
Isolation, Identification, and Antibacterial Properties of Prodigiosin, a Bioactive Product Produced by a New Serratia marcescens JSSCPM1 Strain: Exploring the Biosynthetic Gene Clusters of Serratia Species for Biological Applications

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Isolation of Red-Pigmented Bacteria and Biochemical Characterization
2.2. 16S rRNA Sequencing Report and Phylogenetic Tree Analysis
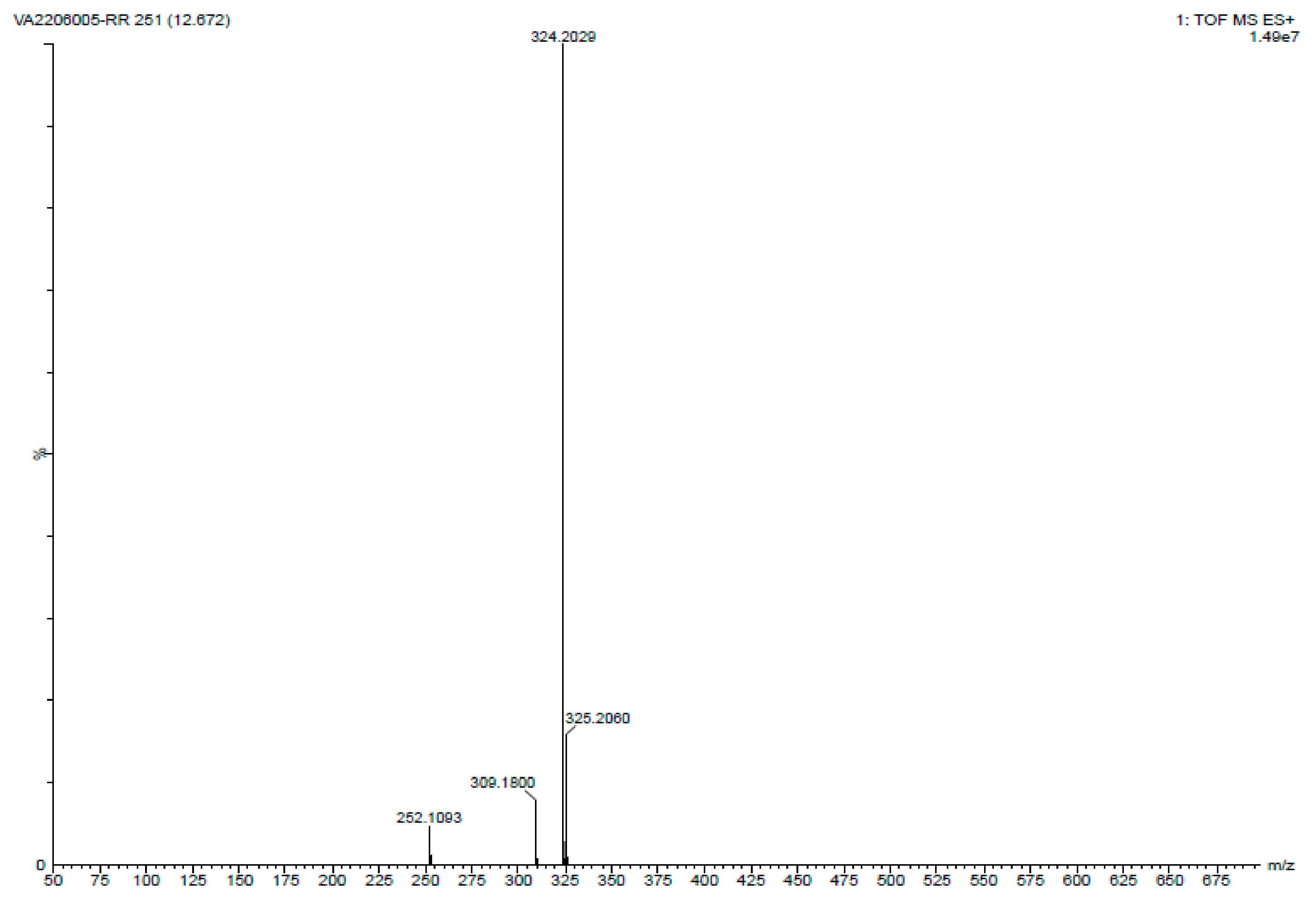
2.3. Analyzing the PG Compound
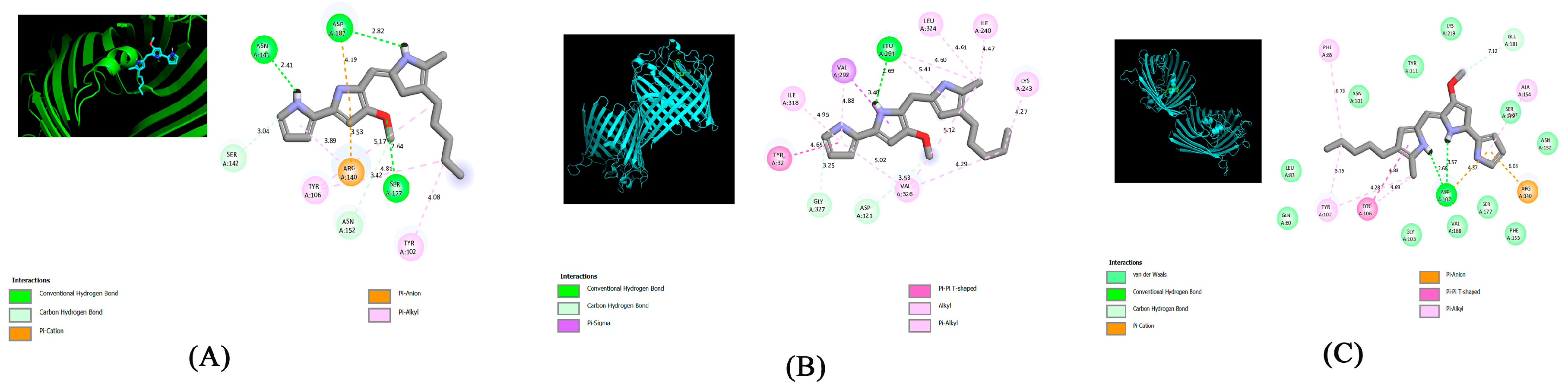
2.4. Molecular Docking Analysis
2.5. Antimicrobial Assay Evaluation Report
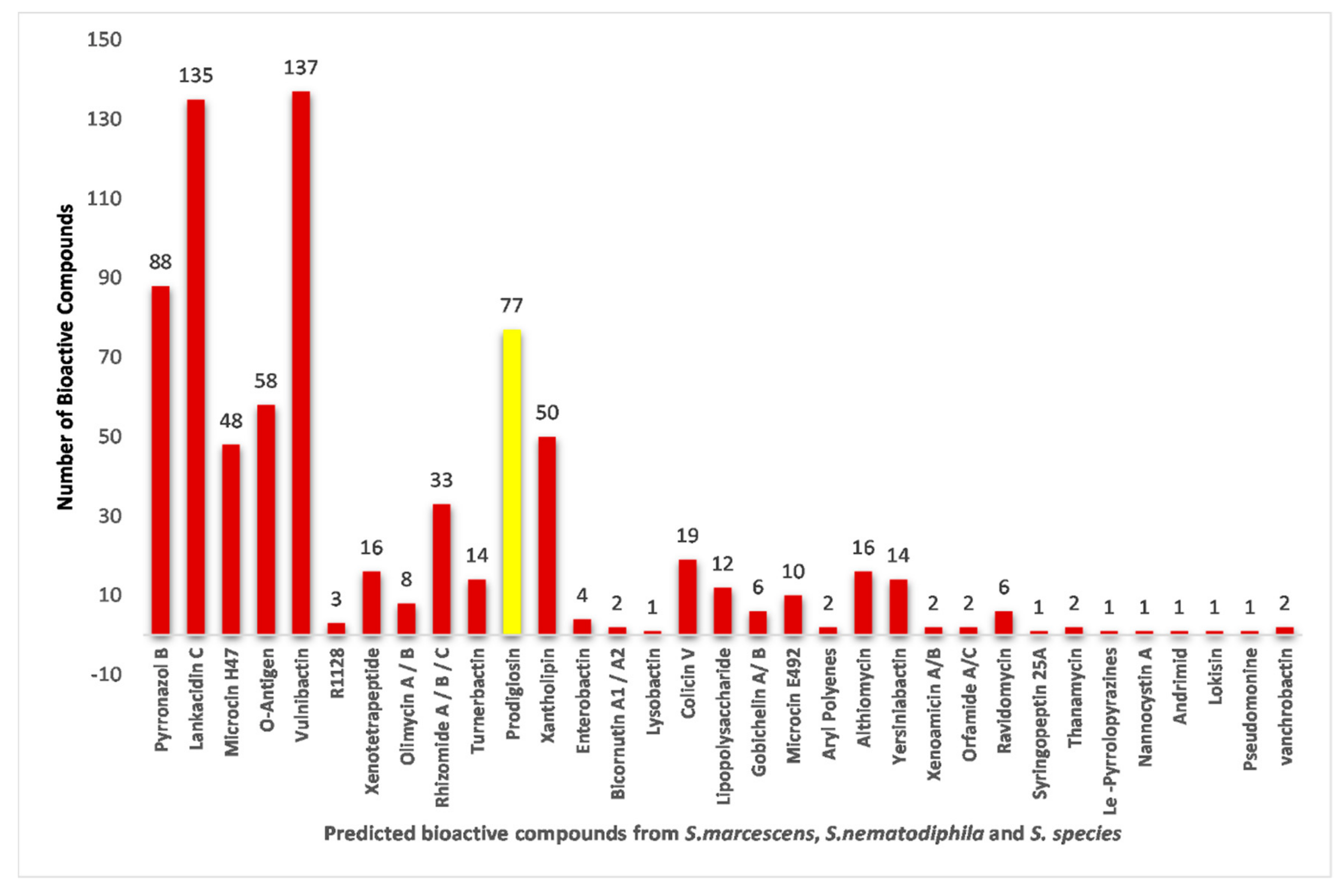
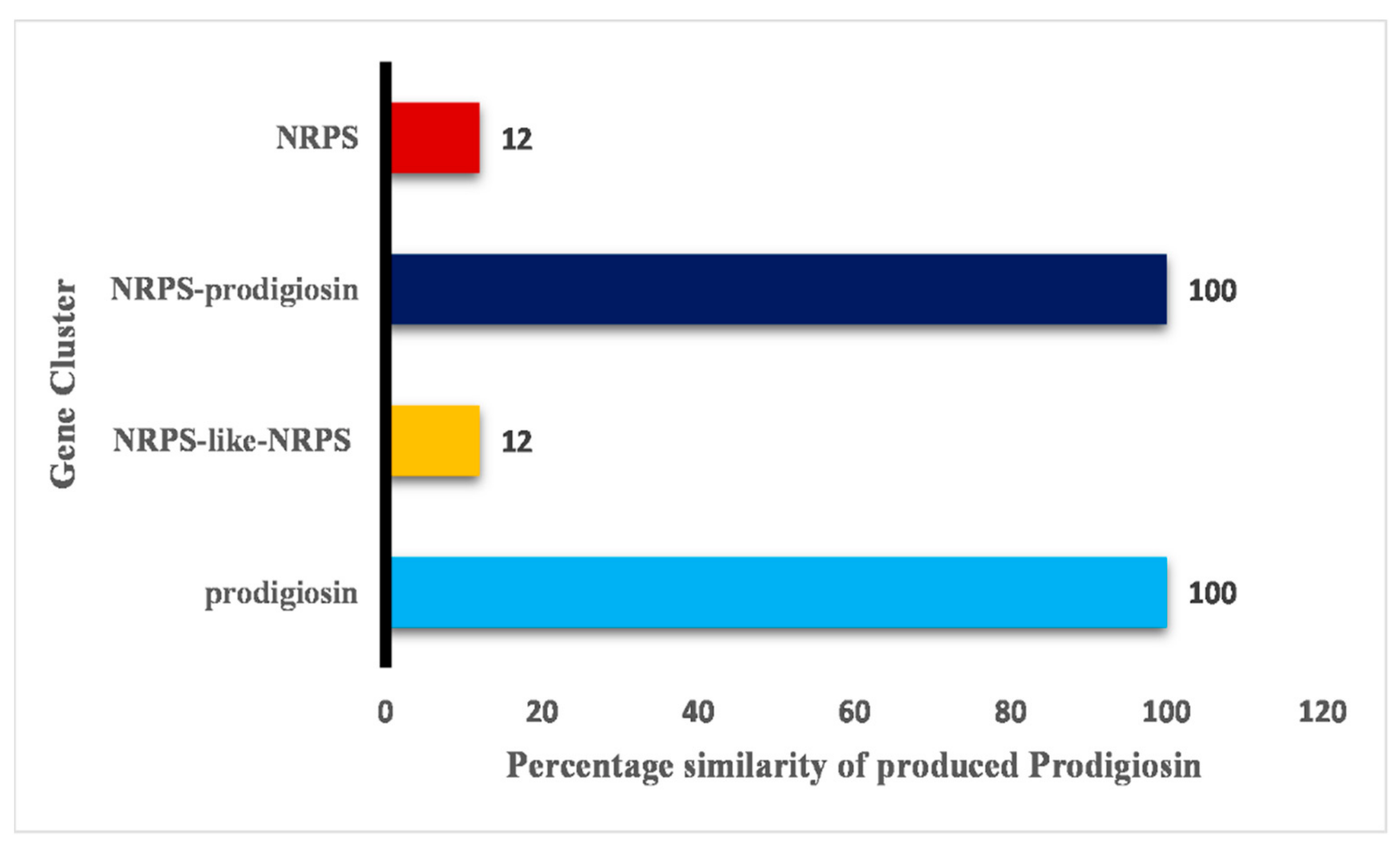
2.6. Biosynthetic Gene Cluster Prediction Report Analysis
3. Discussion
4. Experimental Section
4.1. Isolation and Purification of Soil Bacteria
4.2. Molecular Characterization
4.2.1. Isolation of JSSCPM1 Strain Genomic DNA
4.2.2. 16S rRNA Gene Sequencing of JSSCPM1 Strain
4.2.3. Phylogenetic Tree Construction
4.3. Pigment Extraction and Characterization
4.3.1. UV-Visible Spectroscopic Analysis
4.3.2. HPLC Analysis
4.3.3. LC-MS Analysis
4.3.4. 1H and 13C NMR Spectroscopic Analysis
4.4. Molecular Docking Process—Experimental Protocol
4.4.1. Preparation of Protein Targets
4.4.2. Preparation of Ligand
4.4.3. Molecular Docking
4.4.4. Protein–Ligand Interactions
4.5. Antimicrobial Activity
4.6. Biosynthetic Gene Cluster Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colson, A.R.; Morton, A.; Årdal, C.; Chalkidou, K.; Davies, S.C.; Garrison, L.P.; Jit, M.; Laxminarayan, R.; Megiddo, I.; Morel, C. Antimicrobial Resistance: Is Health Technology Assessment Part of the Solution or Part of the Problem? Value Health 2021, 24, 1828–1834. [Google Scholar] [CrossRef] [PubMed]
- Catalano, A.; Iacopetta, D.; Ceramella, J.; Scumaci, D.; Giuzio, F.; Saturnino, C.; Aquaro, S.; Rosano, C.; Sinicropi, M.S. Multidrug Resistance (MDR): A Widespread Phenomenon in Pharmacological Therapies. Molecules 2022, 27, 616. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.E.; Black, S.; Salisbury, D.; Rappuoli, R. Antimicrobial Resistance and the Role of Vaccines. Proc. Natl. Acad. Sci. USA 2018, 115, 12868–12871. [Google Scholar] [CrossRef] [PubMed]
- Parmanik, A.; Das, S.; Kar, B.; Bose, A.; Dwivedi, G.R.; Pandey, M.M. Current Treatment Strategies against Multidrug-Resistant Bacteria: A Review. Curr. Microbiol. 2022, 79, 388. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, S.E.; Fletcher, M.H.; Wuest, W.M. Natural Products as Platforms to Overcome Antibiotic Resistance. Chem. Rev. 2017, 117, 12415–12474. [Google Scholar] [CrossRef] [PubMed]
- Herráez, R.; Mur, A.; Merlos, A.; Viñas, M.; Vinuesa, T. Using Prodigiosin against Some Gram-Positive and Gram-Negative Bacteria and Trypanosoma cruzi. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e20190001. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, N.; Senerovic, L.; Ilic-Tomic, T.; Vasiljevic, B.; Nikodinovic-Runic, J. Properties and Applications of Undecylprodigiosin and Other Bacterial Prodigiosins. Appl. Microbiol. Biotechnol. 2014, 98, 3841–3858. [Google Scholar] [CrossRef]
- Kamble, K.D.; Hiwarale, V.D. Prodigiosin Production from Serratia marcescens Strains Obtained from Farm Soil. Int. J. Environ. Sci. 2012, 3, 631–638. [Google Scholar]
- Metwally, R.A.; El Sikaily, A.; El-Sersy, N.A.; Ghozlan, H.A.; Sabry, S.A. Antimicrobial Activity of Textile Fabrics Dyed with Prodigiosin Pigment Extracted from Marine Serratia Rubidaea RAM_Alex Bacteria. Egypt. J. Aquat. Res. 2021, 47, 301–305. [Google Scholar] [CrossRef]
- Othman, M.A.; El-Zamik, F.I.; Hegazy, M.I.; Salama, A.S.A. Isolation and Identification of Egyptian Strains of Serratia marcescens Producing Antibacterial and Antioxidant Prodigiosin Pigment. Zagazig J. Agric. Res. 2019, 46, 1573–1582. [Google Scholar] [CrossRef]
- Erandapurathukadumana Sreedharan, H.; Harilal, C.C.; Pradeep, S. Response Surface Optimization of Prodigiosin Production by Phthalate Degrading Achromobacter Denitrificans SP1 and Exploring Its Antibacterial Activity. Prep. Biochem. Biotechnol. 2020, 50, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Hage-Hülsmann, J.; Grünberger, A.; Thies, S.; Santiago-Schübel, B.; Klein, A.S.; Pietruszka, J.; Binder, D.; Hilgers, F.; Domröse, A.; Drepper, T. Natural Biocide Cocktails: Combinatorial Antibiotic Effects of Prodigiosin and Biosurfactants. PLoS ONE 2018, 13, e0200940. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.-H.; Mahalingam, S.; Wan, K.-L.; Nathan, S. Prodigiosin Inhibits Bacterial Growth and Virulence Factors as a Potential Physiological Response to Interspecies Competition. PLoS ONE 2021, 16, e0253445. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Gong, J.; Fu, R.; Zhang, J.; Fang, K.; Liu, X. Antibacterial Dyeing of Silk with Prodigiosins Suspention Produced by Liquid Fermentation. J. Clean. Prod. 2018, 201, 648–656. [Google Scholar] [CrossRef]
- Suryawanshi, R.K.; Patil, C.D.; Koli, S.H.; Hallsworth, J.E.; Patil, S.V. Antimicrobial Activity of Prodigiosin Is Attributable to Plasma-Membrane Damage. Nat. Prod. Res. 2017, 31, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Jimtha, C.J.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E.K. Antifungal Properties of Prodigiosin Producing Rhizospheric Serratia Sp. Rhizosphere 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Elahian, F.; Moghimi, B.; Dinmohammadi, F.; Ghamghami, M.; Hamidi, M.; Mirzaei, S.A. The Anticancer Agent Prodigiosin Is Not a Multidrug Resistance Protein Substrate. DNA Cell Biol. 2013, 32, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Tomás, R.P.; Ruir, C.D.; Montaner, B. Prodigiosin Induces Cell Death and Morphological Changes Indicative of Apoptosis in Gastric Cancer Cell Line HGT-1. Histol. Histopathol. 2001, 16, 415–421. [Google Scholar]
- Cheng, S.-Y.; Chen, N.-F.; Kuo, H.-M.; Yang, S.-N.; Sung, C.-S.; Sung, P.-J.; Wen, Z.-H.; Chen, W.-F. Prodigiosin Stimulates Endoplasmic Reticulum Stress and Induces Autophagic Cell Death in Glioblastoma Cells. Apoptosis 2018, 23, 314–328. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Chen, S.-P.; Nguyen, T.H.; Nguyen, M.T.; Tran, T.T.T.; Doan, C.T.; Tran, T.N.; Nguyen, A.D.; Kuo, Y.-H.; Wang, S.-L. Novel Efficient Bioprocessing of Marine Chitins into Active Anticancer Prodigiosin. Mar. Drugs 2019, 18, 15. [Google Scholar] [CrossRef]
- Akpan, U.M.; Pellegrini, M.; Obayemi, J.D.; Ezenwafor, T.; Browl, D.; Ani, C.J.; Yiporo, D.; Salifu, A.; Dozie-Nwachukwu, S.; Odusanya, S. Prodigiosin-Loaded Electrospun Nanofibers Scaffold for Localized Treatment of Triple Negative Breast Cancer. Mater. Sci. Eng. C 2020, 114, 110976. [Google Scholar] [CrossRef] [PubMed]
- Patil, C.D.; Suryawanshi, R.K.; Koujah, L.; Shukla, D. Antiviral Efficacy of Prodigiosin against Corneal Herpes Simplex Virus Infection. Investig. Ophthalmol. Vis. Sci. 2020, 61, 2982. [Google Scholar]
- Zhou, W.; Zeng, C.; Liu, R.; Chen, J.; Li, R.; Wang, X.; Bai, W.; Liu, X.; Xiang, T.; Zhang, L. Antiviral Activity and Specific Modes of Action of Bacterial Prodigiosin against Bombyx mori Nucleopolyhedrovirus in Vitro. Appl. Microbiol. Biotechnol. 2016, 100, 3979–3988. [Google Scholar] [CrossRef] [PubMed]
- Darshan, N.; Manonmani, H.K. Prodigiosin and Its Potential Applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed]
- Papireddy, K.; Smilkstein, M.; Kelly, J.X.; Shweta; Salem, S.M.; Alhamadsheh, M.; Haynes, S.W.; Challis, G.L.; Reynolds, K.A. Antimalarial Activity of Natural and Synthetic Prodiginines. J. Med. Chem. 2011, 54, 5296–5306. [Google Scholar] [CrossRef] [PubMed]
- Garcha, S.; Verma, N.; Brar, S.K. Isolation, Characterization and Identification of Microorganisms from Unorganized Dairy Sector Wastewater and Sludge Samples and Evaluation of Their Biodegradability. Water Resour. Ind. 2016, 16, 19–28. [Google Scholar] [CrossRef]
- De Clerck, E.; Vanhoutte, T.; Hebb, T.; Geerinck, J.; Devos, J.; De Vos, P. Isolation, Characterization, and Identification of Bacterial Contaminants in Semifinal Gelatin Extracts. Appl. Environ. Microbiol. 2004, 70, 3664–3672. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.K.P.; Williamson, N.R.; Slater, H.; Cox, A.; Abbasi, S.; Foulds, I.; Simonsen, H.T.; Leeper, F.J.; Salmond, G.P.C. The Serratia Gene Cluster Encoding Biosynthesis of the Red Antibiotic, Prodigiosin, Shows Species-and Strain-Dependent Genome Context Variation. Microbiology 2004, 150, 3547–3560. [Google Scholar] [CrossRef]
- Yip, C.-H.; Yarkoni, O.; Ajioka, J.; Wan, K.-L.; Nathan, S. Recent Advancements in High-Level Synthesis of the Promising Clinical Drug, Prodigiosin. Appl. Microbiol. Biotechnol. 2019, 103, 1667–1680. [Google Scholar] [CrossRef]
- Panesar, R.; Kaur, S.; Panesar, P.S. Production of Microbial Pigments Utilizing Agro-Industrial Waste: A Review. Curr. Opin. Food Sci. 2015, 1, 70–76. [Google Scholar] [CrossRef]
- Edelmann, M.; Aalto, S.; Chamlagain, B.; Kariluoto, S.; Piironen, V. Riboflavin, Niacin, Folate and Vitamin B12 in Commercial Microalgae Powders. J. Food Compos. Anal. 2019, 82, 103226. [Google Scholar] [CrossRef]
- Bogacz-Radomska, L.; Harasym, J. β-Carotene—Properties and Production Methods. Food Qual. Saf. 2018, 2, 69–74. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Watanabe, B.; Matsui, K.; Lumyong, S. Characterization of Melanin and Optimal Conditions for Pigment Production by an Endophytic Fungus, Spissiomyces Endophytica SDBR-CMU319. PLoS ONE 2019, 14, e0222187. [Google Scholar] [CrossRef] [PubMed]
- Fang, N.; Wang, C.; Liu, X.; Zhao, X.; Liu, Y.; Liu, X.; Du, Y.; Zhang, Z.; Zhang, H. De Novo Synthesis of Astaxanthin: From Organisms to Genes. Trends Food Sci. Technol. 2019, 92, 162–171. [Google Scholar] [CrossRef]
- Dasgupta Mandal, D.; Majumdar, S.; Dey, S.; Dutta, S.; Mandal, T. Utilization of Low-Cost Fatty Acid Sources by Bacterial Isolate for Improved Production of Valuable Prodigiosin. In Advances in Bioprocess Engineering and Technology: Select Proceedings ICABET 2020; Springer: Singapore, 2021; pp. 21–27. [Google Scholar]
- De Araújo, H.W.C.; Fukushima, K.; Takaki, G.M.C. Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate. Molecules 2010, 15, 6931–6940. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Román, M.I.; Holguín-Meléndez, F.; Bello-Mendoza, R.; Guillén-Navarro, K.; Dunn, M.F.; Huerta-Palacios, G. Production of Prodigiosin and Chitinases by Tropical Serratia marcescens Strains with Potential to Control Plant Pathogens. World J. Microbiol. Biotechnol. 2012, 28, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Jia, X.; Fang, Y.; Chen, L.; Zhang, H.; Lin, R.; Chen, J. Enhanced Production of Prodigiosin by Serratia marcescens FZSF02 in the Form of Pigment Pellets. Electron. J. Biotechnol. 2019, 40, 58–64. [Google Scholar] [CrossRef]
- Naik, C.; Srisevita, J.M.; Shushma, K.N.; Noorin, F.; Shilpa, A.C.; Muttanna, C.D.; Darshan, N.; Sannadurgappa, D. Peanut Oil Cake: A Novel Substrate for Enhanced Cell Growth and Prodigiosin Production from Serratia marcescens CF-53. J. Res. Biol. 2012, 2, 549–557. [Google Scholar]
- Darshan, N.; Manonmani, H.K. Prodigiosin Inhibits Motility and Activates Bacterial Cell Death Revealing Molecular Biomarkers of Programmed Cell Death. Amb Express 2016, 6, 50. [Google Scholar] [CrossRef]
- Manas, N.H.A.; Chong, L.Y.; Tesfamariam, Y.M.; Zulkharnain, A.; Mahmud, H.; Mahmod, D.S.A.; Fuzi, S.F.Z.M.; Azelee, N.I.W. Effects of Oil Substrate Supplementation on Production of Prodigiosin by Serratia nematodiphila for Dye-Sensitized Solar Cell. J. Biotechnol. 2020, 317, 16–26. [Google Scholar] [CrossRef]
- Vitale, G.A.; Sciarretta, M.; Palma Esposito, F.; January, G.G.; Giaccio, M.; Bunk, B.; Spröer, C.; Bajerski, F.; Power, D.; Festa, C. Genomics–Metabolomics Profiling Disclosed Marine Vibrio Spartinae 3.6 as a Producer of a New Branched Side Chain Prodigiosin. J. Nat. Prod. 2020, 83, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Alihosseini, F.; Ju, K.; Lango, J.; Hammock, B.D.; Sun, G. Antibacterial Colorants: Characterization of Prodiginines and Their Applications on Textile Materials. Biotechnol. Prog. 2008, 24, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Montaner, B.; Navarro, S.; Piqué, M.; Vilaseca, M.; Martinell, M.; Giralt, E.; Gil, J.; Pérez-Tomás, R. Prodigiosin from the Supernatant of Serratia marcescens Induces Apoptosis in Haematopoietic Cancer Cell Lines. Br. J. Pharmacol. 2000, 131, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Gulani, C.; Bhattacharya, S.; Das, A. Assessment of Process Parameters Influencing the Enhanced Production of Prodigiosin from Serratia marcescens and Evaluation of Its Antimicrobial, Antioxidant and Dyeing Potentials. Malays. J. Microbiol. 2012, 8, 116–122. [Google Scholar]
- Pore, T.S.; Khanolkar, A.B.; Nadaf, N.H. Production, Purification, Identification of Prodigiosin from Serratia Sp. and Its Antimicrobial Activity. Res. J. Life Sci. Bioinform. Pharm. Chem. Sci. 2016, 1, 326. [Google Scholar]
- Lapenda, J.C.L.; Alves, V.P.; Adam, M.L.; Rodrigues, M.D.; Nascimento, S.C. Cytotoxic Effect of Prodigiosin, Natural Red Pigment, Isolated from Serratia marcescens UFPEDA 398. Indian J. Microbiol. 2020, 60, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Lapenda, J.C.; Silva, P.A.; Vicalvi, M.C.; Sena, K.; Nascimento, S.C. Antimicrobial Activity of Prodigiosin Isolated from Serratia marcescens UFPEDA 398. World J. Microbiol. Biotechnol. 2015, 31, 399–406. [Google Scholar] [CrossRef]
- Kimyon, Ö.; Das, T.; Ibugo, A.I.; Kutty, S.K.; Ho, K.K.; Tebben, J.; Kumar, N.; Manefield, M. Serratia Secondary Metabolite Prodigiosin Inhibits Pseudomonas Aeruginosa Biofilm Development by Producing Reactive Oxygen Species That Damage Biological Molecules. Front. Microbiol. 2016, 7, 972. [Google Scholar] [CrossRef]
- Tao, J.; Wang, X.; Shen, Y.; Wei, D. Strategy for the Improvement of Prodigiosin Production by a Serratia marcescens Mutant through Fed-Batch Fermentation. World J. Microbiol. Biotechnol. 2005, 21, 969–972. [Google Scholar] [CrossRef]
- Merlino, C.P. Bartolomeo Bizio’s Letter to the Most Eminent Priest, Angelo Bellani, Concerning the Phenomenon of the Red-Colored Polenta. J. Bacteriol. 1924, 9, 527–543. [Google Scholar] [CrossRef]
- Setiyono, E.; Adhiwibawa, M.A.S.; Indrawati, R.; Prihastyanti, M.N.U.; Shioi, Y.; Brotosudarmo, T.H.P. An Indonesian Marine Bacterium, Pseudoalteromonas Rubra, Produces Antimicrobial Prodiginine Pigments. ACS Omega 2020, 5, 4626–4635. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.L.T.; Nguyen, T.C.; Do, T.T.; Vu, T.L.; Nguyen, T.T.; Do, T.T.; Nguyen, T.H.T.; Le, T.H.; Trinh, D.K.; Nguyen, T.A.T. Study on the Anticancer Activity of Prodigiosin from Variants of Serratia marcescens QBN VTCC 910026. BioMed Res. Int. 2022, 2022, 4053074. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, Y.-S.; Park, S.; Kim, J.; Kang, S.-J.; Lee, M.-H.; Ryu, S.; Choi, J.M.; Oh, T.-K.; Yoon, J.-H. Exceptional Production of Both Prodigiosin and Cycloprodigiosin as Major Metabolic Constituents by a Novel Marine Bacterium, Zooshikella Rubidus S1-1. Appl. Environ. Microbiol. 2011, 77, 4967–4973. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, J.; Wang, X.; Kong, D.; Du, W.; Li, H.; Hse, C.-Y.; Shupe, T.; Zhou, D.; Zhao, K. Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 3465. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-J.; Bae, J.; Lee, D.-S.; Kim, C.-H.; Kim, J.-S.; Kim, S.-W.; Hong, S.-I. Purification and Characterization of Prodigiosin Produced by Integrated Bioreactor from Serratia Sp. KH-95. J. Biosci. Bioeng. 2006, 101, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Pagès, J.M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef]
- Ibrahim, D.; Nazari, T.F.; Kassim, J.; Lim, S.-H. Prodigiosin—An Antibacterial Red Pigment Produced by Serratia marcescens IBRL USM 84 Associated with a Marine Sponge Xestospongia Testudinaria. J. Appl. Pharm. Sci. 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Alqahtani, S.S.; Moni, S.S.; Sultan, M.H.; Bakkari, M.A.; Madkhali, O.A.; Alshahrani, S.; Makeen, H.A.; Menachery, S.J.; ur Rehman, Z.; Alam, M.S. Potential Bioactive Secondary Metabolites of Actinomycetes Sp. Isolated from Rocky Soils of the Heritage Village Rijal Alma, Saudi Arabia. Arab. J. Chem. 2022, 15, 103793. [Google Scholar] [CrossRef]
- Chakraborty, B.; Kumar, R.S.; Almansour, A.I.; Gunasekaran, P.; Nayaka, S. Bioprospection and Secondary Metabolites Profiling of Marine Streptomyces Levis Strain KS46. Saudi J. Biol. Sci. 2022, 29, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Njenga, P.; Wagacha, J.M.; Gathuru, E.M. Methods of Isolating Actinomycetes from the Soils of Menengai Crater in Kenya. Arch. Clin. Microbiol. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Nayaka, S.; Chakraborty, B.; Bhat, M.P.; Nagaraja, S.K.; Airodagi, D.; Swamy, P.S.; Rudrappa, M.; Hiremath, H.; Basavarajappa, D.S.; Kanakannanavar, B. Biosynthesis, Characterization, and in Vitro Assessment on Cytotoxicity of Actinomycete-Synthesized Silver Nanoparticles on Allium cepa Root Tip Cells. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 51. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, S.; Kumar, A. Extraction and Characterization of Secondary Metabolites Produced by Bacteria Isolated from Industrial Wastewater. J. Water Process Eng. 2021, 40, 101811. [Google Scholar] [CrossRef]
- Jeong, S.W.; Yang, J.E.; Choi, Y.J. Isolation and Characterization of a Yellow Xanthophyll Pigment-Producing Marine Bacterium, Erythrobacter Sp. SDW2 Strain, in Coastal Seawater. Mar. Drugs 2022, 20, 73. [Google Scholar] [CrossRef] [PubMed]
- Wood, W.A.; Krieg, N.R. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1989; pp. 1–9. [Google Scholar]
- Lewis, S.M.; Corpe, W.A. Prodigiosin-Producing Bacteria from Marine Sources. Appl. Microbiol. 1964, 12, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Lee, C.-C.; Lin, Y.-L.; Yin, K.-M.; Ho, C.-L.; Liu, T. Obtaining Long 16S RDNA Sequences Using Multiple Primers and Its Application on Dioxin-Containing Samples. BMC Bioinform. 2015, 16, S13. [Google Scholar] [CrossRef]
- Fahmy, N.M.; Abdel-Tawab, A.M. Isolation and Characterization of Marine Sponge–Associated Streptomyces Sp. NMF6 Strain Producing Secondary Metabolite(s) Possessing Antimicrobial, Antioxidant, Anticancer, and Antiviral Activities. J. Genet. Eng. Biotechnol. 2021, 19, 102. [Google Scholar] [CrossRef]
- Almalki, M.A. Isolation and Characterization of Polyketide Drug Molecule from Streptomyces Species with Antimicrobial Activity against Clinical Pathogens. J. Infect. Public Health 2020, 13, 125–130. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zerrifi, S.E.A.; Redouane, E.M.; Mugani, R.; Ribeiro, I.; de Fátima Carvalho, M.; Campos, A.; Barakate, M.; Vasconcelos, V.; Oudra, B.; El Khalloufi, F. Moroccan Actinobacteria with Promising Activity against Toxic Cyanobacteria Microcystis Aeruginosa. Environ. Sci. Pollut. Res. 2021, 28, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Garzón, J.F.; Zehl, M.; Schneider, O.; Rückert, C.; Busche, T.; Kalinowski, J.; Bredholt, H.; Zotchev, S.B. Streptomyces Spp. from the Marine Sponge Antho Dichotoma: Analyses of Secondary Metabolite Biosynthesis Gene Clusters and Some of Their Products. Front. Microbiol. 2020, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Shen, J.; Ou, P.; Liu, L.; Chen, Z.; Chu, F.; Wang, J.; Jin, X. Prodigiosin Isolated from Serratia marcescens in the Periplaneta Americana Gut and Its Apoptosis-inducing Activity in HeLa Cells. Oncol. Rep. 2019, 41, 3377–3385. [Google Scholar] [CrossRef] [PubMed]
- Koyun, M.T.; Sirin, S.; Aslim, B.; Taner, G.; Dolanbay, S.N. Characterization of Prodigiosin Pigment by Serratia marcescens and the Evaluation of Its Bioactivities. Toxicol. In Vitro 2022, 82, 105368. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Isbrandt, T.; Christensen, E.Ø.; Melchiorsen, J.; Larsen, T.O.; Zhang, S.-D.; Gram, L. Identification and Verification of the Prodigiosin Biosynthetic Gene Cluster (BGC) in Pseudoalteromonas Rubra S4059. Microbiol. Spectr. 2021, 9, e01171-21. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-A.; Liu, Y. Sequencing and Characterization of a Novel Serine Metalloprotease from Burkholderia pseudomallei. FEMS Microbiol. Lett. 2000, 192, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Aruldass, C.A.; Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Brown Sugar as a Low-Cost Medium for the Production of Prodigiosin by Locally Isolated Serratia marcescens UTM1. Int. Biodeterior. Biodegrad. 2014, 95, 19–24. [Google Scholar] [CrossRef]
- Nastasă, C.; Tamaian, R.; Oniga, O.; Tiperciuc, B. 5-Arylidene (Chromenyl-Methylene)-Thiazolidinediones: Potential New Agents against Mutant Oncoproteins K-Ras, N-Ras and B-Raf in Colorectal Cancer and Melanoma. Medicina 2019, 55, 85. [Google Scholar] [CrossRef]
- Jabir, N.R.; Rehman, M.T.; Alsolami, K.; Shakil, S.; Zughaibi, T.A.; Alserihi, R.F.; Khan, M.S.; AlAjmi, M.F.; Tabrez, S. Concatenation of Molecular Docking and Molecular Simulation of BACE-1, γ-Secretase Targeted Ligands: In Pursuit of Alzheimer’s Treatment. Ann. Med. 2021, 53, 2332–2344. [Google Scholar] [CrossRef]
- Nishimura, Y.; Hara, H. Drug Repositioning: Current Advances and Future Perspectives. Front. Pharmacol. 2018, 9, 1068. [Google Scholar] [CrossRef] [PubMed]
- Ziervogel, B.K.; Roux, B. The Binding of Antibiotics in OmpF Porin. Structure 2013, 21, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated Comparative Protein Structure Modeling with SWISS-MODEL and Swiss-PdbViewer: A Historical Perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef] [PubMed]
- Bhimaneni, S.P.; Bhati, V.; Bhosale, S.; Kumar, A. Investigates Interaction between Abscisic Acid and Bovine Serum Albumin Using Various Spectroscopic and In-Silico Techniques. J. Mol. Struct. 2021, 1224, 129018. [Google Scholar] [CrossRef]
- Kant, K.; Rawat, R.; Bhati, V.; Bhosale, S.; Sharma, D.; Banerjee, S.; Kumar, A. Computational Identification of Natural Product Leads That Inhibit Mast Cell Chymase: An Exclusive Plausible Treatment for Japanese Encephalitis. J. Biomol. Struct. Dyn. 2021, 39, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Singh, A.; Sharma, R.; Kumar, A. Repurposing of Auranofin against Bacterial Infections: An in Silico and in Vitro Study. Curr. Comput. Aided. Drug Des. 2021, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Shaker, B.; Yu, M.-S.; Lee, J.; Lee, Y.; Jung, C.; Na, D. User Guide for the Discovery of Potential Drugs via Protein Structure Prediction and Ligand Docking Simulation. J. Microbiol. 2020, 58, 235–244. [Google Scholar] [CrossRef]
- Verma, E.; Mishra, A.K.; Singh, A.K.; Singh, V.K. Structural Elucidation and Molecular Docking of a Novel Antibiotic Compound from Cyanobacterium Nostoc Sp. MGL001. Front. Microbiol. 2016, 7, 1899. [Google Scholar]
- Gurram, S.R.; Azam, M.A. Design, Synthesis and Biological Evaluation of Some Novel N’-(1, 3-Benzothiazol-2-Yl)-Arylamide Derivatives as Antibacterial Agents. Chem. Pap. 2021, 75, 5435–5452. [Google Scholar] [CrossRef]
- Javid, S.; Ather, H.; Hani, U.; Siddiqua, A.; Asif Ansari, S.M.; Shanmugarajan, D.; Yogish Kumar, H.; Arivuselvam, R.; Purohit, M.N.; Kumar, B.P. Discovery of Novel Myristic Acid Derivatives as N-Myristoyltransferase Inhibitors: Design, Synthesis, Analysis, Computational Studies and Antifungal Activity. Antibiotics 2023, 12, 1167. [Google Scholar] [CrossRef]
- Levison, M.E. Pharmacodynamics of Antimicrobial Drugs. Infect. Dis. Clin. 2004, 18, 451–465. [Google Scholar] [CrossRef]
- Kumar, P.; Chauhan, A.; Kumar, M.; Kuanr, B.K.; Solanki, R.; Kapur, M.K. Draft Genome and Secondary Metabolite Biosynthetic Gene Clusters of Streptomyces Sp. Strain 196. Mol. Biol. Rep. 2020, 47, 6741–6747. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 49, D10. [Google Scholar] [CrossRef]
- Majer, H.M.; Ehrlich, R.L.; Ahmed, A.; Earl, J.P.; Ehrlich, G.D.; Beld, J. Whole Genome Sequencing of Streptomyces actuosus ISP-5337, Streptomyces sioyaensis B-5408, and Actinospica acidiphila B-2296 Reveals Secondary Metabolomes with Antibiotic Potential. Biotechnol. Rep. 2021, 29, e00596. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Protein PDB ID | Conventional Hydrogen Bond | Carbon–Hydrogen Bond | Pi-Cation | Pi-Sigma | Pi-Pi T-Shaped | PI-ANION | Alkyl | Pi-Alkyl |
|---|---|---|---|---|---|---|---|---|---|
| PG | 4CGS | ASN141, ASP107, SER177 | SER142, ASN152 | ARG140 | - | - | ARG140 | - | TYR102, TYR106 |
| 4CGP | lEU291 | GLY327, ASP121 | - | VAL292 | TYR32 | - | VAL326 | Leu324, ILE318, ILE240, lYS243 | |
| 4CGQ | ASP107 | GLU181 | ARG140 | - | TYR106 | ARG140 | - | PHE85, TYR102, ALA154 |
| Compound | Minimum Inhibitory Concentration of PG Compound (μg/mL) | ||||
|---|---|---|---|---|---|
| E. coli NCIM 2065 | K. pneumoniae NCIM 2706 | P. aeruginosa NCIM 2036 | B. subtilis NCIM 2545 | MRSA ATCC 43300 | |
| PG | 15.9 ± 0.31 | 22.6 ± 0.08 | 46.1 ± 0.5 | 43 ± 0.49 | 73.6 ± 0.16 |
| Ampicillin | 5.6 ± 0.72 | 3.2 ± 0.60 | 4 ± 0.26 | 3 ± 0.53 | 19.8 ± 0.32 |
| Carbenicillin | 4.1 ± 0.58 | 1.6 ± 0.77 | 3 ± 0.68 | 7 ± 0.08 | 13.1 ± 0.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arivuselvam, R.; Dera, A.A.; Parween Ali, S.; Alraey, Y.; Saif, A.; Hani, U.; Arumugam Ramakrishnan, S.; Azeeze, M.S.T.A.; Rajeshkumar, R.; Susil, A.; et al. Isolation, Identification, and Antibacterial Properties of Prodigiosin, a Bioactive Product Produced by a New Serratia marcescens JSSCPM1 Strain: Exploring the Biosynthetic Gene Clusters of Serratia Species for Biological Applications. Antibiotics 2023, 12, 1466. https://doi.org/10.3390/antibiotics12091466
Arivuselvam R, Dera AA, Parween Ali S, Alraey Y, Saif A, Hani U, Arumugam Ramakrishnan S, Azeeze MSTA, Rajeshkumar R, Susil A, et al. Isolation, Identification, and Antibacterial Properties of Prodigiosin, a Bioactive Product Produced by a New Serratia marcescens JSSCPM1 Strain: Exploring the Biosynthetic Gene Clusters of Serratia Species for Biological Applications. Antibiotics. 2023; 12(9):1466. https://doi.org/10.3390/antibiotics12091466
Chicago/Turabian StyleArivuselvam, Rajaguru, Ayed A. Dera, Syed Parween Ali, Yasser Alraey, Ahmed Saif, Umme Hani, Sivaa Arumugam Ramakrishnan, Mohamed Sheik Tharik Abdul Azeeze, Raman Rajeshkumar, Aishwarya Susil, and et al. 2023. "Isolation, Identification, and Antibacterial Properties of Prodigiosin, a Bioactive Product Produced by a New Serratia marcescens JSSCPM1 Strain: Exploring the Biosynthetic Gene Clusters of Serratia Species for Biological Applications" Antibiotics 12, no. 9: 1466. https://doi.org/10.3390/antibiotics12091466
APA StyleArivuselvam, R., Dera, A. A., Parween Ali, S., Alraey, Y., Saif, A., Hani, U., Arumugam Ramakrishnan, S., Azeeze, M. S. T. A., Rajeshkumar, R., Susil, A., Harindranath, H., & Kumar, B. R. P. (2023). Isolation, Identification, and Antibacterial Properties of Prodigiosin, a Bioactive Product Produced by a New Serratia marcescens JSSCPM1 Strain: Exploring the Biosynthetic Gene Clusters of Serratia Species for Biological Applications. Antibiotics, 12(9), 1466. https://doi.org/10.3390/antibiotics12091466