
Prevalence of the SigB-Deficient Phenotype among Clinical Staphylococcus aureus Isolates Linked to Bovine Mastitis


 , and
, and
Abstract
:1. Introduction
2. Results
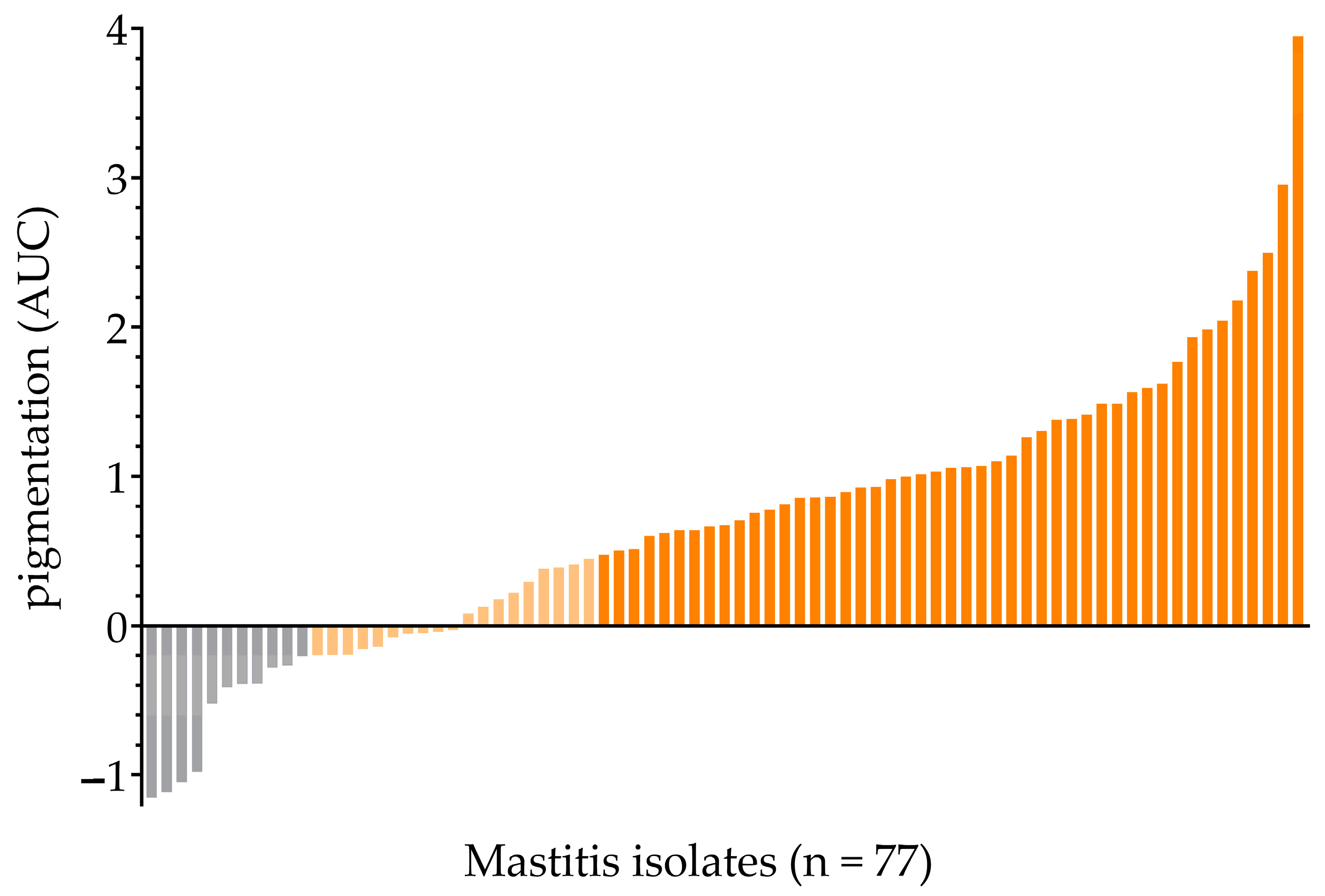
2.1. Screening for the SigB-Deficient Phenotype in Bovine Mastitis Isolates
2.1.1. Measurement of Extracted Pigments
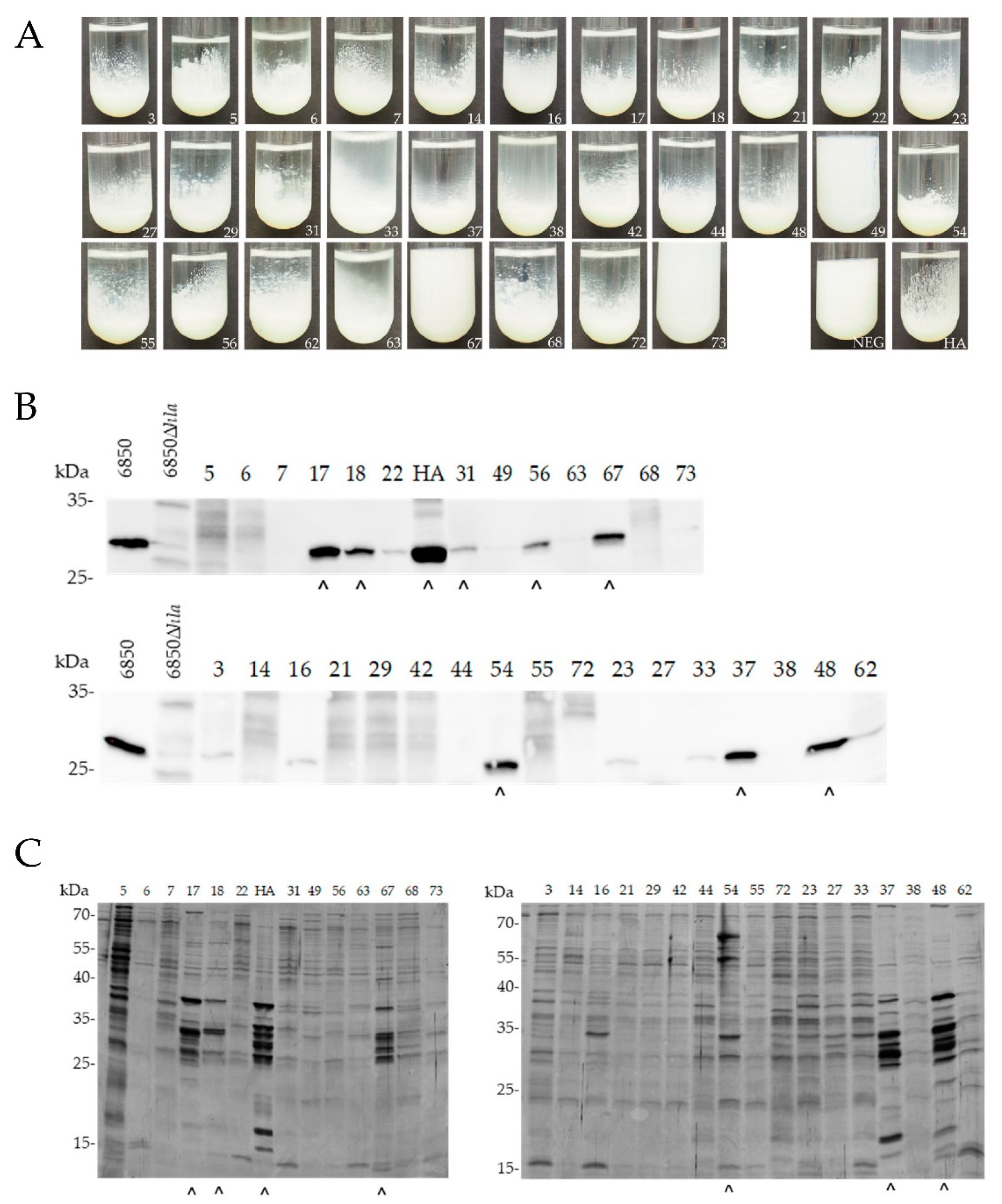
2.1.2. Proteolytic Activity, Immune-Based Detection of α-Hemolysin Secretion and Protein Secretion Profiling
2.1.3. Asp23-Transcriptional Expression (RT-qPCR)
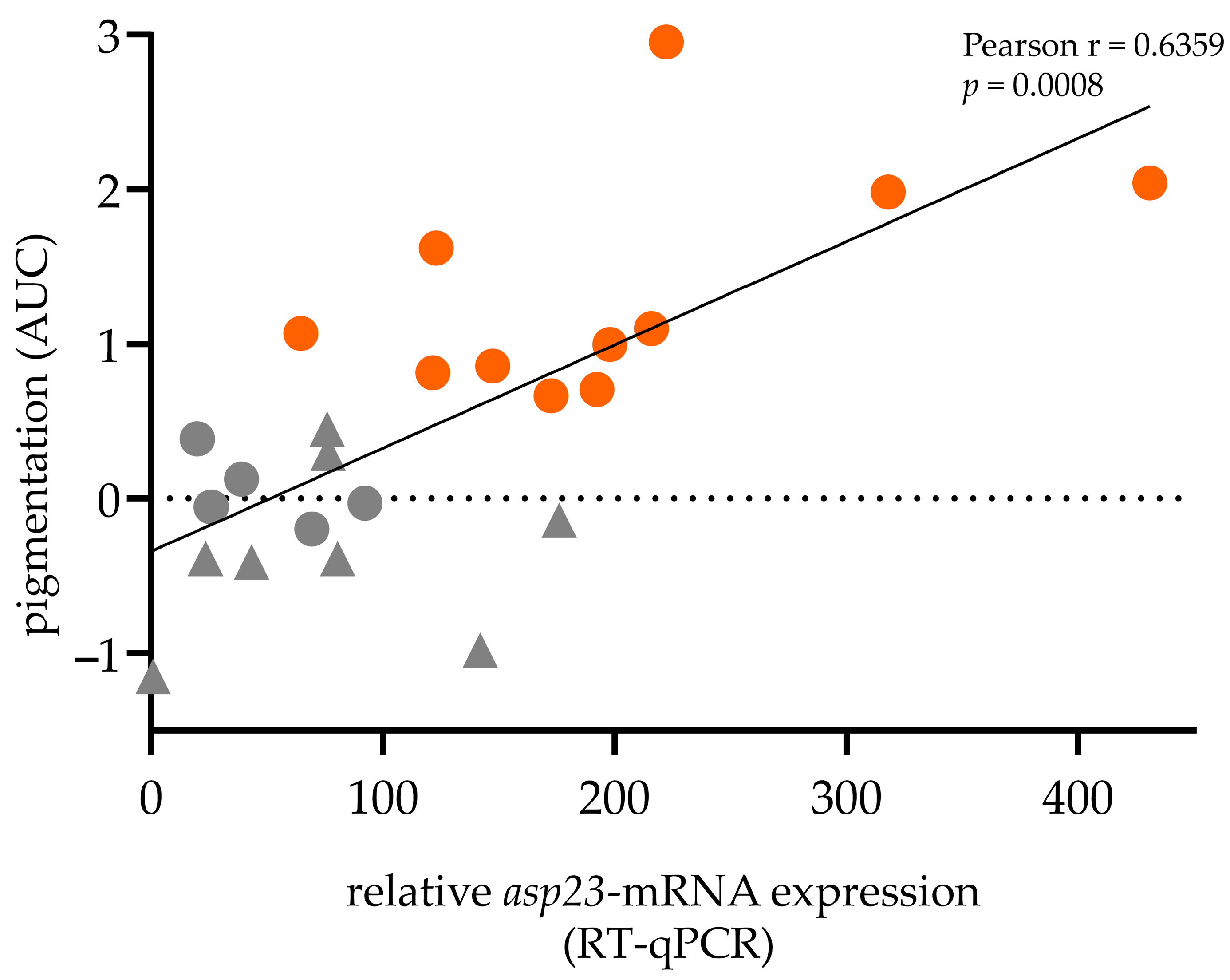
2.2. Positive Correlation between asp23-Transcriptional Expression and Pigmentation
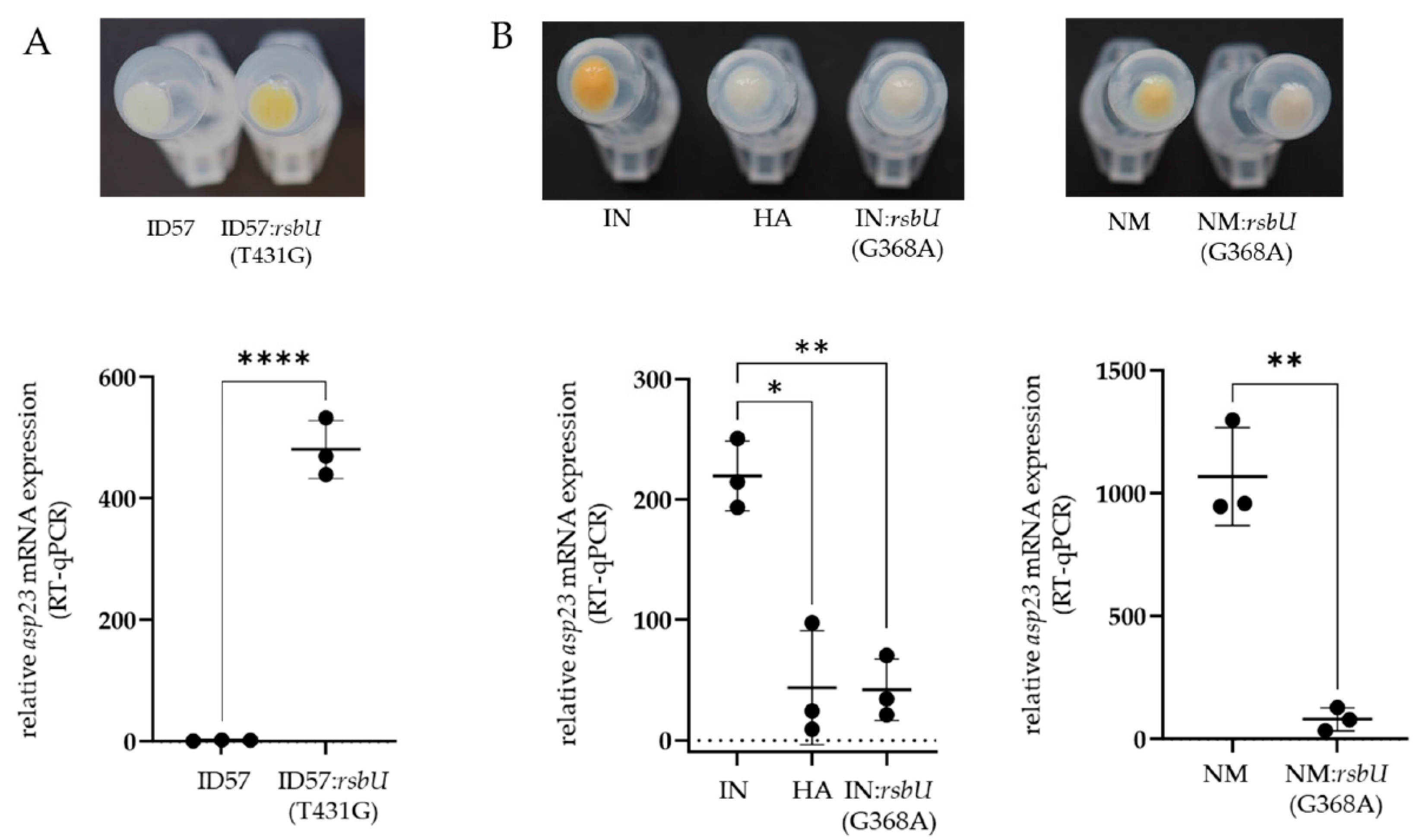
2.3. Sequence Analysis of the sigB Operon and Construction of Mutants to Confirm the SigB-Deficient Phenotype
3. Discussion
4. Materials and Methods
4.1. S. aureus Strains and Growth Conditions
4.2. Visualization and Determination of Carotenoid Pigmentation
4.3. Proteolytic Activity
4.4. α-Hemolysin (Hla) Western Blot and Analysis of Exoproteome Pattern
4.5. Reverse-Transcription (RT)-qPCR
4.6. Genetic Manipulation
4.7. Sequencing of the SigB-Locus and Molecular Strain Typing
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Löffler, B.; Proctor, R.A. Persistence of Staphylococcus aureus: Multiple Metabolic Pathways Impact the Expression of Virulence Factors in Small-Colony Variants (SCVs). Front. Microb. 2020, 11, 1028. [Google Scholar] [CrossRef] [PubMed]
- Atalla, H.; Gyles, C.; Mallard, B. Staphylococcus aureus small colony variants (SCVs) and their role in disease. Anim. Health Res. Rev. 2011, 12, 33–45. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pförtner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Herbert, S.; Ziebandt, A.K.; Ohlsen, K.; Schäfer, T.; Hecker, M.; Albrecht, D.; Novick, R.; Götz, F. Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect. Immun. 2010, 78, 2877–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson-Kanth, A.; Tegmark-Wisell, K.; Arvidson, S.; Oscarsson, J. Natural human isolates of Staphylococcus aureus selected for high production of proteases and alpha-hemolysin are sigmaB deficient. IJMM 2006, 296, 229–236. [Google Scholar] [CrossRef]
- McAdam, P.R.; Holmes, A.; Templeton, K.E.; Fitzgerald, J.R. Adaptive evolution of Staphylococcus aureus during chronic endobronchial infection of a cystic fibrosis patient. PLoS ONE 2011, 6, e24301. [Google Scholar] [CrossRef] [Green Version]
- Marbach, H.; Mayer, K.; Vogl, C.; Lee, J.Y.H.; Monk, I.R.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M.; Grunert, T. Within-host evolution of bovine Staphylococcus aureus selects for a SigB-deficient pathotype characterized by reduced virulence but enhanced proteolytic activity and biofilm formation. Sci. Rep. 2019, 9, 13479. [Google Scholar] [CrossRef] [Green Version]
- Giachino, P.; Engelmann, S.; Bischoff, M. Sigma(B) activity depends on RsbU in Staphylococcus aureus. J. Bacteriol. 2001, 183, 1843–1852. [Google Scholar] [CrossRef] [Green Version]
- Mayer, K.; Kucklick, M.; Marbach, H.; Ehling-Schulz, M.; Engelmann, S.; Grunert, T. Within-Host Adaptation of Staphylococcus aureus in a Bovine Mastitis Infection Is Associated with Increased Cytotoxicity. Int. J. Mol. Sci. 2021, 22, 8840. [Google Scholar] [CrossRef]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 6, GPP3-0031-2018. [Google Scholar] [CrossRef] [PubMed]
- Senn, M.M.; Giachino, P.; Homerova, D.; Steinhuber, A.; Strassner, J.; Kormanec, J.; Flückiger, U.; Berger-Bächi, B.; Bischoff, M. Molecular analysis and organization of the sigmaB operon in Staphylococcus aureus. J. Bacteriol. 2005, 187, 8006–8019. [Google Scholar] [CrossRef] [Green Version]
- Donegan, N.P.; Cheung, A.L. Regulation of the mazEF toxin-antitoxin module in Staphylococcus aureus and its impact on sigB expression. J. Bacteriol. 2009, 191, 2795–2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, P.D.; Endres, J.L.; Yajjala, V.K.; Widhelm, T.J.; Boissy, R.J.; Bose, J.L.; Bayles, K.W. A Genetic Resource for Rapid and Comprehensive Phenotype Screening of Nonessential Staphylococcus aureus Genes. mBio 2013, 4, e00537-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pané-Farré, J.; Jonas, B.; Förstner, K.; Engelmann, S.; Hecker, M. The σB regulon in Staphylococcus aureus and its regulation. IJMM 2006, 296, 237–258. [Google Scholar] [CrossRef]
- Pané-Farré, J.; Jonas, B.; Hardwick, S.W.; Gronau, K.; Lewis, R.J.; Hecker, M.; Engelmann, S. Role of RsbU in Controlling SigB Activity in Staphylococcus aureus following Alkaline Stress. J. Bacteriol. 2009, 191, 2561–2573. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bächi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus sigmaB regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef]
- Marshall, J.H.; Wilmoth, G.J. Proposed pathway of triterpenoid carotenoid biosynthesis in Staphylococcus aureus: Evidence from a study of mutants. J. Bacteriol. 1981, 147, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Pelz, A.; Wieland, K.P.; Putzbach, K.; Hentschel, P.; Albert, K.; Götz, F. Structure and biosynthesis of staphyloxanthin from Staphylococcus aureus. JBC 2005, 280, 32493–32498. [Google Scholar] [CrossRef] [Green Version]
- Wieland, B.; Feil, C.; Gloria-Maercker, E.; Thumm, G.; Lechner, M.; Bravo, J.M.; Poralla, K.; Götz, F. Genetic and biochemical analyses of the biosynthesis of the yellow carotenoid 4,4’-diaponeurosporene of Staphylococcus aureus. J. Bacteriol. 1994, 176, 7719–7726. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Geisinger, E. Quorum sensing in staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.L.; Chien, Y.T.; Bayer, A.S. Hyperproduction of alpha-hemolysin in a sigB mutant is associated with elevated SarA expression in Staphylococcus aureus. Infect. Immun. 1999, 67, 1331–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, M.; Entenza, J.M.; Giachino, P. Influence of a functional sigB operon on the global regulators sar and agr in Staphylococcus aureus. J. Bacteriol. 2001, 183, 5171–5179. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, A.; Saravia-Otten, P.; Tegmark, K.; Morfeldt, E.; Arvidson, S. Decreased Amounts of Cell Wall-Associated Protein A and Fibronectin-Binding Proteins in Staphylococcus aureus sarA Mutants due to Up-Regulation of Extracellular Proteases. Infect. Immun. 2001, 69, 4742–4748. [Google Scholar] [CrossRef] [Green Version]
- Gertz, S.; Engelmann, S.; Schmid, R.; Ohlsen, K.; Hacker, J.; Hecker, M. Regulation of sigmaB-dependent transcription of sigB and asp23 in two different Staphylococcus aureus strains. Mol. Gen. Genet. 1999, 261, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.H.; Wilmoth, G.J. Pigments of Staphylococcus aureus, a series of triterpenoid carotenoids. J. Bacteriol. 1981, 147, 900–913. [Google Scholar] [CrossRef] [Green Version]
- Kullik, I.; Giachino, P.; Fuchs, T. Deletion of the alternative sigma factor sigmaB in Staphylococcus aureus reveals its function as a global regulator of virulence genes. J. Bacteriol. 1998, 180, 4814–4820. [Google Scholar] [CrossRef] [Green Version]
- Kossakowska-Zwierucho, M.; Kaźmierkiewicz, R.; Bielawski, K.P.; Nakonieczna, J. Factors Determining Staphylococcus aureus Susceptibility to Photoantimicrobial Chemotherapy: RsbU Activity, Staphyloxanthin Level, and Membrane Fluidity. Front. Microb. 2016, 7, 1141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Suo, Y.; Zhang, D.; Jin, F.; Zhao, H.; Shi, C. Genetic and Virulent Difference Between Pigmented and Non-pigmented Staphylococcus aureus. Front. Microb. 2018, 9, 598. [Google Scholar] [CrossRef]
- Lan, L.; Cheng, A.; Dunman, P.M.; Missiakas, D.; He, C. Golden pigment production and virulence gene expression are affected by metabolisms in Staphylococcus aureus. J. Bacteriol. 2010, 192, 3068–3077. [Google Scholar] [CrossRef] [Green Version]
- Austin, C.M.; Garabaglu, S.; Krute, C.N.; Ridder, M.J.; Seawell, N.A.; Markiewicz, M.A.; Boyd, J.M.; Bose, J.L. Contribution of YjbIH to Virulence Factor Expression and Host Colonization in Staphylococcus aureus. Infect. Immun. 2019, 87, e00155-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, A.; Panthee, S.; Hamamoto, H.; Grunert, T.; Sekimizu, K. YjbH regulates virulence genes expression and oxidative stress resistance in Staphylococcus aureus. Virulence 2021, 12, 470–480. [Google Scholar] [CrossRef]
- Kwak, Y.K.; Högbom, M.; Colque-Navarro, P.; Möllby, R.; Vécsey-Semjén, B. Biological relevance of natural alpha-toxin fragments from Staphylococcus aureus. J. Membr. Biol. 2010, 233, 93–103. [Google Scholar] [CrossRef]
- Zielinska, A.K.; Beenken, K.E.; Mrak, L.N.; Spencer, H.J.; Post, G.R.; Skinner, R.A.; Tackett, A.J.; Horswill, A.R.; Smeltzer, M.S. sarA-mediated repression of protease production plays a key role in the pathogenesis of Staphylococcus aureus USA300 isolates. Mol. Microbiol. 2012, 86, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antók, F.I.; Mayrhofer, R.; Marbach, H.; Masengesho, J.C.; Keinprecht, H.; Nyirimbuga, V.; Fischer, O.; Lepuschitz, S.; Ruppitsch, W.; Ehling-Schulz, M.; et al. Characterization of Antibiotic and Biocide Resistance Genes and Virulence Factors of Staphylococcus Species Associated with Bovine Mastitis in Rwanda. Antibiotics 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunert, T.; Stessl, B.; Wolf, F.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Distinct phenotypic traits of Staphylococcus aureus are associated with persistent, contagious bovine intramammary infections. Sci. Rep. 2018, 8, 15968. [Google Scholar] [CrossRef] [Green Version]
- Grunert, T.; Wenning, M.; Barbagelata, M.S.; Fricker, M.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Rapid and reliable identification of Staphylococcus aureus capsular serotypes by means of artificial neural network-assisted Fourier transform infrared spectroscopy. J. Clin. Microbiol. 2013, 51, 2261–2266. [Google Scholar] [CrossRef] [Green Version]
- Kummel, J.; Stessl, B.; Gonano, M.; Walcher, G.; Bereuter, O.; Fricker, M.; Grunert, T.; Wagner, M.; Ehling-Schulz, M. Staphylococcus aureus Entrance into the Dairy Chain: Tracking S. aureus from Dairy Cow to Cheese. Front. Microb. 2016, 7, 1603. [Google Scholar] [CrossRef] [Green Version]
- Schabauer, A.; Pinior, B.; Gruber, C.M.; Firth, C.L.; Käsbohrer, A.; Wagner, M.; Rychli, K.; Obritzhauser, W. The relationship between clinical signs and microbiological species, spa type, and antimicrobial resistance in bovine mastitis cases in Austria. Vet. Microbiol. 2018, 227, 52–60. [Google Scholar] [CrossRef]
- Sordelli, D.O.; Buzzola, F.R.; Gomez, M.I.; Steele-Moore, L.; Berg, D.; Gentilini, E.; Catalano, M.; Reitz, A.J.; Tollersrud, T.; Denamiel, G.; et al. Capsule expression by bovine isolates of Staphylococcus aureus from Argentina: Genetic and epidemiologic analyses. J. Clin. Microbiol. 2000, 38, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Balwit, J.M.; van Langevelde, P.; Vann, J.M.; Proctor, R.A. Gentamicin-resistant menadione and hemin auxotrophic Staphylococcus aureus persist within cultured endothelial cells. J. Infect. Dis. 1994, 170, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, M.J.; Aish, J.L.; White, I.J.; Shaw, L.; Lithgow, J.K.; Foster, S.J. sigmaB modulates virulence determinant expression and stress resistance: Characterization of a functional rsbU strain derived from Staphylococcus aureus 8325-4. J. Bacteriol. 2002, 184, 5457–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novick, R. Properties of a cryptic high-frequency transducing phage in Staphylococcus aureus. Virology 1967, 33, 155–166. [Google Scholar] [CrossRef]
- Lâm, T.T.; Giese, B.; Chikkaballi, D.; Kühn, A.; Wolber, W.; Pané-Farré, J.; Schäfer, D.; Engelmann, S.; Fraunholz, M.; Sinha, B. Phagolysosomal integrity is generally maintained after Staphylococcus aureus invasion of nonprofessional phagocytes but is modulated by strain 6850. Infect. Immun. 2010, 78, 3392–3403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morikawa, K.; Maruyama, A.; Inose, Y.; Higashide, M.; Hayashi, H.; Ohta, T. Overexpression of Sigma Factor, ςB, Urges Staphylococcus aureus to Thicken the Cell Wall and to Resist β-Lactams. BBRC 2001, 288, 385–389. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Grunert, T.; Leitner, N.R.; Marchetti-Deschmann, M.; Miller, I.; Wallner, B.; Radwan, M.; Vogl, C.; Kolbe, T.; Kratky, D.; Gemeiner, M.; et al. A comparative proteome analysis links tyrosine kinase 2 (Tyk2) to the regulation of cellular glucose and lipid metabolism in response to poly(I:C). J. Proteom. 2011, 74, 2866–2880. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 2012, 3, e00277-11. [Google Scholar] [CrossRef] [Green Version]
- Monk, I.R.; Tree, J.J.; Howden, B.P.; Stinear, T.P.; Foster, T.J. Complete Bypass of Restriction Systems for Major Staphylococcus aureus Lineages. mBio 2015, 6, e00308-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, J.; Vergara-Irigaray, M.; Merino, N.; Penadés, J.R.; Lasa, I. sigmaB regulates IS256-mediated Staphylococcus aureus biofilm phenotypic variation. J. Bacteriol. 2007, 189, 2886–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, C.H.; Hacker, J.; Ziebuhr, W.; Lee, B.K.; Cho, S.H. Molecular characterization of regulatory genes associated with biofilm variation in a Staphylococcus aureus strain. J. Microbiol. Biotechnol. 2008, 18, 28–34. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Phenotypic Traits of SigB-deficiency | SigB-activity | sigB-operon | ST (CC) | |||
|---|---|---|---|---|---|---|---|
| ID | Pigmentation | Proteolysis | Hla | Protein | rel. asp23 | Mutations compared to | |
| (AUC) | secretion | secretion | expression | SigB-functional strains | |||
| 17 | −0.141 | + | + | + | 176.05 | no difference | ST504 (151) |
| 18 | −0.411 | + | + | + | 43.26 | syn. SNPs in mazeF/rsbW/sigB and one SNP in the non-coding region between rsbV and rsbU | ST5477 (3666) |
| 31 | −0.390 | + | + | 23.55 | SNP rsbU(G395A → S132N) # | ST8 (8) | |
| 37 | 0.291 | + | + | + | 76.30 | no difference | ST504 (151) |
| 48 | 0.447 | + | + | + | 76.02 | no difference | ST151 (151) |
| 54 | 0.386 | + | + | + | 19.89 | no difference | ST97 (97) |
| 56 | −1.150 | + | + | 0.99 | SNP rsbU(G431T → G144V) ## | ST9 (9) | |
| 67 | −0.980 | + | + | 142.00 | no difference | ST5476 (3591) | |
| Mastitis-derived SigB-deficient reference strain (Marbach et al., 2019) | |||||||
| HA | −1.10 | + | + | + | 4.83 | SNP rsbU(G368A → G122D) ## | ST352 (97) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walzl, A.; Marbach, H.; Belikova, D.; Vogl, C.; Ehling-Schulz, M.; Heilbronner, S.; Grunert, T. Prevalence of the SigB-Deficient Phenotype among Clinical Staphylococcus aureus Isolates Linked to Bovine Mastitis. Antibiotics 2023, 12, 699. https://doi.org/10.3390/antibiotics12040699
Walzl A, Marbach H, Belikova D, Vogl C, Ehling-Schulz M, Heilbronner S, Grunert T. Prevalence of the SigB-Deficient Phenotype among Clinical Staphylococcus aureus Isolates Linked to Bovine Mastitis. Antibiotics. 2023; 12(4):699. https://doi.org/10.3390/antibiotics12040699
Chicago/Turabian StyleWalzl, Anna, Helene Marbach, Darya Belikova, Claus Vogl, Monika Ehling-Schulz, Simon Heilbronner, and Tom Grunert. 2023. "Prevalence of the SigB-Deficient Phenotype among Clinical Staphylococcus aureus Isolates Linked to Bovine Mastitis" Antibiotics 12, no. 4: 699. https://doi.org/10.3390/antibiotics12040699
APA StyleWalzl, A., Marbach, H., Belikova, D., Vogl, C., Ehling-Schulz, M., Heilbronner, S., & Grunert, T. (2023). Prevalence of the SigB-Deficient Phenotype among Clinical Staphylococcus aureus Isolates Linked to Bovine Mastitis. Antibiotics, 12(4), 699. https://doi.org/10.3390/antibiotics12040699