
Acinetobacter Non-baumannii Species: Occurrence in Infections in Hospitalized Patients, Identification, and Antibiotic Resistance

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
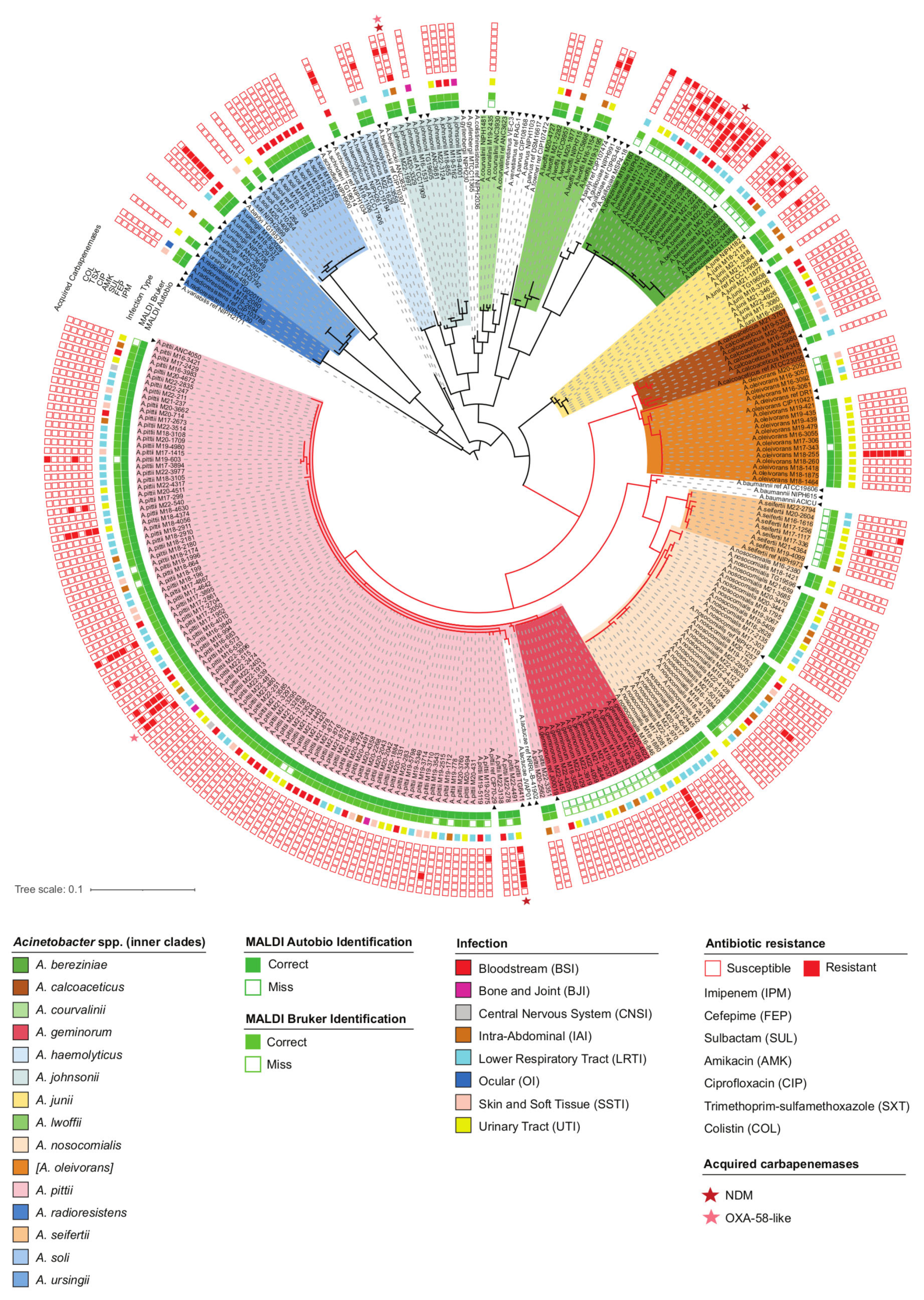
2.1. Species Distribution of Acinetobacter Isolates
2.2. Accuracy of Anb Species Identification Using MALDI-TOF MS Systems
2.3. Infections Caused by Anb Species
2.4. Susceptibility to Antibiotics and the Presence of Acquired Carbapenemase Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Species Identification with MALDI-TOF MS
4.3. Additional Tests to Distinguish A. baumannii
4.4. Sequencing of the rpoB Gene and Analysis of the Sequence Data
4.5. Antimicrobial Susceptibility Testing
4.6. Real-Time PCR Detection of Carbapenemase Genes
4.7. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-Based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, L.C.S.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a Global Pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis. 2017. Available online: https://www.who.int/publications/i/item/WHO-EMP-IAU-2017.12 (accessed on 17 July 2023).
- Wolf, S.; Barth-Jakschic, E.; Birkle, K.; Bader, B.; Marschal, M.; Liese, J.; Peter, S.; Oberhettinger, P. Acinetobacter geminorum sp. nov., Isolated from Human Throat Swabs. Int. J. Syst. Evol. Microbiol. 2021, 71, 005018. [Google Scholar] [CrossRef] [PubMed]
- Cosgaya, C.; Marí-Almirall, M.; Van Assche, A.; Fernández-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a Member of the Acinetobacter Calcoaceticus-Acinetobacter baumannii Complex Mainly Recovered from Clinical Samples in Different Countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.S.; Jung, J.; Jeon, C.O.; Park, W. Acinetobacter oleivorans sp. nov. Is Capable of Adhering to and Growing on Diesel-Oil. J. Microbiol. 2011, 49, 29–34. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Rooney, A.P. Acinetobacter dijkshoorniae Is a Later Heterotypic Synonym of Acinetobacter Lactucae. Int. J. Syst. Evol. Microbiol. 2018, 68, 131–132. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.K.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and Phenotypic Characterization of the Acinetobacter calcoaceticus–Acinetobacter baumannii Complex with the Proposal of Acinetobacter pittii sp. nov. (Formerly Acinetobacter Genomic Species 3) and Acinetobacter nosocomialis sp. nov. (Formerly Acinetobacter Genomic Species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef]
- Glover, J.S.; Ticer, T.D.; Engevik, M.A. Profiling Antibiotic Resistance in Acinetobacter calcoaceticus. Antibiotics 2022, 11, 978. [Google Scholar] [CrossRef] [PubMed]
- Diakite, A.; Dubourg, G.; Raoult, D. Updating the Repertoire of Cultured Bacteria from the Human Being. Microb. Pathog. 2021, 150, 104698. [Google Scholar] [CrossRef]
- Salzer, H.J.F.; Rolling, T.; Schmiede, S.; Klupp, E.M.; Lange, C.; Seifert, H. Severe Community-Acquired Bloodstream Infection with Acinetobacter ursingii in Person Who Injects Drugs. Emerg. Infect. Dis. 2016, 22, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chusri, S.; Chongsuvivatwong, V.; Rivera, J.I.; Silpapojakul, K.; Singkhamanan, K.; McNeil, E.; Doi, Y. Clinical Outcomes of Hospital-Acquired Infection with Acinetobacter nosocomialis and Acinetobacter pittii. Antimicrob. Agents Chemother. 2014, 58, 4172–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turton, J.F.; Shah, J.; Ozongwu, C.; Pike, R. Incidence of Acinetobacter Species Other than A. baumannii among Clinical Isolates of Acinetobacter: Evidence for Emerging Species. J. Clin. Microbiol. 2010, 48, 1445. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; Sharma, M.; Yadav, A.; Bala, K.; Chaudhary, U. Acinetobacter lwoffii an Emerging Pathogen in Neonatal ICU. Infect. Disord. Drug Targets 2015, 15, 184–188. [Google Scholar] [CrossRef]
- Fitzpatrick, M.A.; Ozer, E.; Bolon, M.K.; Hauser, A.R. Influence of ACB Complex Genospecies on Clinical Outcomes in a U.S. Hospital with High Rates of Multidrug Resistance. J. Infect. 2015, 70, 144. [Google Scholar] [CrossRef] [Green Version]
- Matsui, M.; Suzuki, S.; Yamane, K.; Suzuki, M.; Konda, T.; Arakawa, Y.; Shibayama, K. Distribution of Carbapenem Resistance Determinants among Epidemic and Non-Epidemic Types of Acinetobacter Species in Japan. J. Med. Microbiol. 2014, 63, 870–877. [Google Scholar] [CrossRef]
- Al Atrouni, A.; Joly-Guillou, M.L.; Hamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, S.; Biswas, I.; Veeraraghavan, B. Accurate Identification of Clinically Important Acinetobacter spp.: An Update. Future Sci. OA 2019, 5, FSO395. [Google Scholar] [CrossRef] [Green Version]
- La Scola, B.; Gundi, V.A.K.B.; Khamis, A.; Raoult, D. Sequencing of the RpoB Gene and Flanking Spacers for Molecular Identification of Acinetobacter Species. J. Clin. Microbiol. 2006, 44, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Jang, S.J.; Li, X.M.; Park, G.; Kook, J.K.; Kim, M.J.; Chang, Y.H.; Shin, J.H.; Kim, S.H.; Kim, D.M.; et al. Comparison of RpoB Gene Sequencing, 16S RRNA Gene Sequencing, GyrB Multiplex PCR, and the VITEK2 System for Identification of Acinetobacter Clinical Isolates. Diagn. Microbiol. Infect. Dis. 2014, 78, 29–34. [Google Scholar] [CrossRef]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by Detection of the BlaOXA-51-like Carbapenemase Gene Intrinsic to This Species. J. Clin. Microbiol. 2006, 44, 2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Chui, H.; Domish, L.; Hernandez, D.; Wang, G. Recent Development of Mass Spectrometry and Proteomics Applications in Identification and Typing of Bacteria. Proteomics Clin. Appl. 2016, 10, 346–357. [Google Scholar] [CrossRef]
- Marí-Almirall, M.; Cosgaya, C.; Higgins, P.G.; Van Assche, A.; Telli, M.; Huys, G.; Lievens, B.; Seifert, H.; Dijkshoorn, L.; Roca, I.; et al. MALDI-TOF/MS Identification of Species from the Acinetobacter baumannii (Ab) Group Revisited: Inclusion of the Novel A. seifertii and A. dijkshoorniae Species. Clin. Microbiol. Infect. 2017, 23, e1–e210. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tang, Y.; Lu, X. Insight into Identification of Acinetobacter Species by Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) in the Clinical Laboratory. J. Am. Soc. Mass. Spectrom. 2018, 29, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, M.; Nigro, S.J. Emergence, Molecular Mechanisms and Global Spread of Carbapenem-Resistant Acinetobacter baumannii. Microb. Genom. 2019, 5, e000306. [Google Scholar] [CrossRef]
- Li, P.; Yang, C.; Xie, J.; Liu, N.; Wang, H.; Zhang, L.; Wang, X.; Wang, Y.; Qiu, S.; Song, H. Acinetobacter calcoaceticus from a Fatal Case of Pneumonia Harboring BlaNDM-1 on a Widely Distributed Plasmid. BMC Infect. Dis. 2015, 15, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, F.M.; Rahman, N.I.A.; Ismail, S.; Abdullah, F.H.; Othman, N.; Alattraqchi, A.G.; Cleary, D.W.; Clarke, S.C.; Yeo, C.C. Prevalence and Antimicrobial Susceptibilities of Acinetobacter baumannii and Non-baumannii Acinetobacters from Terengganu, Malaysia and Their Carriage of Carbapenemase Genes. J. Med. Microbiol. 2018, 67, 1538–1543. [Google Scholar] [CrossRef]
- Lee, K.; Kim, M.N.; Choi, T.Y.; Cho, S.E.; Lee, S.; Whang, D.H.; Yong, D.; Chong, Y.; Woodford, N.; Livermore, D.M. Wide Dissemination of OXA-Type Carbapenemases in Clinical Acinetobacter spp. Isolates from South Korea. Int. J. Antimicrob. Agents 2009, 33, 520–524. [Google Scholar] [CrossRef]
- Tietgen, M.; Kramer, J.S.; Brunst, S.; Djahanschiri, B.; Wohra, S.; Higgins, P.G.; Weidensdorfer, M.; Riedel-Christ, S.; Pos, K.M.; Gonzaga, A.; et al. Identification of the Novel Class D β-Lactamase OXA-679 Involved in Carbapenem Resistance in Acinetobacter calcoaceticus. J. Antimicrob. Chemother. 2019, 74, 1494–1502. [Google Scholar] [CrossRef]
- Lasarte-Monterrubio, C.; Guijarro-Sánchez, P.; Bellés, A.; Vázquez-Ucha, J.C.; Arca-Suárez, J.; Fernández-Lozano, C.; Bou, G.; Beceiro, A. Carbapenem Resistance in Acinetobacter nosocomialis and Acinetobacter junii Conferred by Acquisition of BlaOXA-24/40 and Genetic Characterization of the Transmission Mechanism between Acinetobacter Genomic Species. Microbiol. Spectr. 2022, 10, e02734-21. [Google Scholar] [CrossRef]
- Singkham-In, U.; Chatsuwan, T. Mechanisms of Carbapenem Resistance in Acinetobacter pittii and Acinetobacter nosocomialis Isolates from Thailand. J. Med. Microbiol. 2018, 67, 1667–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, X.M.; Pan, Q.; Seifert, H.; Xing, X.W.; Yuan, J.; Zhou, Z.Y.; Luo, X.Y.; Liu, H.M.; Xie, Y.L.; Yang, L.Q.; et al. First Identification of Multidrug-Resistant Acinetobacter bereziniae Isolates Harboring BlaNDM-1 from Hospitals in South China. Heliyon 2023, 9, e12365. [Google Scholar] [CrossRef]
- Tavares, L.C.B.; Cunha, M.P.V.; De Vasconcellos, F.M.; Bertani, A.M.D.J.; De Barcellos, T.A.F.; Bueno, M.S.; Santos, C.A.; Sant’Ana, D.A.; Ferreira, A.M.; Mondelli, A.L.; et al. Genomic and Clinical Characterization of IMP-1-Producing Multidrug-Resistant Acinetobacter bereziniae Isolates from Bloodstream Infections in a Brazilian Tertiary Hospital. Microb. Drug Resist. 2020, 26, 1399–1404. [Google Scholar] [CrossRef]
- Cayô, R.; Rodrigues-Costa, F.; Matos, A.P.; Carvalhaes, C.G.; Dijkshoorn, L.; Gales, A.C. Old Clinical Isolates of Acinetobacter seifertii in Brazil Producing OXA-58. Antimicrob. Agents Chemother. 2016, 60, 2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaña, S.; Palombarani, S.; Carulla, M.; Kunst, A.; Rodriguez, C.H.; Nastro, M.; Vay, C.; Ramirez, M.S.; Almuzara, M. First Case of Bacteraemia Due to Acinetobacter schindleri Harbouring BlaNDM-1 in an Immunocompromised Patient. New Microbes New Infect. 2018, 21, 28. [Google Scholar] [CrossRef]
- Cayô, R.; Streling, A.P.; Nodari, C.S.; Matos, A.P.; De Paula Luz, A.; Dijkshoorn, L.; Pignatari, A.C.C.; Gales, A.C. Occurrence of IMP-1 in Non-baumannii Acinetobacter Clinical Isolates from Brazil. J. Med. Microbiol. 2018, 67, 628–630. [Google Scholar] [CrossRef]
- Park, Y.K.; Jung, S.I.; Park, K.H.; Kim, S.H.; Ko, K.S. Characteristics of Carbapenem-Resistant Acinetobacter spp. Other than Acinetobacter baumannii in South Korea. Int. J. Antimicrob. Agents 2012, 39, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.Y.; Chen, C.; Liu, B.T.; He, Q.; Wu, X.T.; Sun, R.Y.; Zhang, Y.; Cui, Z.H.; Guo, W.Y.; Jia, Q.L.; et al. Co-Occurrence of Plasmid-Mediated Tigecycline and Carbapenem Resistance in Acinetobacter spp. from Waterfowls and Their Neighboring Environment. Antimicrob. Agents Chemother. 2020, 64, 02502-19. [Google Scholar] [CrossRef]
- Kimura, Y.; Harada, K.; Shimizu, T.; Sato, T.; Kajino, A.; Usui, M.; Tamura, Y.; Tsuyuki, Y.; Miyamoto, T.; Ohki, A.; et al. Species Distribution, Virulence Factors, and Antimicrobial Resistance of Acinetobacter spp. Isolates from Dogs and Cats: A Preliminary Study. Microbiol. Immunol. 2018, 62, 462–466. [Google Scholar] [CrossRef] [Green Version]
- Baraka, A.; Traglia, G.M.; Montaña, S.; Tolmasky, M.E.; Ramirez, M.S. An Acinetobacter Non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs). Antibiotics 2021, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Radolfova-Krizova, L.; Maixnerova, M.; Vrestiakova, E.; Jezek, P.; Sedo, O. Taxonomy of Haemolytic and/or Proteolytic Strains of the Genus Acinetobacter with the Proposal of Acinetobacter courvalinii sp. nov. (Genomic Species 14 Sensu Bouvet & Jeanjean), Acinetobacter dispersus sp. nov. (Genomic Species 17), Acinetobacter modestus sp. nov., Acinetobacter proteolyticus sp. nov. and Acinetobacter vivianii sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 1673–1685. [Google Scholar] [CrossRef]
- Dey, D.K.; Park, J.; Kang, S.C. Genotypic, Phenotypic, and Pathogenic Characterization of the Soil Isolated Acinetobacter courvalinii. Microb. Pathog. 2020, 149, 104287. [Google Scholar] [CrossRef]
- Poirel, L.; Figueiredo, S.; Cattoir, V.; Carattoli, A.; Nordmann, P. Acinetobacter Radioresistens as a Silent Source of Carbapenem Resistance for Acinetobacter spp. Antimicrob. Agents Chemother. 2008, 52, 1252–1256. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenkov, A.Y.; Trushin, I.V.; Vinogradova, A.G.; Avramenko, A.A.; Sukhorukova, M.V.; Malhotra-Kumar, S.; Dekhnich, A.V.; Edelstein, M.V.; Kozlov, R.S. AMRmap: An Interactive Web Platform for Analysis of Antimicrobial Resistance Surveillance Data in Russia. Front. Microbiol. 2021, 12, 620002. [Google Scholar] [CrossRef]
- Gkorezis, P.; Rineau, F.; Van Hamme, J.; Franzetti, A.; Daghio, M.; Thijs, S.; Weyens, N.; Vangronsveld, J. Draft Genome Sequence of Acinetobacter Oleivorans PF1, a Diesel-Degrading and Plant-Growth-Promoting Endophytic Strain Isolated from Poplar Trees Growing on a Diesel-Contaminated Plume. Genome Announc. 2015, 3, e01430-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deems, A.; Du Prey, M.; Dowd, S.E.; McLaughlin, R.W. Characterization of the Biodiesel Degrading Acinetobacter oleivorans Strain PT8 Isolated from the Fecal Material of a Painted Turtle (Chrysemys picta). Curr. Microbiol. 2021, 78, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wei, H.; Ke, T.; Fu, Y.; Zeng, Y.; Chen, C.; Chen, L. Characterization and Genome Analysis of Acinetobacter oleivorans S4 as an Efficient Hydrocarbon-Degrading and Plant-Growth-Promoting rhizobacterium. Chemosphere 2023, 331, 138732. [Google Scholar] [CrossRef]
- Park, J.H.; Jang, Y.; Yoon, I.; Kim, T.S.; Park, H. Comparison of Autof Ms1000 and Bruker Biotyper MALDI-TOF MS Platforms for Routine Identification of Clinical Microorganisms. Biomed. Res. Int. 2021, 2021, 6667623. [Google Scholar] [CrossRef]
- Kozlov, R.S.; Azizov, I.S.; Dekhnich, V.; Ivanchik, N.V.; Kuzmenkov, Y.; Martinovich; Mikotina, V.; Sukhorukova, V.; Trushin, I.V.; Edelstein, V. In Vitro Activity of Biapenem and Other Carbapenems against Russian Clinical Isolates of Pseudomonas aeruginosa, Acinetobacter spp., and Enterobacterales. Klin. Mikrobiol. Antimikrobn. Himioter. 2021, 23, 280–291. [Google Scholar] [CrossRef]
- Shek, E.A.; Sukhorukova, M.V.; Edelstein, M.V.; Skleenova, E.Y.; Ivanchik, N.V.; Shajdullina, E.R.; Kuzmenkov, A.Y.; Dekhnich, A.V.; Kozlov, R.S.; Semyonova, N.V.; et al. Antimicrobial Resistance, Carbapenemase Production, and Genotypes of Nosocomial Acinetobacter spp. Isolates in Russia: Results of Multicenter Epidemiological Study “MARATHON 2015–2016”. Klin. Mikrobiol. Antimikrobn. Himioter. 2019, 21, 171–180. [Google Scholar] [CrossRef]
- Jayol, A.; Poirel, L.; Brink, A.; Villegas, M.V.; Yilmaz, M.; Nordmann, P. Resistance to Colistin Associated with a Single Amino Acid Change in Protein PmrB among Klebsiella pneumoniae Isolates of Worldwide Origin. Antimicrob. Agents Chemother. 2014, 58, 4762–4766. [Google Scholar] [CrossRef] [PubMed]
- Brovedan, M.; Repizo, G.D.; Marchiaro, P.; Viale, A.M.; Limansky, A. Characterization of the Diverse Plasmid Pool Harbored by the BlaNDM-1-Containing Acinetobacter bereziniae HPC229 Clinical Strain. PLoS ONE 2019, 14, e0220584. [Google Scholar] [CrossRef] [Green Version]
- Brovedan, M.; Marchiaro, P.M.; Morán-Barrio, J.; Revale, S.; Cameranesi, M.; Brambilla, L.; Viale, A.M.; Limansky, A.S. Draft Genome Sequence of Acinetobacter bereziniae HPC229, a Carbapenem-Resistant Clinical Strain from Argentina Harboring BlaNDM-1. Genome Announc. 2016, 4, 00117-16. [Google Scholar] [CrossRef] [Green Version]
- Kamolvit, W.; Higgins, P.G.; Paterson, D.L.; Seifert, H. Multiplex PCR to Detect the Genes Encoding Naturally Occurring Oxacillinases in Acinetobacter spp. J. Antimicrob. Chemother. 2014, 69, 959–963. [Google Scholar] [CrossRef] [Green Version]
- Marqué, S.; Poirel, L.; Héritier, C.; Brisse, S.; Blasco, M.D.; Filip, R.; Coman, G.; Naas, T.; Nordmann, P. Regional Occurrence of Plasmid-Mediated Carbapenem-Hydrolyzing Oxacillinase OXA-58 in Acinetobacter spp. in Europe. J. Clin. Microbiol. 2005, 43, 4885–4888. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.; Woodford, N.; Afzal-Shah, M.; Livermore, D. Occurrence of OXA-58-like Carbapenemases in Acinetobacter spp. Collected over 10 Years in Three Continents. Antimicrob. Agents Chemother. 2006, 50, 756–758. [Google Scholar] [CrossRef]
- Mendes, R.E.; Bell, J.M.; Turnidge, J.D.; Castanheira, M.; Deshpande, L.M.; Jones, R.N. Codetection of BlaOXA-23-like Gene (BlaOXA-133) and BlaOXA-58 in Acinetobacter Radioresistens: Report from the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2009, 53, 843–844. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Franklin, C.; Walters, L.J.; Bell, J.M.; Spelman, D.W. OXA-58 and IMP-4 Carbapenem-Hydrolyzing Beta-Lactamases in an Acinetobacter junii Blood Culture Isolate from Australia. Antimicrob. Agents Chemother. 2006, 50, 399–400. [Google Scholar] [CrossRef]
- Huang, L.Y.; Lu, P.L.; Chen, T.L.; Chang, F.Y.; Fung, C.P.; Siu, L.K. Molecular Characterization of Beta-Lactamase Genes and Their Genetic Structures in Acinetobacter Genospecies 3 Isolates in Taiwan. Antimicrob. Agents Chemother. 2010, 54, 2699–2703. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yang, P.; Wang, X.; Zong, Z. Characterization of Acinetobacter johnsonii Isolate XBB1 Carrying Nine Plasmids and Encoding NDM-1, OXA-58 and PER-1 by Genome Sequencing. J. Antimicrob. Chemother. 2016, 71, 71–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Zhang, X.; Zhou, Y.; Zhang, Z.; Zheng, X. Whole-Genome Sequencing of an NDM-1- and OXA-58-Producing Acinetobacter towneri Isolate from Hospital Sewage in Sichuan Province, China. J. Glob. Antimicrob. Resist. 2019, 16, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Endo, S.; Nakano, R.; Nakano, A.; Saito, K.; Kakuta, R.; Kakuta, N.; Horiuchi, S.; Yano, H.; Kaku, M. Emergence of IMP-34- and OXA-58-Producing Carbapenem-Resistant Acinetobacter colistiniresistens. Antimicrob. Agents Chemother. 2019, 63, e02633-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrickx, A.P.A.; Schade, R.P.; Landman, F.; Bosch, T.; Schouls, L.M.; van Dijk, K. Comparative Analysis of IMP-4- and OXA-58-Containing Plasmids of Three Carbapenemase-Producing Acinetobacter ursingii Strains in the Netherlands. J. Glob. Antimicrob. Resist. 2022, 31, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Fávaro, L.D.S.; de Paula-Petroli, S.B.; Romanin, P.; Tavares, E.D.R.; Ribeiro, R.A.; Hungria, M.; de Oliveira, A.G.; Yamauchi, L.M.; Yamada-Ogatta, S.F.; Carrara-Marroni, F.E. Detection of OXA-58-Producing Acinetobacter bereziniae in Brazil. J. Glob. Antimicrob. Resist. 2019, 19, 53–55. [Google Scholar] [CrossRef]
- Ang, G.Y.; Yu, C.Y.; Cheong, Y.M.; Yin, W.F.; Chan, K.G. Emergence of ST119 Acinetobacter pittii Co-Harbouring NDM-1 and OXA-58 in Malaysia. Int. J. Antimicrob. Agents 2016, 47, 168–169. [Google Scholar] [CrossRef]
- Strateva, T.; Sirakov, I.; Savov, E.; Mitov, I. First Detection of an OXA-58 Carbapenemase-Producing Acinetobacter nosocomialis Clinical Isolate in the Balkan States. J. Glob. Antimicrob. Resist. 2018, 13, 123–124. [Google Scholar] [CrossRef]
- Tognim, M.C.B.; Gales, A.C.; Penteado, A.P.; Silbert, S.; Sader, H.S. Dissemination of IMP-1 Metallo- Beta -Lactamase-Producing Acinetobacter Species in a Brazilian Teaching Hospital. Infect. Control Hosp. Epidemiol. 2006, 27, 742–747. [Google Scholar] [CrossRef]
- Lee, K.; Kim, C.K.; Hong, S.G.; Choi, J.; Song, S.; Koh, E.; Yong, D.; Jeong, S.H.; Yum, J.H.; Docquier, J.D.; et al. Characteristics of Clinical Isolates of Acinetobacter Genomospecies 10 Carrying Two Different Metallo-Beta-Lactamases. Int. J. Antimicrob. Agents 2010, 36, 259–263. [Google Scholar] [CrossRef]
- Hu, H.; Hu, Y.; Pan, Y.; Liang, H.; Wang, H.; Wang, X.; Hao, Q.; Yang, X.; Yang, X.; Xiao, X.; et al. Novel Plasmid and Its Variant Harboring Both a Bla(NDM-1) Gene and Type IV Secretion System in Clinical Isolates of Acinetobacter lwoffii. Antimicrob. Agents Chemother. 2012, 56, 1698–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, C.; Zhang, Q.; Qi, J.; Liu, H.; Wang, Y.; He, T.; Ma, L.; Lai, J.; Shen, Z.; et al. Identification of New Delhi Metallo-β-Lactamase 1 in Acinetobacter lwoffii of Food Animal Origin. PLoS ONE 2012, 7, e37152. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Hotta, G.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Regional Dissemination of Acinetobacter Species Harbouring Metallo-β-Lactamase Genes in Japan. Clin. Microbiol. Infect. 2013, 19, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Alattraqchi, A.G.; Mohd Rani, F.; Rahman, N.I.A.; Ismail, S.; Cleary, D.W.; Clarke, S.C.; Yeo, C.C. Complete Genome Sequencing of Acinetobacter baumannii AC1633 and Acinetobacter nosocomialis AC1530 Unveils a Large Multidrug-Resistant Plasmid Encoding the NDM-1 and OXA-58 Carbapenemases. mSphere 2021, 6, e01076-20. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, P.; Huang, H.; Huang, Y.; Wu, Z.; Liao, K. Detection of Co-Harboring OXA-58 and NDM-1 Carbapenemase Producing Genes Resided on a Same Plasmid from an Acinetobacter pittii Clinical Isolate in China. Iran. J. Basic Med. Sci. 2019, 22, 106–111. [Google Scholar] [CrossRef]
- Cheikh, H.B.; Domingues, S.; Silveira, E.; Kadri, Y.; Rosário, N.; Mastouri, M.; Da Silva, G.J. Molecular Characterization of Carbapenemases of Clinical Acinetobacter baumannii-calcoaceticus Complex Isolates from a University Hospital in Tunisia. 3 Biotech. 2018, 8, 297. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, X.; Meng, X.; Zhang, G.; Wang, J.; Zhou, D.; Guo, X. “Roar” of BlaNDM-1 and “Silence” of BlaOXA-58 Co-Exist in Acinetobacter pittii. Sci. Rep. 2015, 5, 8976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, H.; Cai, K.; Yu, P.; Liu, Y.; Zhao, G.; Chen, R.; Xu, R.; Yu, M. Evaluation of Three Sample Preparation Methods for the Identification of Clinical Strains by Using Two MALDI-TOF MS Systems. J. Mass Spectrom. 2021, 56, e4696. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Methods for the Identification of Cultured Microorganisms Using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry, 1st ed.; CLSI supplement M58; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The Population Structure of Acinetobacter baumannii: Expanding Multiresistant Clones from an Ancestral Susceptible Genetic Pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [Green Version]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.D.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a Multilocus Sequence Typing Scheme for Characterization of Clinical Isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382. [Google Scholar] [CrossRef] [Green Version]
- Gundi, V.A.K.B.; Dijkshoorn, L.; Burignat, S.; Raoult, D.; La Scola, B. Validation of Partial RpoB Gene Sequence Analysis for the Identification of Clinically Important and Emerging Acinetobacter Species. Microbiology 2009, 155, 2333–2341. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- ISO 20776-1:2019; Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Broth Micro-Dilution Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases. ISO: Geneva, Switzerland, 2019.
- The European Committee on Antimicrobial Susceptibility Testing. EUCAST Reading Guide for Broth Microdilution v. 4.0. 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2022_manuals/Reading_guide_BMD_v_4.0_2022.pdf (accessed on 17 July 2023).
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.1. 2023. Available online: http://www.eucast.org (accessed on 17 July 2023).
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed.; CLSI supplement M100; CLSI: Wayne, PA, USA, 2023. [Google Scholar]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Anb Species 1 | No. of Isolates | No. (Percent) Correctly Identified | |
|---|---|---|---|
| Autobio Autof | Bruker Biotyper | ||
| A. bereziniae | 18 | 0 (0%) | 18 (100%) |
| A. calcoaceticus/oleivorans | 21 | 17 (81%) | 21 (100%) |
| A. courvalinii | 1 | 0 (0%) | 1 (100%) |
| A. geminorum | 14 | 0 (0%) | 0 (0%) |
| A. haemolyticus | 3 | 3 (100%) | 3 (100%) |
| A. johnsonii | 7 | 7 (100%) | 7 (100%) |
| A. junii | 9 | 9 (100%) | 9 (100%) |
| A. lwoffii | 5 | 5 (100%) | 5 (100%) |
| A. nosocomialis | 32 | 32 (100%) | 30 (93.8%) |
| A. pittii | 100 | 100 (100%) | 89 (89%) |
| A. radioresistens | 2 | 2 (100%) | 2 (100%) |
| A. seifertii | 8 | 0 (0%) | 8 (100%) |
| A. soli | 9 | 9 (100%) | 9 (100%) |
| A. ursingii/septicus | 5 | 5 (100%) | 5 (100%) |
| Antibiotic | MIC, mg/L | Percent by Category 1 | ||||
|---|---|---|---|---|---|---|
| Range | 50% | 90% | S | I | R | |
| Amikacin | ≤0.5–≥256 | 1 | 8 | 90.1 | - | 9.9 |
| Cefepime 2 | ≤0.5–≥256 | 2 | 16 | 88.7 | 5.7 | 5.7 |
| Ciprofloxacin | ≤0.06–≥128 | 0.125 | 0.5 | - | 92.6 | 7.4 |
| Colistin | ≤0.06–≥64 | 0.5 | 2 | 97.0 | - | 3.0 |
| Gentamicin | ≤0.25–≥256 | 0.5 | 8 | 88.8 | - | 11.2 |
| Imipenem | ≤0.06–≥128 | 0.125 | 0.5 | 95.7 | - | 4.3 |
| Meropenem | ≤0.06–≥128 | 0.25 | 0.5 | 95.2 | 0.9 | 3.9 |
| Sulbactam 3 | ≤0.25–≥256 | 1 | 4 | 92.2 | 2.2 | 5.6 |
| Tigecycline 4 | ≤0.03–≥16 | 0.25 | 1 | - | - | - |
| Tobramycin | ≤0.25–≥256 | 0.25 | 2 | 91.3 | - | 8.7 |
| Trimethoprim– sulfamethoxazole (1:19) 5 | ≤0.125–≥256 | 0.125 | 4 | 89.6 | 1.7 | 8.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheck, E.; Romanov, A.; Shapovalova, V.; Shaidullina, E.; Martinovich, A.; Ivanchik, N.; Mikotina, A.; Skleenova, E.; Oloviannikov, V.; Azizov, I.; et al. Acinetobacter Non-baumannii Species: Occurrence in Infections in Hospitalized Patients, Identification, and Antibiotic Resistance. Antibiotics 2023, 12, 1301. https://doi.org/10.3390/antibiotics12081301
Sheck E, Romanov A, Shapovalova V, Shaidullina E, Martinovich A, Ivanchik N, Mikotina A, Skleenova E, Oloviannikov V, Azizov I, et al. Acinetobacter Non-baumannii Species: Occurrence in Infections in Hospitalized Patients, Identification, and Antibiotic Resistance. Antibiotics. 2023; 12(8):1301. https://doi.org/10.3390/antibiotics12081301
Chicago/Turabian StyleSheck, Eugene, Andrey Romanov, Valeria Shapovalova, Elvira Shaidullina, Alexey Martinovich, Natali Ivanchik, Anna Mikotina, Elena Skleenova, Vladimir Oloviannikov, Ilya Azizov, and et al. 2023. "Acinetobacter Non-baumannii Species: Occurrence in Infections in Hospitalized Patients, Identification, and Antibiotic Resistance" Antibiotics 12, no. 8: 1301. https://doi.org/10.3390/antibiotics12081301
APA StyleSheck, E., Romanov, A., Shapovalova, V., Shaidullina, E., Martinovich, A., Ivanchik, N., Mikotina, A., Skleenova, E., Oloviannikov, V., Azizov, I., Vityazeva, V., Lavrinenko, A., Kozlov, R., & Edelstein, M. (2023). Acinetobacter Non-baumannii Species: Occurrence in Infections in Hospitalized Patients, Identification, and Antibiotic Resistance. Antibiotics, 12(8), 1301. https://doi.org/10.3390/antibiotics12081301