
Evaluation of Disinfectant Efficacy against Biofilm-Residing Wild-Type Salmonella from the Porcine Industry

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Screening of Strains from the Pig Industry for Biofilm Production, Morphotypes, and Phage Typing
2.2. Selection of Strains
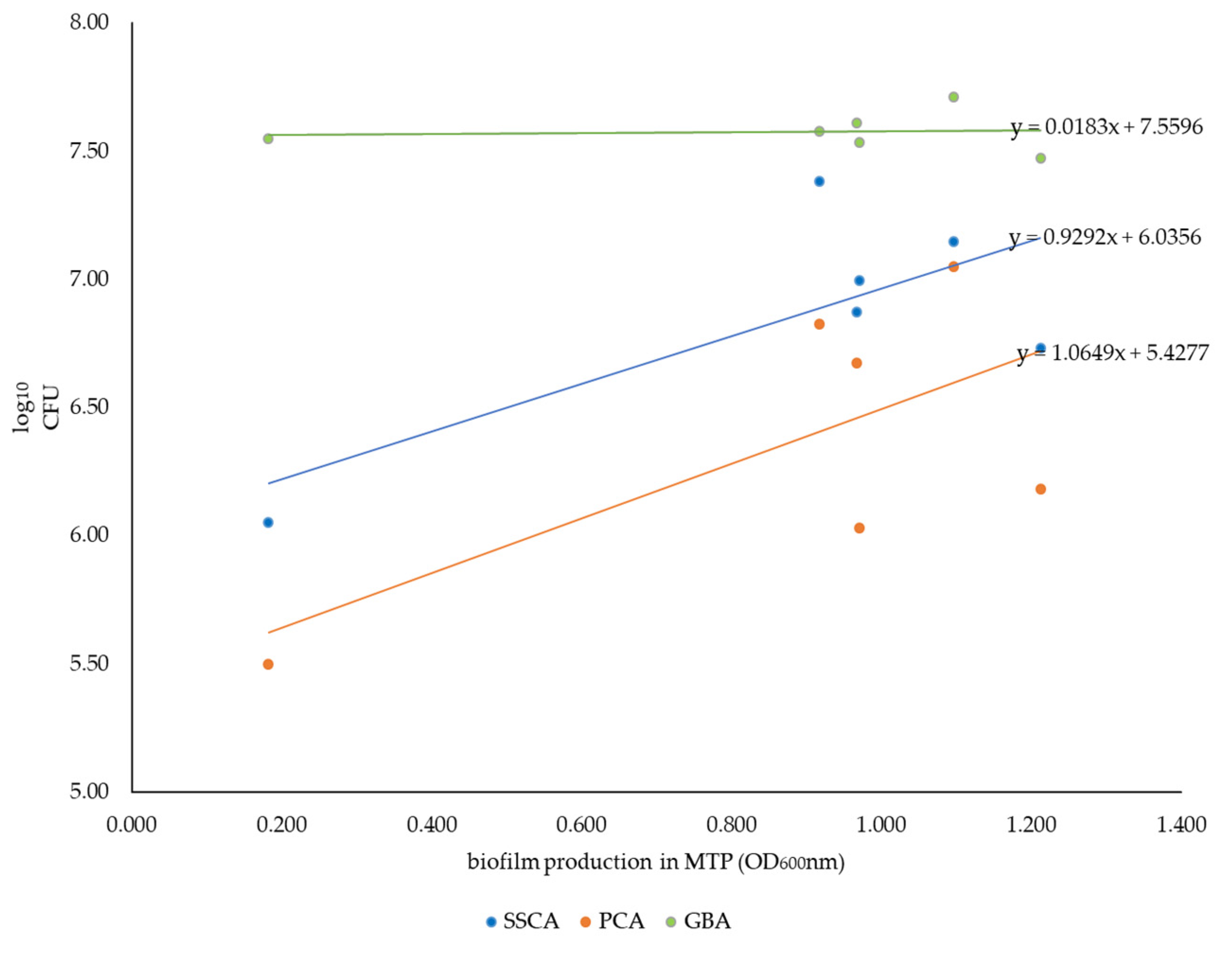
2.3. Correlation between Biofilm Formation Capacity in the MTP Assay and Achievable Cell Counts in Coupon/Bead Assays
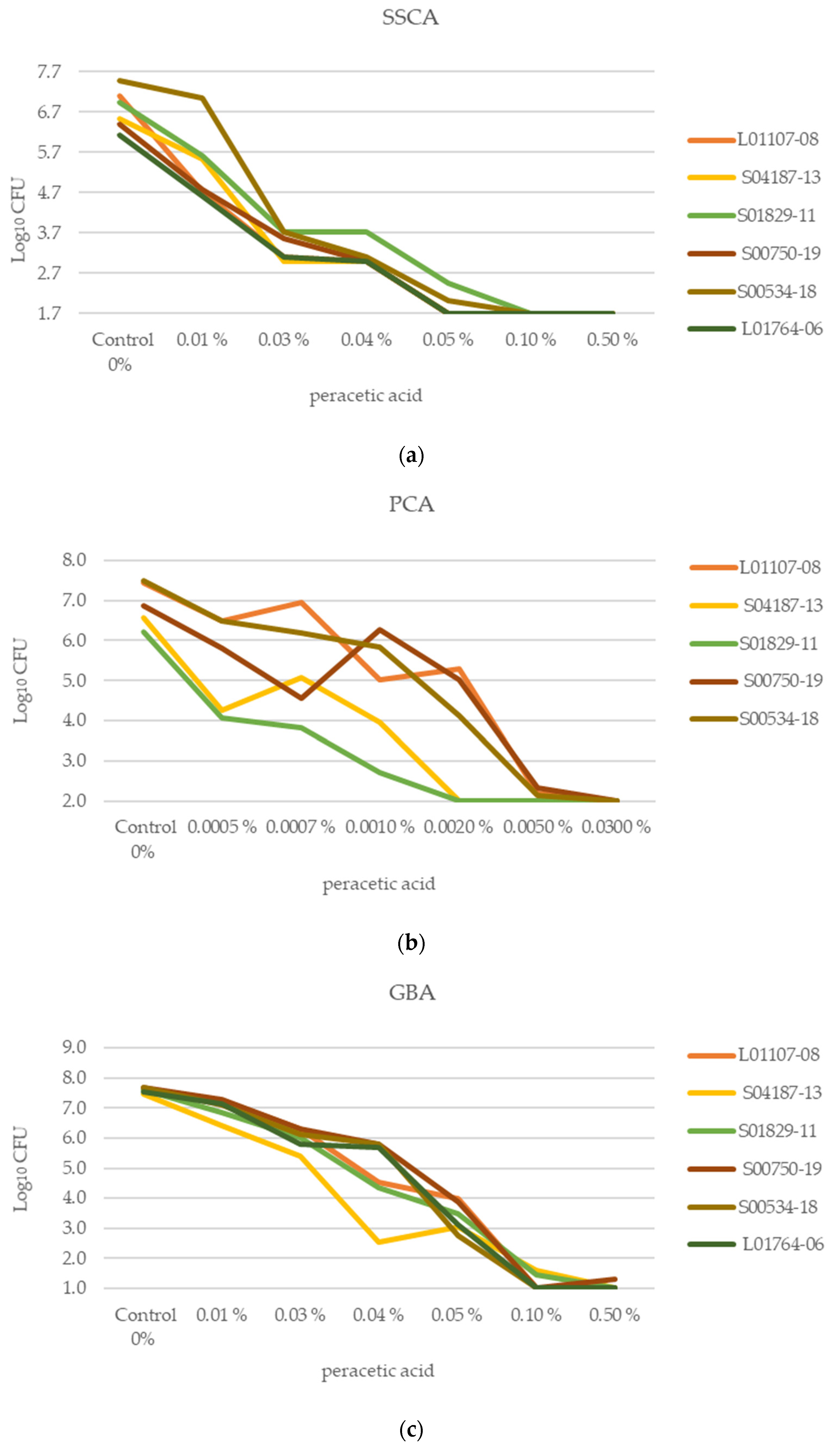
2.4. Effect of Disinfection on Biofilm of Selected Wild-Type Strains
2.4.1. Achievable CFU per Biofilm/mL of Separate Strains Following Disinfection
2.4.2. Mean log10 CFU Reduction after Disinfection
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Media
4.2. Biofilm Production on Polystyrene (Microtiter Plate Assay)
4.3. Biofilm Morphotyping
4.4. Disinfectant Testing
4.4.1. Study Design
4.4.2. Criteria for Successful Disinfection
4.4.3. Stainless-Steel Coupon Assay (SSCA)
4.4.4. Polyvinylchloride (PVC) Coupon Assay (PCA)
4.4.5. Glass Bead Assay (GBA)
4.5. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- Herrero-Fresno, A.; Olsen, J.E. Salmonella Typhimurium metabolism affects virulence in the host–A mini-review. Food Microbiol. 2018, 71, 98–110. [Google Scholar] [CrossRef]
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef]
- Ekperigin, H.E.; Nagaraja, K.V. Salmonella. Vet. Clin. N. Am. Food Anim. Pract. 1998, 14, 17–29. [Google Scholar] [CrossRef]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. mBio 2019, 10, 10–1128. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.G.; Tengelsen, L.A.; Smith, K.E.; Bender, J.B.; Frank, R.K.; Grendon, J.H.; Rice, D.H.; Thiessen, A.M.; Gilbertson, C.J.; Sivapalasingam, S.; et al. Multidrug-resistant Salmonella Typhimurium in four animal facilities. Emerg. Infect. Dis. 2005, 11, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Hanes, D. Nontyphoid Salmonella. In International Handbook of Foodborne Pathogens; CRC Press: Boca Raton, FL, USA, 2003; pp. 157–170. [Google Scholar]
- Rönnqvist, M.; Välttilä, V.; Ranta, J.; Tuominen, P. Salmonella risk to consumers via pork is related to the Salmonella prevalence in pig feed. Food Microbiol. 2018, 71, 93–97. [Google Scholar] [CrossRef]
- Kirchner, M.J.; Liebana, E.; McLaren, I.; Clifton-Hadley, F.A.; Wales, A.D.; Davies, R.H. Comparison of the environmental survival characteristics of Salmonella Dublin and Salmonella Typhimurium. Vet. Microbiol. 2012, 159, 509–514. [Google Scholar] [CrossRef]
- Jensen, A.N.; Dalsgaard, A.; Stockmarr, A.; Nielsen, E.M.; Baggesen, D.L. Survival and Transmission of Salmonella enterica Serovar Typhimurium in an Outdoor Organic Pig Farming Environment. Appl. Environ. Microbiol. 2006, 72, 1833–1842. [Google Scholar] [CrossRef] [Green Version]
- Vestby, L.K.; Møretrø, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonella are correlated with persistence in fish meal- and feed factories. BMC Vet. Res. 2009, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, D.C.; Fernandes Júnior, A.; Kaneno, R.; Silva, M.G.; Araújo Júnior, J.P.; Silva, N.C.; Rall, V.L. Ability of Salmonella spp. to produce biofilm is dependent on temperature and surface material. Foodborne Pathog. Dis. 2014, 11, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; Bloomfield, S.F. Survival of Salmonella in bathrooms and toilets in domestic homes following salmonellosis. J. Appl. Microbiol. 2000, 89, 137–144. [Google Scholar] [CrossRef]
- Jain, S.; Chen, J. Attachment and biofilm formation by various serotypes of Salmonella as influenced by cellulose production and thin aggregative fimbriae biosynthesis. J. Food Prot. 2007, 70, 2473–2479. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C. The perfect slime. Colloids Surf. B Biointerfaces 2011, 86, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef]
- Solano, C.; García, B.; Valle, J.; Berasain, C.; Ghigo, J.M.; Gamazo, C.; Lasa, I. Genetic analysis of Salmonella enteritidis biofilm formation: Critical role of cellulose. Mol. Microbiol. 2002, 43, 793–808. [Google Scholar] [CrossRef]
- Gosling, R.J.; Mawhinney, I.; Vaughan, K.; Davies, R.H.; Smith, R.P. Efficacy of disinfectants and detergents intended for a pig farm environment where Salmonella is present. Vet. Microbiol. 2017, 204, 46–53. [Google Scholar] [CrossRef]
- EN13697:2015+A12019; Chemical Disinfectants and Antiseptics, Quantitative Non-Porous Surface Test for the Evaluation of Bactericidal and/or Fungicidal Activity of Chemical Disinfectants Used in Food, Industrial, Domestic and Institutional Areas. Test Method and Requirements without Mechanical Action (Phase 2, Step 2). CEN-CENELEC Management Centre: Brussels, Belgium, 2019.
- Osland, A.M.; Vestby, L.K.; Nesse, L.L. The Effect of Disinfectants on Quinolone Resistant E. coli (QREC) in Biofilm. Microorganisms 2020, 8, 1831. [Google Scholar] [CrossRef]
- Stoller, A.; Stevens, M.J.A.; Stephan, R.; Guldimann, C. Characteristics of Listeria Monocytogenes Strains Persisting in a Meat Processing Facility over a 4-Year Period. Pathogens 2019, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.S.; Townsend, K.M.; Fenwick, S.G.; Trengove, R.D.; O’Handley, R.M. Comparative susceptibility of planktonic and 3-day-old Salmonella Typhimurium biofilms to disinfectants. J. Appl. Microbiol. 2010, 108, 2222–2228. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.M.; Konrat, K.; Osland, A.M.; Brook, E.; Oastler, C.; Vestby, L.K.; Gosling, R.J.; Nesse, L.L.; Arvand, M. Evaluation of Biofilm Cultivation Models for Efficacy Testing of Disinfectants against Salmonella Typhimurium Biofilms. Microorganisms 2023, 11, 761. [Google Scholar] [CrossRef]
- Römling, U.; Bokranz, W.; Rabsch, W.; Zogaj, X.; Nimtz, M.; Tschäpe, H. Occurrence and regulation of the multicellular morphotype in Salmonella serovars important in human disease. Int. J. Med. Microbiol. 2003, 293, 273–285. [Google Scholar] [CrossRef]
- Walia, K.; Argüello, H.; Lynch, H.; Grant, J.; Leonard, F.C.; Lawlor, P.G.; Gardiner, G.E.; Duffy, G. The efficacy of different cleaning and disinfection procedures to reduce Salmonella and Enterobacteriaceae in the lairage environment of a pig abattoir. Int. J. Food Microbiol. 2017, 246, 64–71. [Google Scholar] [CrossRef]
- Møretrø, T.; Vestby, L.K.; Nesse, L.L.; Storheim, S.E.; Kotlarz, K.; Langsrud, S. Evaluation of efficacy of disinfectants against Salmonella from the feed industry. J. Appl. Microbiol. 2009, 106, 1005–1012. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- EN 1656:2019; Chemical Disinfectants and Antiseptics-Quantitative Suspension Test for the Evaluation of Bactericidal Activity of Chemical Disinfectants and Antiseptics Used in the Veterinary Area–Test Method and Requirements (Phase 2, Step 1). CEN-CENELEC Management Centre: Brussels, Belgium, 2019.
- Corcoran, M.; Morris, D.; De Lappe, N.; O’Connor, J.; Lalor, P.; Dockery, P.; Cormican, M. Commonly used disinfectants fail to eradicate Salmonella enterica biofilms from food contact surface materials. Appl. Environ. Microbiol. 2014, 80, 1507–1514. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; Carballo, J.; García-Fernández, C.; Capita, R.; Alonso-Calleja, C. Structure and viability of 24- and 72-h-old biofilms formed by four pathogenic bacteria on polystyrene and glass contact surfaces. Food Microbiol. 2018, 76, 513–517. [Google Scholar] [CrossRef]
- Nesse, L.L.; Osland, A.M.; Mo, S.S.; Sekse, C.; Slettemeås, J.S.; Bruvoll, A.E.E.; Urdahl, A.M.; Vestby, L.K. Biofilm forming properties of quinolone resistant Escherichia coli from the broiler production chain and their dynamics in mixed biofilms. BMC Microbiol. 2020, 20, 46. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef] [PubMed]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.-J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [Green Version]
- Høiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.J.; Moser, C.; Jensen, P.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The clinical impact of bacterial biofilms. Int. J. Oral. Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Romeu, M.J.; Rodrigues, D.; Azeredo, J. Effect of sub-lethal chemical disinfection on the biofilm forming ability, resistance to antibiotics and expression of virulence genes of Salmonella enteritidis biofilm-surviving cells. Biofouling 2020, 36, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Vestby, L.K.; Møretrø, T.; Ballance, S.; Langsrud, S.; Nesse, L.L. Survival potential of wild type cellulose deficient Salmonella from the feed industry. BMC Vet. Res. 2009, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Scher, K.; Kesselman, E.; Shimoni, E.; Yaron, S. Morphological analysis of young and old pellicles of Salmonella Typhimurium. Biofouling 2007, 23, 385–394. [Google Scholar] [CrossRef]
- Allen, A.; Habimana, O.; Casey, E. The effects of extrinsic factors on the structural and mechanical properties of Pseudomonas fluorescens biofilms: A combined study of nutrient concentrations and shear conditions. Colloids Surf, B Biointerfaces 2018, 165, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, D.O.; Richter, A.M.; Hengge, R. Cellulose as an Architectural Element in Spatially Structured Escherichia coli Biofilms. J. Bacteriol. 2013, 195, 5540–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, S.E.; Sun, Y.; Secor, P.R.; Rhoads, D.D.; Wolcott, B.M.; James, G.A.; Wolcott, R.D. Survey of bacterial diversity in chronic wounds using pyrosequencing, DGGE, and full ribosome shotgun sequencing. BMC Microbiol. 2008, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Shatila, F.; Yaşa, İ.; Yalçın, H.T. Biofilm Formation by Salmonella enterica Strains. Curr. Microbiol. 2021, 78, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Fernandez-No, I.C.; Miranda, J.M.; Vázquez, B.; Cepeda, A.; Franco, C.M. Biofilm Formation and Morphotypes of Salmonella enterica subsp.arizonae Differs from Those of Other Salmonella enterica Subspecies in Isolates from Poultry Houses. J. Food Prot. 2016, 79, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ge, H.; Lin, W.; Song, Y.; Ge, F.; Huang, X.; Meng, X. Effect of different disinfection treatments on the adhesion and separation of biofilm on stainless steel surface. Water Sci. Technol. 2021, 83, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Farjami, A.; Hatami, M.S.; Siahi-Shadbad, M.R.; Lotfipour, F. Peracetic acid activity on biofilm formed by Escherichia coli isolated from an industrial water system. Lett. Appl. Microbiol. 2022, 74, 613–621. [Google Scholar] [CrossRef]
- Cordeiro, M.A.; Werle, C.H.; Milanez, G.P.; Yano, T. Curli fimbria: An Escherichia coli adhesin associated with human cystitis. Braz. J. Microbiol. 2016, 47, 414–416. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E.S.; Ward, L.R.; de Saxe, M.J.; de Sa, J.D.H. Bacteriophage-Typing Designations of Salmonella typhimurium. J. Hyg. 1977, 78, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Grimont, P.; Weill, F.-X. Antigenic Formulae of the Salmonella serovars, (9th ed.) Paris: WHO Collaborating Centre for Reference and Research on Salmonella. Inst. Pasteur. 2007, 9, 1–166. [Google Scholar]
- Ward, L.R.; de Sa, J.D.; Rowe, B. A phage-typing scheme for Salmonella enteritidis. Epidemiol. Infect. 1987, 99, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Malcova, M.; Hradecka, H.; Karpiskova, R.; Rychlik, I. Biofilm formation in field strains of Salmonella enterica serovar Typhimurium: Identification of a new colony morphology type and the role of SGI1 in biofilm formation. Vet. Microbiol. 2008, 129, 360–366. [Google Scholar] [CrossRef]
- Čabarkapa, I.; Škrinjar, M.; Lević, J.; Kokić, B.; Blagojev, N.; Milanov, D.; Suvajdžić, L. Biofilm forming ability of Salmonella enteritidis in vitro. Acta Vet. Beogr. 2015, 65, 371–389. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No | Mean OD600 ± SD | |
|---|---|---|
| S. Typhimurium | 35 | 0.851 ± 0.189 A |
| S. Derby | 15 | 0.189 ± 0.298 B |
| Morphotype | No | Mean OD600 ± SD | % of ST | % of SD |
|---|---|---|---|---|
| RDAR | 31 | 0.892 ± 0.294 A | 77.1 | 26.7 |
| BDAR | 6 | 0.507 ± 0.327 B | 17.1 | 0.0 |
| SAW | 13 | 0.149 ± 0.222 B | 5.7 | 73.3 |
| Strain | Serovar | Production Stage 1 | OD600nm in MTP Assay 2 | Morphology 3 | Phage Type |
|---|---|---|---|---|---|
| S04187-13 | S. Typhimurium | Finisher | 1.21 | RDAR | U288 |
| L01107-08 | S. Typhimurium | Finisher | 1.10 | RDAR | DT193 |
| S00750-19 | Monophasic S. Typhimurium | Finisher | 0.97 | RDAR | DT193 |
| S00534-18 | Monophasic S. Typhimurium | Sow/gilt | 0.92 | RDAR | DT193 |
| S01829-11 | S. Derby | Finisher | 0.97 | RDAR | n/a |
| L01764-06 | S. Typhimurium | Finisher | 0.18 | BDAR | U302 |
| Controls Mean Total log10CFU | Mean log10CFU Reduction ± CI 95% at Each Peracetic Acid Concentration | |||||||
|---|---|---|---|---|---|---|---|---|
| Assay | LOD a log10CFU | 0.01% | 0.03% | 0.04% | 0.05% | 0.10% | 0.50% | |
| SSCA | 1.7 | 6.76 ± 0.52 | 1.38 ± 0.67 | 3.40 ± 0.46 | 3.63 ± 0.53 | 4.88 ± 0.47 | 5.06 ± 0.53 | 5.06 ± 0.53 |
| GBA | 1 | 7.60 ± 0.10 | 0.59 ± 0.28 | 1.61 ± 0.28 | 2.83 ± 1.27 | 4.23 ± 0.48 | 6.43 ± 0.3 6 | 6.55 ± 0.11 |
| Controls Mean Total log10CFU | Mean log10CFU Reduction ± CI at each Peracetic Acid Concentration | |||||||
|---|---|---|---|---|---|---|---|---|
| Assay | LOD a log10CFU | 0.0005% | 0.0007% | 0.001% | 0.002% | 0.005% | 0.03% | |
| PCA | 2 | 6.92 ±0.59 | 1.49 ±0.83 | 1.61 ±0.98 | 2.16 ±1.35 | 3.24 ±1.50 | 4.79 ± 0.61 | 4.92 ± 0.69 |
| Controls Mean Total log10CFU | Mean log10CFU Reduction ± CI 95% at Each Glutaraldehyde Concentration | ||||||
|---|---|---|---|---|---|---|---|
| Assay | LOD a log10CFU | 0.01% | 0.05% | 0.10% | 0.50% | 1.00% | |
| SSCA | 1.7 | 6.97 ±0.54 | 0.51 ± 0.54 | 1.27 ± 0.47 | 2.45 ± 1.15 | 5.18 ± 0.59 | 5.27 ± 0.53 |
| PCA | 2 | 6.18 ±0.39 | 0.05 ± 0.25 | 2.26 ± 0.92 | 3.63 ± 0.81 | 4.18 ± 0.46 | b |
| GBA | 1 | 7.51 ±0.13 | 0.26 ± 0.12 | 1.13 ± 0.29 | 2.40 ± 1.04 | 4.21 ± 1.26 | 4.54 ± 1.44 |
| Glutaraldehyde | Peracetic Acid | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SSCA | 0.01% | 0.05% | 0.1% | 0.5% | 1% | 0.01% | 0.03% | 0.04% | 0.05% | 0.1% | 0.5% |
| GBA | 0.01% | 0.05% | 0.1% | 0.5% | 1% | 0.01% | 0.03% | 0.04% | 0.05% | 0.1% | 0.5% |
| PCA | 0.01% | 0.05% | 0.1% | 0.5% | - | 0.0005% | 0.0007% | 0.001% | 0.002% | 0.005% | 0.03% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osland, A.M.; Oastler, C.; Konrat, K.; Nesse, L.L.; Brook, E.; Richter, A.M.; Gosling, R.J.; Arvand, M.; Vestby, L.K. Evaluation of Disinfectant Efficacy against Biofilm-Residing Wild-Type Salmonella from the Porcine Industry. Antibiotics 2023, 12, 1189. https://doi.org/10.3390/antibiotics12071189
Osland AM, Oastler C, Konrat K, Nesse LL, Brook E, Richter AM, Gosling RJ, Arvand M, Vestby LK. Evaluation of Disinfectant Efficacy against Biofilm-Residing Wild-Type Salmonella from the Porcine Industry. Antibiotics. 2023; 12(7):1189. https://doi.org/10.3390/antibiotics12071189
Chicago/Turabian StyleOsland, Ane Mohr, Claire Oastler, Katharina Konrat, Live L. Nesse, Emma Brook, Anja M. Richter, Rebecca J. Gosling, Mardjan Arvand, and Lene K. Vestby. 2023. "Evaluation of Disinfectant Efficacy against Biofilm-Residing Wild-Type Salmonella from the Porcine Industry" Antibiotics 12, no. 7: 1189. https://doi.org/10.3390/antibiotics12071189
APA StyleOsland, A. M., Oastler, C., Konrat, K., Nesse, L. L., Brook, E., Richter, A. M., Gosling, R. J., Arvand, M., & Vestby, L. K. (2023). Evaluation of Disinfectant Efficacy against Biofilm-Residing Wild-Type Salmonella from the Porcine Industry. Antibiotics, 12(7), 1189. https://doi.org/10.3390/antibiotics12071189