
Membrane Phenotypic, Metabolic and Genotypic Adaptations of Streptococcus oralis Strains Destined to Rapidly Develop Stable, High-Level Daptomycin Resistance during Daptomycin Exposures

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. In Vitro DAP Passage
2.3. Minimum Inhibitory Concentrations (MICs)
2.4. Phenotypic Assays
2.4.1. Surface Charge
2.4.2. CM Fluidity
2.4.3. CM Phospholipid (PL) Composition
2.4.4. Lipidomics Analyses by Mass Spectrometry
2.4.5. Quantification of DAP Binding
2.5. Metabolic Assays
2.5.1. Cultivation Conditions
2.5.2. Measurement of Growth Rates
2.5.3. Determination of Glucose, Lactate, Ammonium, and Acetate Concentrations in Cultivation Media
2.5.4. Lactate Dehydrogenase (LDH) and Glycerol-3-Phosphate Dehydrogenase (GPDH) Enzymatic Assays
2.6. Genotypic Assay
2.6.1. Whole Genome Sequencing
2.6.2. Genome Assembly
2.6.3. Taxon Identification
2.6.4. Resistome
2.7. Statistical Analysis
3. Results
3.1. CM Phenotypic Characteristics of HLDR vs. Non-HLDR S. oralis Strains
3.1.1. Emergence of HLDR and Non-HLDR Variants by In Vitro DAP Passage
3.1.2. Cell Surface Charge
3.1.3. CM Fluidity
3.1.4. DAP Binding
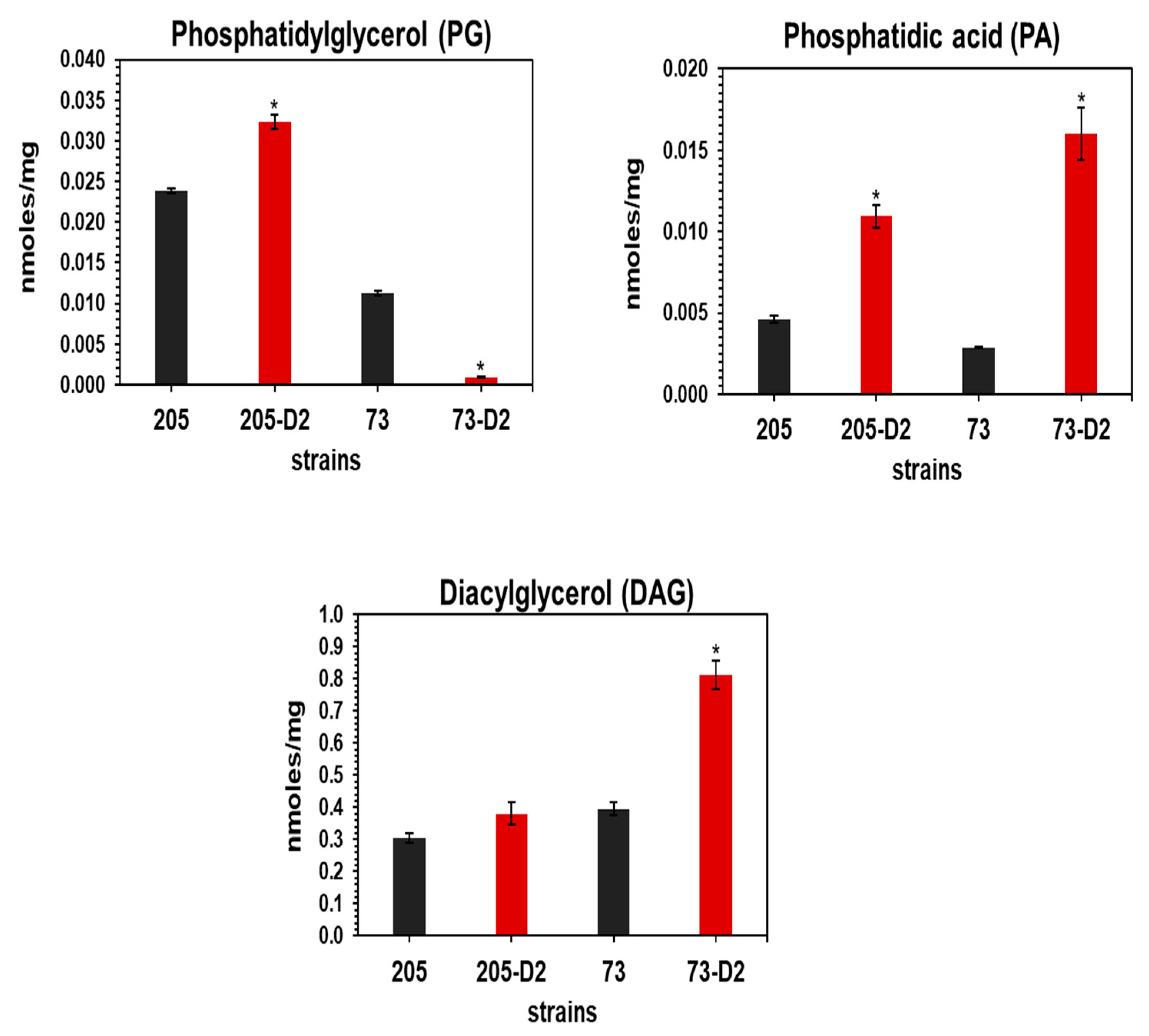
3.1.5. Quantitative Measurements of PL Contents by 2D-TLC and Lipidomics Analyses
4. Metabolic-Biochemical Profiles of HLDR Vs. Non-HLDR S. oralis Strains
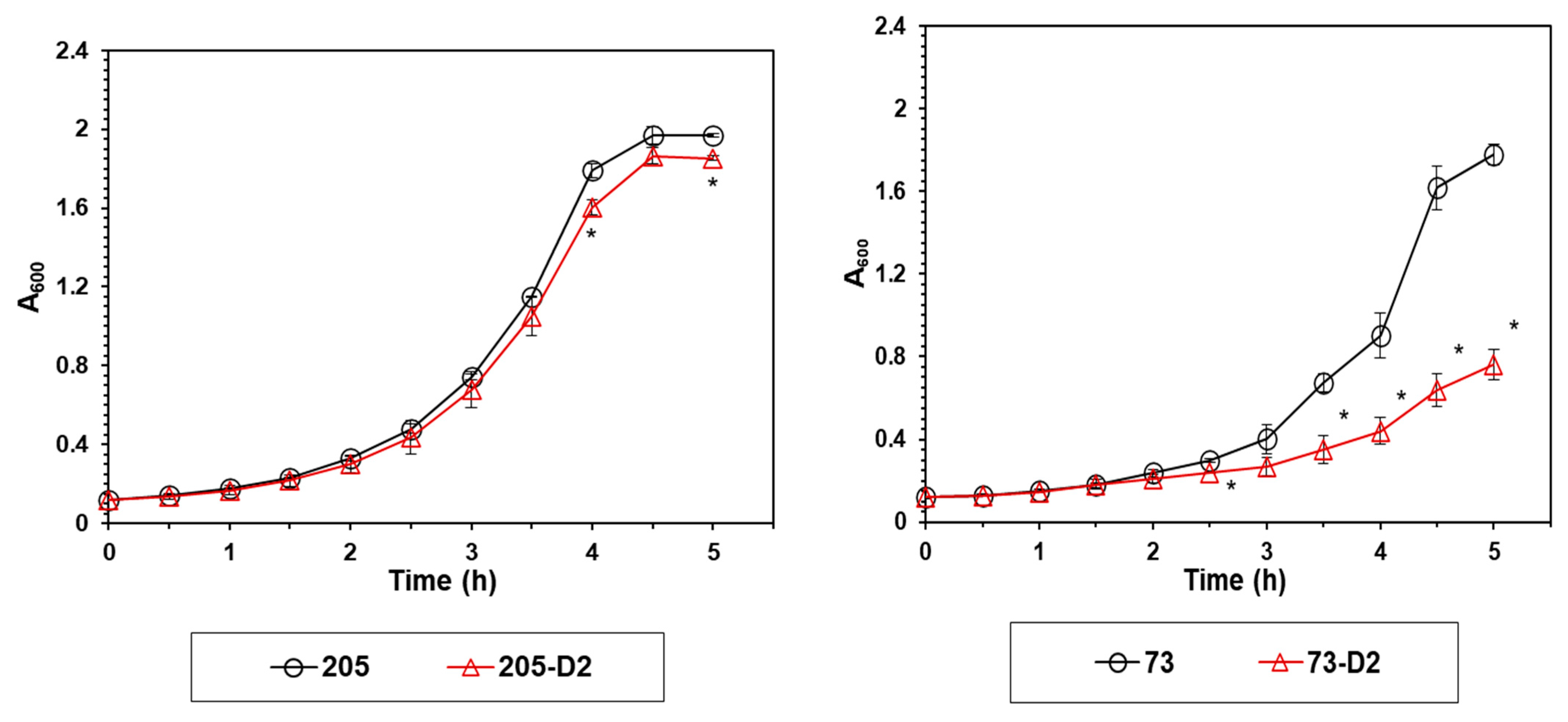
4.1. Growth Profiles
4.1.1. Glucose/Pyruvate Catabolism
4.1.2. Lactate Dehydrogenase (LDH) Activity
4.1.3. Glycerol-3-Phosphate Dehydrogenase (GPDH) Activity
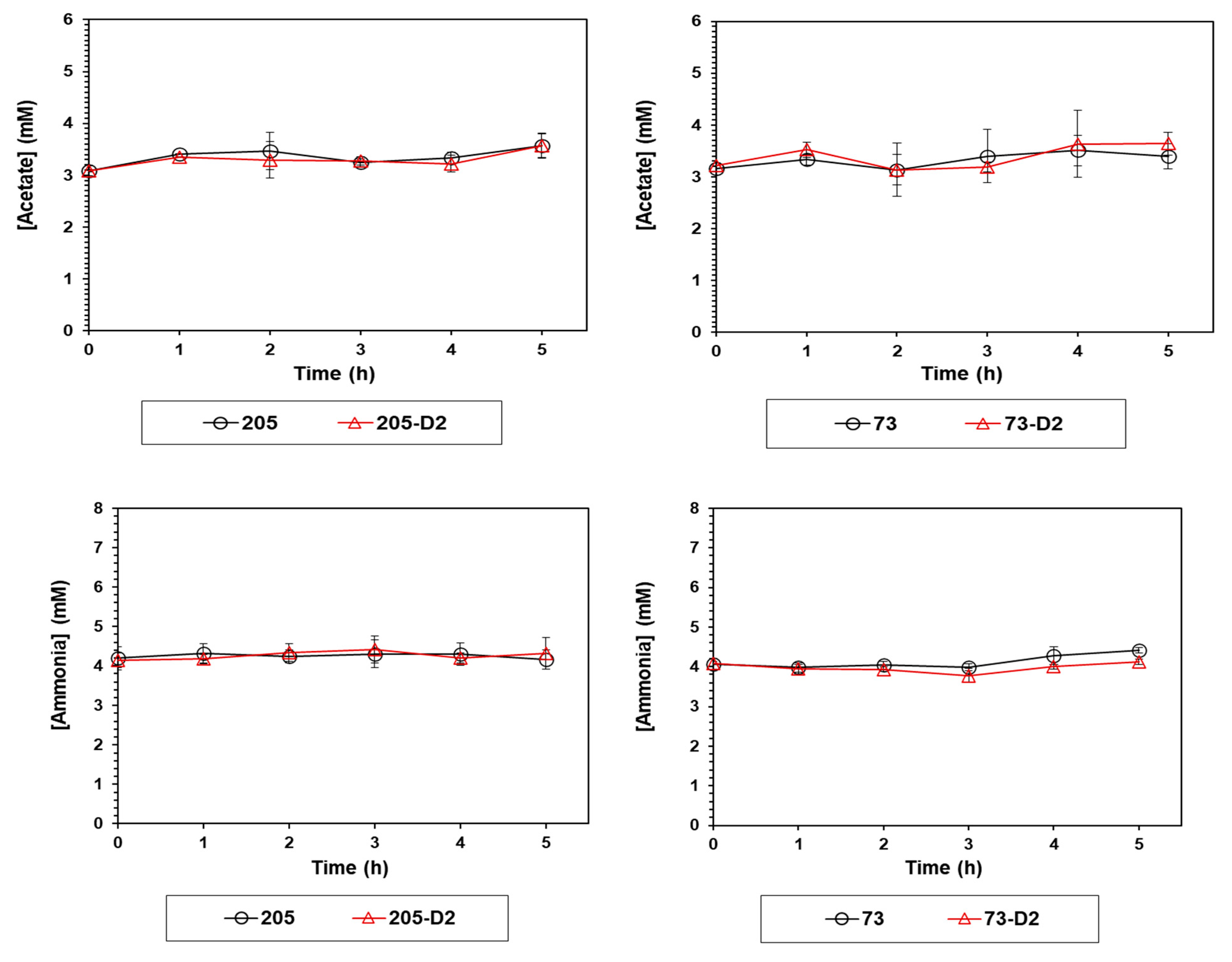
4.1.4. Measurement of Acetate and Ammonium in Culture
5. Genotypic Profiling by WGS
6. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-de-la-Maria, C.; Pericas, J.M.; Del Rio, A.; Castaneda, X.; Vila-Farres, X.; Arme, X.; Armeropinal, P.A.; Cervera, C.; Soy, D.; Falces, C.; et al. Hospital clinic experimental endocarditis dtudy group. Early in vitro and in vivo development of high-level daptomycin resistance is common in mitis group streptococci after exposure to daptomycin. Antimicrob. Agents Chemother. 2013, 57, 2319–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, T.L.; Bayer, A.S.; Fowler, V.G. Endocarditis and Intravascular Infections. In Principles and Practices of Infectious Diseases, 9th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Chapter 80. [Google Scholar]
- Marron, A.; Carratala, J.; Gonzalez-Barca, E.; Fernandez-Sevilla, A.; Alcaide, F.; Gudiol, F. Serious complications of bacteremia caused by viridans streptococci in neutropenic patients with cancer. Clin. Infect. Dis. 2000, 31, 1126–1130. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.T.; Chang, L.Y.; Hsueh, P.R.; Lu, C.Y.; Shao, P.L.; Huang, F.Y.; Lee, P.I.; Chen, C.M.; Lee, C.Y.; Huang, L.M. Clinical features and complications of viridans streptococci bloodstream infection in pediatric hemato-oncology patients. J. Microbiol. Immunol. Infect. 2007, 40, 349–354. [Google Scholar] [PubMed]
- Shelburne, S.A.; Sahasrabhojane, P.; Saldana, M.; Hui, Y.; Xiaoping, S.; Horstmann, N.; Thompson, E.; Flores, A.R. Streptococcus mitis strains causing severe clinical disease in cancer patients. Emerg. Infect. Dis. 2014, 20, 762–771. [Google Scholar] [CrossRef]
- Safdar, A.; Rolston, K.V. Vancomycin tolerance, a potential mechanism for refractory gram-positive bacteremia observational study in patients with cancer. Cancer 2006, 106, 1815–1820. [Google Scholar] [CrossRef]
- Akins, R.L.; Katz, B.D.; Monahan, C.; Alexander, D. Characterization of high-level daptomycin resistance in viridans group streptococci developed upon in vitro exposure to daptomycin. Antimicrob. Agents Chemother. 2015, 59, 2102–2112. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.N.; Tran, T.T.; Seepersaud, R.; Garcia-de-la-Maria, C.; Faull, K.; Yoon, A.; Proctor, R.A.; Miro, J.M.; Rybak, M.J.; Bayer, A.S.; et al. Perturbations of phosphatidate cytidylyltransferase (CdsA) mediate daptomycin resistance in Streptococcus mitis/oralis by a novel mechanism. Antimicrob. Agents Chemother. 2017, 61, e02435-16. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.N.; Tran, T.T.; Arias, C.A.; Seepersaud, R.; Sullam, P.M.; Bayer, A.S. Strain-specific adaptations of Streptococcus mitis-oralis to serial in vitro passage in daptomycin (DAP): Genotypic and phenotypic characteristics. Antibiotics 2020, 15, 520. [Google Scholar] [CrossRef]
- Tran, T.T.; Mishra, N.N.; Seepersaud, R.; Diaz, L.; Rios, R.; Dinh, A.Q.; Garcia-de-la-Maria, C.; Rybak, M.J.; Miro, J.M.; Shelburne, S.A.; et al. Mutations in cdsA and pgscorrelate with daptomycin resistance in Streptococcus mitis and S. oralis. Antimicrob. Agents Chemother. 2019, 63, e01531-18. [Google Scholar]
- Parrett, A.; Reed, J.M.; Gardner, S.G.; Mishra, N.N.; Bayer, A.S.; Powers, R.; Somerville, G.A. Metabolic changes associated with adaptive resistance to daptomycin in Streptococcus mitis-oralis. BMC Microbiol. 2020, 20, 162. [Google Scholar] [CrossRef]
- Lee, J.H.; Ji, H.J.; Seo, H.S.; Sullam, P.M. Complete genome sequence of Streptococcus oralis SF100, isolated from blood cultures from a patient with infective endocarditis. Microbiol. Resource Announc. 2021, 10, e0017621. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; McIver, L.J.; Blanco-Míguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M.; et al. Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery. eLife 2021, 10, e65088. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, W.Y.; Williams, K.J.; Su, B.; Bensinger, S.J. Profiling of mouse macrophage lipidome using direct infusion shotgun mass spectrometry. STAR Protoc. 2020, 2, 100235. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Bettcher, L.F.; Hsieh, W.Y.; Hornburg, D.; Pearson, M.J.; Blomberg, N.; Giera, M.; Snyder, M.P.; Raftery, D.; Bensinger, S.J.; et al. A DMS shotgun lipidomics workflow application to facilitate high-throughput, comprehensive lipidomics. J. Am. Soc. Mass Spectrom. 2021, 32, 2655–2663. [Google Scholar] [CrossRef]
- Mishra, N.N.; Bayer, A.S.; Baines, S.L.; Hayes, A.S.; Howden, B.P.; Lapitan, C.K.; Lew, C.; Rose, W.E. Cell membrane adaptations mediate β-lactam-induced resensitization of daptomycin-resistant (DAP-R) Staphylococcus aureus in vitro. Microorganisms 2021, 9, 1028. [Google Scholar] [CrossRef]
- Kebriaei, R.; Bayer, A.S.; Lapitan, C.K.; Rybak, M.J.; Somerville, G.A.; Mishra, N.N. Activity of the lactate dehydrogenase inhibitor oxamic acid against the fermentative bacterium Streptococcus mitis/oralis: Bactericidal effects and prevention of daptomycin resistance in vitro and in an ex vivo model. Antibiotics 2022, 11, 1409. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Hu, J.; Fan, J.; Sun, Z.; Liu, S. Next Polish: A fast and efficient genome polishing tool for long-read assembly. Bioinformatics 2020, 36, 2253–2255. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.N.; Bayer, A.S.; Tran, T.T.; Shamoo, Y.; Mileykovskaya, E.; Dowhan, W.; Guan, Z.; Arias, C.A. Daptomycin resistance in enterococci is associated with distinct alterations of cell membrane phospholipid content. PLoS ONE 2012, 7, e43958. [Google Scholar] [CrossRef]
- Kebriaei, R.; Rice, S.A.; Stamper, K.C.; Seepersaud, R.; Garcia-de-la-Maria, C.; Mishra, N.N.; Miro, J.M.; Arias, C.A.; Tran, T.T.; Sullam, P.M.; et al. Daptomycin dose-ranging evaluation with single-dose versus multidose ceftriaxone combinations against Streptococcus mitis/oralis in an ex vivo simulated endocarditis vegetation model. Antimicrob. Agents Chemother. 2019, 263, e00386-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaupp, R.; Lei, S.; Reed, J.M.; Peisker, H.; Boyle-Vavra, S.; Bayer, A.S.; Bischoff, M.; Herrmann, M.; Daum, R.S.; Powers, R.; et al. Staphylococcus aureus metabolic adaptations during the transition from a daptomycin susceptible phenotype to a daptomycin non-susceptible phenotype. Antimicrob. Agents Chemother. 2015, 59, 4226–4238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, L.; Andersson, C.; Sund, M.L.; Shockman, G.D. Protoplast formation and localization of enzymes in Streptococcus mitis. Infect. Immun. 1983, 40, 1146–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinhal, S.; Ropers, D.; Geiselmann, J.; de Jong, H. Acetate metabolism and the inhibition of bacterial growth by acetate. J. Bacteriol. 2019, 201, e00147-19. [Google Scholar] [CrossRef] [Green Version]
- Hagiya, H.; Sugawara, Y.; Kimura, K.; Hamaguchi, S.; Nishi, I.; Hayashi, M.; Akeda, Y.; Tomono, K. Emergence of daptomycin non-susceptible coagulase-negative staphylococci in patients with cardiovascular device infections: Two cases report investigated by whole genome analysis. Medicine 2018, 97, e13487. [Google Scholar] [CrossRef]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 2137–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, D.R.; Mortin, L.I.; Rubio, A.; Mylonakis, E.; Moellering, R.C.; Eliopoulos, G.M., Jr.; Peleg, A.Y. Impact of daptomycin resistance on Staphylococcus aureus virulence. Virulence 2015, 6, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Higgs, C.; Turner, A.M.; Nong, Y.; Gorrie, C.L.; Sherry, N.L.; Dyet, K.H.; Seemann, T.; Williamson, D.A.; Stinear, T.P.; et al. Daptomycin resistance occurs predominantly in vanA-type vancomycin-resistant Enterococcus faecium in Australa and is associated with heterogeneous and novel mutations. Front. Microbiol. 2021, 12, 749935. [Google Scholar] [CrossRef]
- Mollerup, S.; Elmeskov, C.; Pinholt, M.; Sejersen, T.S.; Pedersen, M.S.; Worning, P.; Frees, D.; West, H. Rapid in vivo development of resistance to daptomycin in vancomycin-resistant Enterococcus faecium due to genomic alterations. FEMS Microbiol. Lett. 2022, 369, fnac063. [Google Scholar] [CrossRef]
- Adams, H.M.; Joyce, L.R.; Guan, Z.; Akins, R.L.; Palmer, K.L. Streptococcus mitis and S. oralis lack a requirement for CdsA, the enzyme required for synthesis of major membrane phospholipids in bacteria. Antimicrob. Agents Chemother. 2017, 61, e02552-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeaman, M.R.; Chan, L.C.; Mishra, N.N.; Bayer, A.S. Mechanistic fingerprinting reveals kinetic signatures of resistance to daptomycin and host defense peptides in Streptococcus mitis-oralis. Antibiotics 2021, 10, 404. [Google Scholar] [CrossRef]
- Muraih, J.K.; Harris, J.; Taylor, S.D.; Palmer, M. Characterization of daptomycin oligomerization with perylene excimer fluorescence: Stoichiometric binding of phosphatidylglycerol triggers oligomer formation. Biochim. Biophys. Acta. 2012, 1818, 673–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraih, J.K.; Pearson, A.; Silverman, J.; Palmer, M. Oligomerization of daptomycin on membranes. Biochim. Biophys. Acta. 2011, 1808, 1154–1160. [Google Scholar] [CrossRef] [Green Version]
- Carman, G.M.; Han, G.S. Phosphatidic acid phosphatase, a key enzyme in the regulation of lipid synthesis. J. Biol. Chem. 2009, 284, 2593–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2006, 40, 133–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, L.R.; Guan, Z.; Palmer, K.L. Phosphatidylcholine biosynthesis in mitis group streptococci via host metabolite scavenging. J. Bacteriol. 2019, 201, e00495-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, T.C.; Kozjak-Pavlovic, V. Diverse facets of sphingolipid involvement in bacterial infections. Front. Cell. Dev. Biol. 2019, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- McMurchie, E.J.; Raison, J.K. Membrane lipid fluidity and its effect on the activation energy of membrane-associated enzymes. Biochim. Biophys. Acta 1979, 554, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Carman, G.M.; Han, G.S. Roles of phosphatidate phosphatase enzymes in lipid metabolism. Trends Biochem. Sci. 2006, 31, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.Y.; Kennedy, E.P. Phosphatidyl glycerophosphate phosphatase. J. Lipid Res. 1967, 8, 456–462. [Google Scholar] [CrossRef]
- Yeh, J.I.; Chinte, U.; Du, S. Structure of glycerol-3-phosphate dehydrogenase, an essential monotopic membrane enzyme involved in respiration and metabolism. Proc. Natl. Acad. Sci. USA 2008, 105, 3280–3285. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.A.; Panesso, D.; McGrath, D.M.; Qin, X.; Mojica, M.F.; Miller, C.; Diaz, L.; Tran, T.T.; Rincon, S.; Barbu, E.M.; et al. Genetic basis for in vivo daptomycin resistance in enterococci. N. Engl. J. Med. 2011, 365, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.; Panesso, D.; Gao, H.; Roh, J.H.; Munita, J.M.; Reyes, J.; Diaz, L.; Lobos, E.A.; Shamoo, Y.; Mishra, N.N.; et al. Whole-genome analysis of a daptomycin-susceptible Enterococcus faecium strain and its daptomycin-resistant variant arising during therapy. Antimicrob. Agents Chemother. 2013, 57, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Hines, K.M.; Waalkes, A.; Penewit, K.; Holmes, E.A.; Salipante, S.J.; Werth, B.J.; Xu, L. Characterization of the mechanisms of daptomycin resistance among gram-positive bacterial pathogens by multidimensional lipidomics. mSphere 2017, 2, e00492-17. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.; Panesso, D.; Mishra, N.N.; Mileykovskaya, E.; Guan, Z.; Munita, J.M.; Reyes, J.; Diaz, L.; Weinstock, G.M.; Murray, B.E.; et al. Daptomycin-resistant Enterococcus faecalis diverts the antibiotic molecule from the division septum and remodels cell membrane phospholipids. mBio 2013, 4, e00281-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.N.; Abdelhady, W.; Elsayed, A.M.; Lapitan, C.; Proctor, R.A.; Rybak, M.J.; Miro, J.M.; Bayer, A.S. Combinations of daptomycin plus ceftriaxone, but not ascending daptomycin dose-regimens, are effective in experimental endocarditis caused by Streptococcus mitis-oralis strains: Target tissue clearances and prevention of emergence of daptomycin-resistance. Antimicrob. Agents Chemother. 2023, 67, e0147222. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. oralis Strains | DAP MIC (µg/mL) | DAP MIC (Passaged in DAP Free Media for 5 Days) | |
|---|---|---|---|
| 73 | 0.5 | 0.5 | |
| HLDR | 73-D2 | >256 * | >256 |
| 205 | 0.5 | 0.5 | |
| Non-HLDR | 205-D2 | 8 ** | 2 |
| S. oralis Strains | CM Fluidity: PI Value (Mean ± SD) | Surface Charge: % of Cyt C in Supernatant (Mean ± SD) | BODIPY-DAP Binding: Fluorescence Intensity (Mean ± SD) |
|---|---|---|---|
| 73 | 0.298 ± 0.01 | 76 ± 3 | 553 ± 28 |
| 73-D2 | 0.271 ± 0.01 * | 61 ± 1 * | 592 ± 17 * |
| 205 | 0.268 ± 0.01 | 62 ± 8 | 405 ± 3 |
| 205-D2 | 0.269 ± 0.01 | 58 ± 7 | 430 ± 24 |
| % of PL Species (Mean ± SD) | |||
|---|---|---|---|
| Strains | PG | CL | PA |
| 73 | 18 ± 8 | 52 ± 4 | 31 ± 10 |
| 73-D2 | 0 ± 0 | 0 ± 0 | 100 ± 0 * |
| 205 | 29 ± 22 | 67 ± 19 | 4 ± 4 |
| 205-D2 | 18 ± 7 | 68 ± 6 | 15 ± 5 * |
| Predicted Amino Acid Change a | Predicted Protein Function | Gene ID |
|---|---|---|
| Val876Ile | LPXTG-anchored beta-N-acetylhexosaminidase StrH | gene-EL140_RS00270 |
| Lys3Glu | PspC domain-containing protein | gene-EL140_RS00510 |
| Lys175Gly | DUF1307 domain-containing protein | gene-EL140_RS02430 |
| Lys144Gln | response regulator transcription factor VncR | gene-EL140_RS02580 |
| Thr131Ser, Lys135Val | rRNA pseudouridine synthase | gene-EL140_RS01050 |
| Lys320Asn | glucosaminidase domain-containing protein | gene-EL140_RS03645 |
| 2_5delATGGTinsTGGCa, Lys6Gln | tagatose-bisphosphate aldolase | gene-EL140_RS04205 |
| Arg518Ser, Val527Leu | glycoside hydrolase family 13 protein | gene-EL140_RS04725 |
| Ile131Val | ABC transporter permease | gene-EL140_RS00390 |
| Ile27Met | hypothetical protein | gene-EL140_RS00190 |
| Thr28Asn, Ile194Val | L-lactate dehydrogenase | gene-EL140_RS05540 |
| Ser134Gly | hypothetical protein | gene-EL140_RS00190 |
| 66delAa | Fe-S cluster assembly protein SufB | gene-EL140_RS05570 |
| Lys1240Asp | Cna B-type domain-containing protein | gene-EL140_RS05785 |
| Gly71fs | YdbC family protein | gene-EL140_RS05795 |
| Ser522Leu | ABC transporter ATP-binding protein/permease | gene-EL140_RS00650 |
| Gln569Glu | aminodeoxychorismate synthase component I | gene-EL140_RS06465 |
| Leu60Pro | response regulator | gene-EL140_RS06480 |
| Ser242Thr | ammonia-dependent NAD(+) synthetase | gene-EL140_RS06505 |
| Leu417Gln | YSIRK-type signal peptide-containing protein | gene-EL140_RS06920 |
| Val144Ala | MarR family transcriptional regulator | gene-EL140_RS01740 |
| 2652delGACATinsAACACa | valine—tRNA ligase | gene-EL140_RS07795 |
| Ile1380Thr | YSIRK-type signal peptide-containing protein | gene-EL140_RS06920 |
| GlnAla68GluLeu | phosphoribulokinase | gene-EL140_RS08745 |
| Cys610Gly | magnesium-translocating P-type ATPase | gene-EL140_RS08825 |
| Cys104Ser | GH92 family glycosyl hydrolase | gene-EL140_RS09195 |
| TyrLys427PheGlu | cell surface protein | gene-EL140_RS09325 |
| Ala900Gly, Ala918Val, Ala912Val, Phe461Leu, Asp456Asn | LPXTG cell wall anchor domain-containing protein | gene-EL140_RS06375 |
| Asn82Ser | DUF6261 family protein | gene-EL140_RS09590 |
| Lys292Asn, Ala298Gln | LPXTG cell wall anchor domain-containing protein | gene-EL140_RS06375 |
| Ile99Val, Ala249Ser | methyltransferase domain-containing protein | gene-EL140_RS00590 |
| Lys192Asn | hemolysin family protein | gene-EL140_RS01300 |
| Thr68Met | endonuclease MutS2 | gene-EL140_RS01700 |
| Met44Val | hypothetical protein | gene-EL140_RS00190 |
| Ile166Ser | N-acetylmuramoyl-L-alanine amidase | gene-EL140_RS02630 |
| Arg68Gly, Glu72Asp | GDSL-type esterase/lipase family protein | gene-EL140_RS03180 |
| His4Tyr | Nramp family divalent metal transporter | gene-EL140_RS03335 |
| ArgLysGly154LysAlaGlu, Glu308fs, Ala2Val, Gln6Asn | tagatose-6-phosphate kinase | gene-EL140_RS04200 |
| Asn240Asp | pneumococcal-type histidine triad protein | gene-EL140_RS04260 |
| Asp61Tyr, Val70Ala | HNH endonuclease | gene-EL140_RS04615 |
| Leu229Ile | hypothetical protein | gene-EL140_RS00190 |
| Phe125Leu, Thr105Pro | hypothetical protein | gene-EL140_RS00190 |
| Ile110Met | nicotinate phosphoribosyltransferase | gene-EL140_RS06510 |
| Ser755Thr, Gln681Glu | G5 domain-containing protein | gene-EL140_RS06930 |
| Asn624Glu | penicillin-binding protein PBP2B | gene-EL140_RS07010 |
| Asn861Ser | antigen I/II family LPXTG-anchored adhesin | gene-EL140_RS07990 |
| Thr33Ala, Asp35Gly | ABC transporter permease | gene-EL140_RS00390 |
| Pro1264Leu | glycoside hydrolase N-terminal domain-containing protein | gene-EL140_RS08115 |
| Glu379Asp | penicillin-binding protein PBP2X | gene-EL140_RS08125 |
| Ser90Cys | phosphatase PAP2 family protein | gene-EL140_RS01360 |
| Glu871Asp | DNA-directed RNA polymerase subunit beta’ | gene-EL140_RS08440 |
| Predicted Amino Acid Change a | Predicted Protein Function | Gene ID |
|---|---|---|
| Glu144Gln | tagatose-bisphosphate aldolase | gene-EL140_RS04205 |
| Glu458Asp | fibronectin-binding SSURE repeat-containing protein | gene-EL140_RS00420 |
| Lys3Glu | PspC domain-containing protein | gene-EL140_RS00510 |
| Met6Leu | ribonuclease M5 | gene-EL140_RS01180 |
| Asn331Gln, Val2276Leu, His/Lys331Gln | accessory Sec-dependent serine-rich glycoprotein adhesin | gene-EL140_RS01855 |
| leu51Met, Ser151Ala, Arg165Lys | serine O-acetyltransferase | gene-EL140_RS02135 |
| Asn8Asp | nucleoside phosphorylase | gene-EL140_RS00405 |
| Asp43Glu | DUF1307 domain-containing protein | gene-EL140_RS02430 |
| Arg6Trp | DUF6287 domain-containing protein | gene-EL140_RS02620 |
| 599insAT | N-acetylmuramoyl-L-alanine amidase | gene-EL140_RS02630 |
| Ile53Val, His139Arg, Ser690Phe, Leu694Val | heavy metal translocating P-type ATPase | gene-EL140_RS00600 |
| Asn295Ser | hypothetical protein | gene-EL140_RS00190 |
| Gln992His | carbamoyl-phosphate synthase large subunit | gene-EL140_RS03330 |
| Glu438ins, GluGlnPro439HisProThr | DNA primase | gene-EL140_RS03860 |
| Arg51His, AspGln566GlyLeu, Val570Ala | glycoside hydrolase family 13 protein | gene-EL140_RS04725 |
| Glu229Lys | YjjG family noncanonical pyrimidine nucleotidase | gene-EL140_RS04975 |
| Asn142Asp | SUF system NifU family Fe-S cluster assembly protein | gene-EL140_RS05575 |
| Ala17Val | aminoglycoside 6-adenylyltransferase | gene-EL140_RS05640 |
| Phe93Cys | VanZ family protein | gene-EL140_RS05855 |
| Glu47Lys | UPF0223 family protein | gene-EL140_RS06150 |
| Asp65Glu | SDR family oxidoreductase | gene-EL140_RS06415 |
| Val294Ile | ribonuclease Z | gene-EL140_RS06420 |
| LeuAla83LeuThr, GlyLeu86AlaIle | hypothetical protein | gene-EL140_RS00190 |
| Asn350Lys | G5 domain-containing protein | gene-EL140_RS06930 |
| Ser824Asn, Thr1012Asn, Thr843Ile, Thr820Asn | G5 domain-containing protein | gene-EL140_RS06930 |
| Val1055Ala | exo-alpha-sialidase | gene-EL140_RS07065 |
| Val138Met | DUF421 domain-containing protein | gene-EL140_RS07435 |
| Asp1472Glu | glycoside hydrolase N-terminal domain-containing protein | gene-EL140_RS08115 |
| Asp871Glu | DNA-directed RNA polymerase subunit beta’ | gene-EL140_RS08440 |
| Cys610Gly | magnesium-translocating P-type ATPase | gene-EL140_RS08825 |
| Met267Ile | D-alanyl-lipoteichoic acid biosynthesis protein DltD | gene-EL140_RS09230 |
| Arg74Ser | hypothetical protein | gene-EL140_RS00190 |
| Ala862Thr | LPXTG cell wall anchor domain-containing protein | gene-EL140_RS06375 |
| Val137Leu | NAD(P)H-dependent glycerol-3-phosphate dehydrogenase | gene-EL140_RS00645 |
| Thr718Ala | HAD-IC family P-type ATPase | gene-EL140_RS01060 |
| LysValLeuValGly150AsnAlaLeuThrIle, Ala159Ser | sugar transferase | gene-EL140_RS01435 |
| IleAspLeu211ValAlaIle | metal ABC transporter ATP-binding protein | gene-EL140_RS02210 |
| Asn117Ser | DNA/RNA non-specific endonuclease | gene-EL140_RS01295 |
| Leu6Gln | NAD-dependent protein deacylase | gene-EL140_RS03745 |
| Asp74His | nucleotidyltransferase | gene-EL140_RS01975 |
| Gly14Ala | MBL fold metallo-hydrolase | gene-EL140_RS02220 |
| Lys12Gln | MmcQ/YjbR family DNA-binding protein | gene-EL140_RS05435 |
| Ala177Val | DNA topology modulation protein | gene-EL140_RS05725 |
| AsnLeu84ThrIle | prephenate dehydrogenase | gene-EL140_RS06020 |
| Asn78Ser, Thr72Ala | inositol monophosphatase family protein | gene-EL140_RS06145 |
| His772Asn, Asn770Tyr, Arg768Gln | Ltp family lipoprotein | gene-EL140_RS06460 |
| AlaAspVal352ValAlaIle | sensor histidine kinase | gene-EL140_RS01655 |
| Val24Glu | response regulator | gene-EL140_RS06480 |
| Phe200Tyr | Nif3-like dinuclear metal center hexameric protein | gene-EL140_RS06860 |
| Ter295Lysext a,* | ROK family protein | gene-EL140_RS02070 |
| Ala120Val, ThrIle109LeuLeu | ABC transporter permease | gene-EL140_RS00390 |
| Ser1567Thr, GlyThrGlyAla1549SerThrAsnVal | YSIRK-type signal peptide-containing protein | gene-EL140_RS06920 |
| Ala22Thr | 50S ribosomal protein L23 | gene-EL140_RS08720 |
| GluLeu68GlnAla, Val65Ile, Asn60Glu | phosphoribulokinase | gene-EL140_RS08745 |
| Ala123del | CDP-diacylglycerol—glycerol-3-phosphate 3-phosphatidyltransferase | gene-EL140_RS09460 |
| Pro15Leu | AraC family transcriptional regulator | gene-EL140_RS05010 |
| Met1fs | hypothetical protein | gene-EL140_RS00190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, N.N.; de Paula Baptista, R.; Tran, T.T.; Lapitan, C.K.; Garcia-de-la-Maria, C.; Miró, J.M.; Proctor, R.A.; Bayer, A.S. Membrane Phenotypic, Metabolic and Genotypic Adaptations of Streptococcus oralis Strains Destined to Rapidly Develop Stable, High-Level Daptomycin Resistance during Daptomycin Exposures. Antibiotics 2023, 12, 1083. https://doi.org/10.3390/antibiotics12071083
Mishra NN, de Paula Baptista R, Tran TT, Lapitan CK, Garcia-de-la-Maria C, Miró JM, Proctor RA, Bayer AS. Membrane Phenotypic, Metabolic and Genotypic Adaptations of Streptococcus oralis Strains Destined to Rapidly Develop Stable, High-Level Daptomycin Resistance during Daptomycin Exposures. Antibiotics. 2023; 12(7):1083. https://doi.org/10.3390/antibiotics12071083
Chicago/Turabian StyleMishra, Nagendra N., Rodrigo de Paula Baptista, Truc T. Tran, Christian K. Lapitan, Cristina Garcia-de-la-Maria, Jose M. Miró, Richard A. Proctor, and Arnold S. Bayer. 2023. "Membrane Phenotypic, Metabolic and Genotypic Adaptations of Streptococcus oralis Strains Destined to Rapidly Develop Stable, High-Level Daptomycin Resistance during Daptomycin Exposures" Antibiotics 12, no. 7: 1083. https://doi.org/10.3390/antibiotics12071083
APA StyleMishra, N. N., de Paula Baptista, R., Tran, T. T., Lapitan, C. K., Garcia-de-la-Maria, C., Miró, J. M., Proctor, R. A., & Bayer, A. S. (2023). Membrane Phenotypic, Metabolic and Genotypic Adaptations of Streptococcus oralis Strains Destined to Rapidly Develop Stable, High-Level Daptomycin Resistance during Daptomycin Exposures. Antibiotics, 12(7), 1083. https://doi.org/10.3390/antibiotics12071083