Liu, D.; Clarke, J.L.; van Eerde, A. Pollution by Antibiotics and Antimicrobial Resistance in LiveStock and Poultry Manure in China, and Countermeasures. Antibiotics 2021, 10, 539. [Google Scholar] [CrossRef]
- Romandini, A.; Pani, A.; Schenardi, P.A.; Pattarino, G.A.C.; De Giacomo, C.; Scaglione, F. Antibiotic Resistance in Pediatric Infections: Global Emerging Threats, Predicting the Near Future. Antibiotics 2021, 10, 393. [Google Scholar] [CrossRef] [PubMed]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Brosius, J.; Palmer, M.L.; Kennedy, P.J.; Noller, H.F. Complete nucleotide sequence of a 16S ribosomal RNA gene from Escherichia coli. Proc. Natl. Acad. Sci. USA 1978, 75, 4801–4805. [Google Scholar] [CrossRef] [Green Version]
- Mariana, N.S.; Salman, S.A.; Neela, V.; Zamberi, S. Evaluation of modified Congo red agar for detection of biofilm produced by clinical isolates of methicillinresistance Staphylococcus aureus. Afr. J. Microbiol. Res. 2009, 3, 330–338. [Google Scholar]
- Zhang, H.; Xie, L.; Zhang, W.; Zhou, W.; Su, J.; Liu, J. The association of biofilm formation with antibiotic resistance in lactic acid bacteria from fermented foods. J. Food Saf. 2013, 33, 114–120. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Test for Bacteria Isolated from Animals. In Approved Standard Document, 5th ed.; CLSI supplement VET01S; CLSI: Malvern, PA, USA, 2020. [Google Scholar]
- De Jong, A.; Simjee, S.; El Garch, F. Antimicrobial susceptibility of enterococci recovered from healthy cattle, pigs and chickens in nine EU countries (EASSA Study) to critically important antibiotics. Vet. Microbiol. 2018, 216, 168–175. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Resistance (n = 84) | |||||
|---|---|---|---|---|---|
| Drug | R (n) | I (n) | Drug | R (n) | I (n) |
| TE | 3 | 6 | AMC | - | - |
| N | 9 | 6 | OX | 15 | - |
| K | 18 | 6 | FOX | 13 | - |
| CN | 3 | 15 | CIP | 3 | 3 |
| S | 15 | 30 | IMP | - | - |
| VA | 1 | - | E | 15 | 36 |
| F | - | 12 | P | 16 | - |
| SAM | - | - | |||
| Tür (n = 84) | Hemolysis | Weak Biofilm | Strong Biofilm | Coagulase | coa | icaB | mecA | blaZ |
|---|---|---|---|---|---|---|---|---|
| S. aureus (n = 27) | 6 | - | 15 | 18 | 18 | 15 | 7 | 4 |
| S. borealis (n = 12) | 6 | - | 6 | 6 | 6 | 6 | 3 | - |
| S. chromogenes (n = 9) | 6 | - | 9 | 6 | 6 | 9 | - | - |
| S. epidermidis (n = 9) | - | - | 9 | 3 | 3 | 9 | 3 | 3 |
| S. haemolyticus (n = 3) | - | 3 | - | - | - | - | - | - |
| S. hominis (n = 3) | - | - | - | - | - | - | - | - |
| S. hyicus (n = 6) | 3 | 6 | 6 | 6 | 6 | - | - | |
| S. warneri (n = 9) | 3 | 6 | 3 | - | - | 3 | - | - |
| S. xylosus (n = 3) | - | 3 | - | - | - | - | - | - |
| S. vitulinus (n = 3) | 3 | - | - | - | - | - | - | - |
| Correlations | |||||||
|---|---|---|---|---|---|---|---|
| Biofilm | Slime | icaB | coa | mecA | blaZ | ||
| Biofilm | Pearson Correlation | 1 | 1000 ** | 0.730 ** | 0.113 | 0.198 | 0.191 |
| p-Value | 0.000 | 0.000 | 0.305 | 0.071 | 0.082 | ||
| Slime | Pearson Correlation | 1000 ** | 1 | 0.730 ** | 0.113 | 0.198 | 0.191 |
| p-Value | 0.000 | 0.000 | 0.305 | 0.071 | 0.082 | ||
| icaB | Pearson Correlation | 0.730 ** | 0.730 ** | 1 | 0.372 ** | 0.304 ** | 0.261 * |
| p-Value | 0.000 | 0.000 | 0.000 | 0.005 | 0.016 | ||
| coa | Pearson Correlation | 0.113 | 0.113 | 0.372 ** | 1 | 0.196 | 0.151 |
| p-Value | 0.305 | 0.305 | 0.000 | 0.075 | 0.170 | ||
| mecA | Pearson Correlation | 0.198 | 0.198 | 0.304 ** | 0.196 | 1 | 0.586 ** |
| p-Value | 0.071 | 0.071 | 0.005 | 0.075 | 0.000 | ||
| blaZ | Pearson Correlation | 0.191 | 0.191 | 0.261 * | 0.151 | 0.586 ** | 1 |
| p-Value | 0.082 | 0.082 | 0.016 | 0.170 | 0.000 | ||
| Gene | 5′–3′ | Amplicon Size (bp) | References |
|---|---|---|---|
| coa | F-ACC ACA AGG TAC TGA ATC AAC G R-TGC TTT CGA TTG TTC GAT GC | 500–1000 | [55] |
| blaZ | F-CAAAGATGATATAGTTGCTTATTCTCC R-TGCTTGACCACTTTTATCAGC | 421 | [51] |
| mecA | F-AGA AGA TGG TAT GTG GAA GTT AG R-ATG TAT GTG CGA TTG TAT TGC | 583 | [55] |
| icaB | F-AGAATCGTGAAGTATAGAAAATT R-TCTAATCTTTTTCATGGAATCCGT | 880 | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sipahi, N.; Kaya, E.; Çelik, C.; Pınar, O. The Characterization and Beta-Lactam Resistance of Staphylococcal Community Recovered from Raw Bovine Milk. Antibiotics 2023, 12, 556. https://doi.org/10.3390/antibiotics12030556
Sipahi N, Kaya E, Çelik C, Pınar O. The Characterization and Beta-Lactam Resistance of Staphylococcal Community Recovered from Raw Bovine Milk. Antibiotics. 2023; 12(3):556. https://doi.org/10.3390/antibiotics12030556
Chicago/Turabian StyleSipahi, Nisa, Ertugrul Kaya, Cansu Çelik, and Orhan Pınar. 2023. "The Characterization and Beta-Lactam Resistance of Staphylococcal Community Recovered from Raw Bovine Milk" Antibiotics 12, no. 3: 556. https://doi.org/10.3390/antibiotics12030556
APA StyleSipahi, N., Kaya, E., Çelik, C., & Pınar, O. (2023). The Characterization and Beta-Lactam Resistance of Staphylococcal Community Recovered from Raw Bovine Milk. Antibiotics, 12(3), 556. https://doi.org/10.3390/antibiotics12030556