
Sub-MIC Antibiotics Modulate Productions of Outer Membrane Vesicles in Tigecycline-Resistant Escherichia coli

 and
and
Abstract
:1. Introduction
2. Results
2.1. AMR Phenotypes of E. coli 47EC
2.2. Effects of Antibiotics on the Production of OMVs from E. coli 47EC
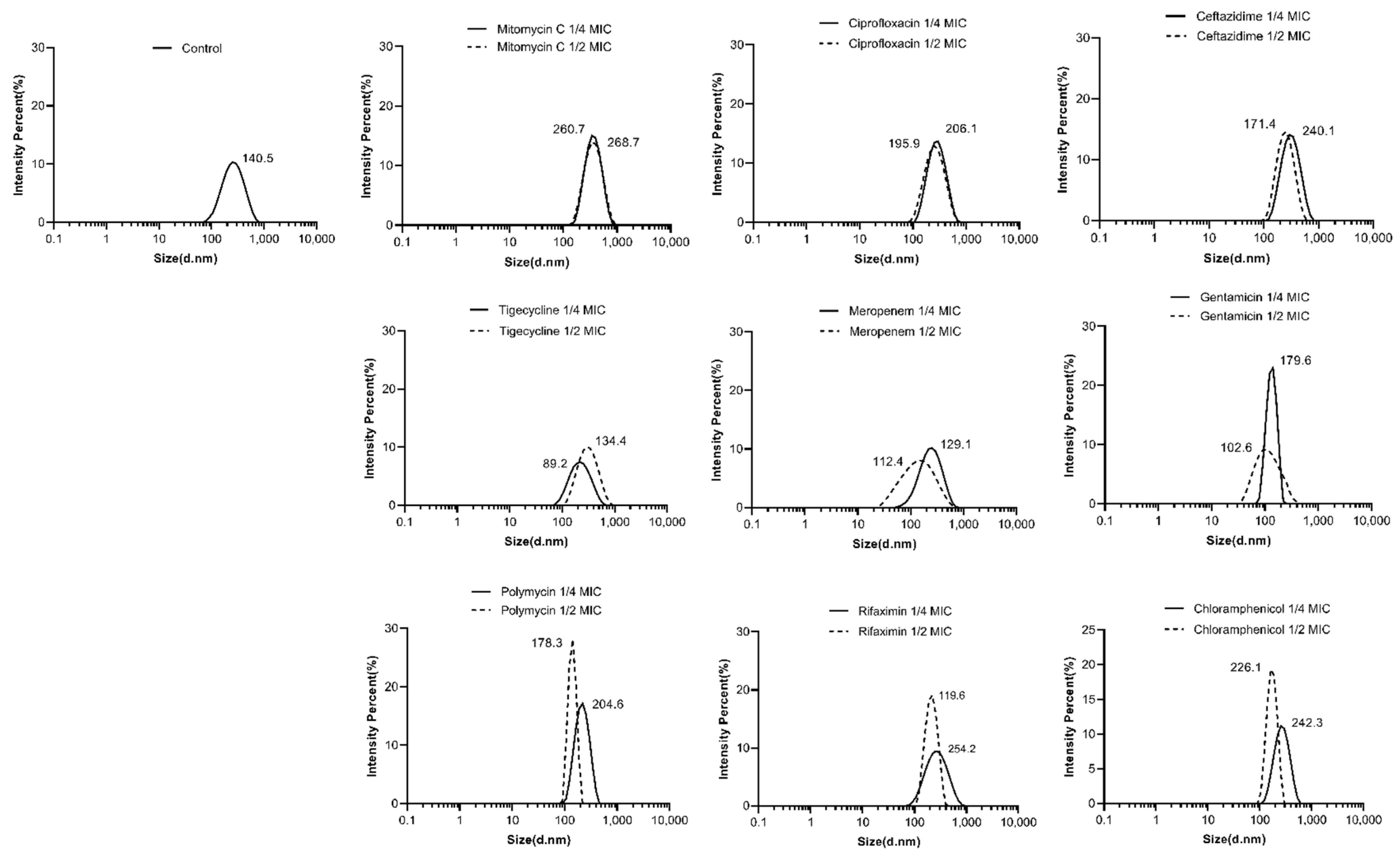
2.3. Effects of Antibiotics on the Characterizations of OMVs from E. coli 47EC
2.4. Abundance of tet(X4) in OMVs
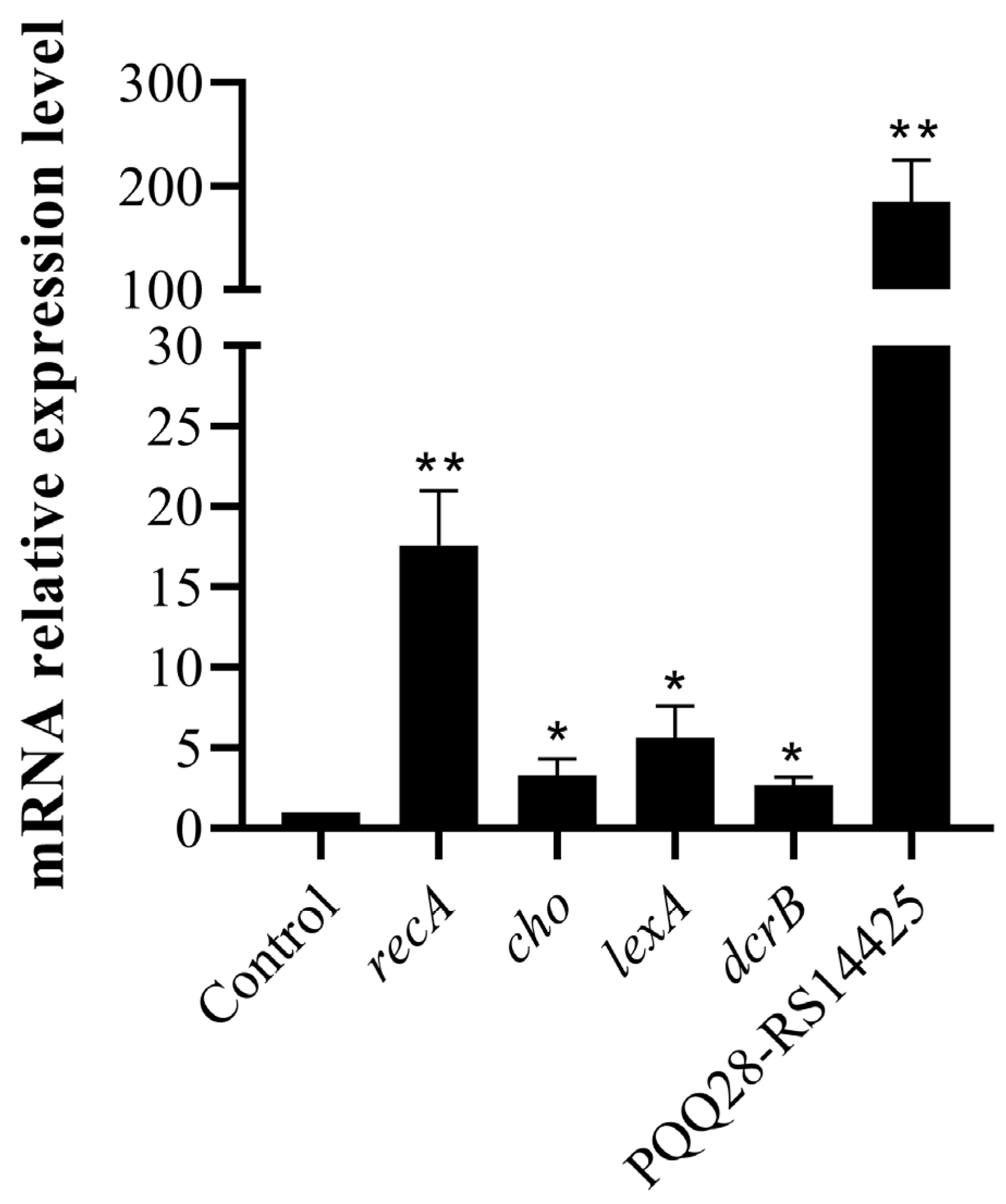
2.5. Transcriptome Changes in E. coli 47EC Induced by Ciprofloxacin
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strain
5.2. Antimicrobial Susceptibility Testing
5.3. Isolation of OMV
5.4. Quantification of OMVs
5.5. Size Distribution and Zeta Potential of OMVs
5.6. Detection of tet(X4) in OMVs
5.7. Morphology of OMVs
5.8. Transcriptome Analysis of E. coli 47EC under Ciprofloxacin Treatment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Djordjevic, S.P.; Jarocki, V.M.; Seemann, T.; Cummins, M.L.; Watt, A.E.; Drigo, B.; Wyrsch, E.R.; Reid, C.J.; Donner, E.; Howden, B.P. Genomic surveillance for antimicrobial resistance—A One Health perspective. Nat. Rev. Genet. 2023, 25, 142–157. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.A.; Wang, Y.X.; Diarra, M.S.; Alexander, T.; Stanford, K. Challenges of a one-health approach to the development of alternatives to antibiotics. Anim. Front. 2018, 8, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Shuai, X.Y.; Lin, Z.J.; Sun, Y.J.; Zhou, Z.C.; Meng, L.X.; Zhu, Y.G.; Chen, H. Landscape of genes in hospital wastewater breaking through the defense line of last-resort antibiotics. Water Res. 2022, 209, 117907. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, S.; Zekiy, A.O.; Krutova, M.; Gholami, M.; Kouhsari, E.; Sholeh, M.; Ghafouri, Z.; Maleki, F. Tigecycline antibacterial activity, clinical effectiveness, and mechanisms and epidemiology of resistance: Narrative review. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1003–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wen, J.; Wang, Y.; Wang, M.; Jia, R.; Chen, S.; Liu, M.; Zhu, D.; Zhao, X.; Wu, Y.; et al. Dissemination and prevalence of plasmid-mediated high-level tigecycline resistance gene tet(X4). Front. Microbiol. 2022, 13, 969769. [Google Scholar] [CrossRef]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Sun, J.; Chen, C.; Cui, C.; Zhang, Y.; Liu, X.; Cui, Z.; Ma, X.; Feng, Y.; Fang, L.; Lian, X.; et al. Plasmid-encoded tet(X) genes that confer high-level tigecycline resistance in Escherichia coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef]
- Dell’Annunziata, F.; Folliero, V.; Giugliano, R.; De Filippis, A.; Santarcangelo, C.; Izzo, V.; Daglia, M.; Galdiero, M.; Arciola, C.R.; Franci, G. Gene transfer potential of outer membrane vesicles of Gram-Negative bacteria. Int. J. Mol. Sci. 2021, 22, 5985. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Sartorio, M.G.; Pardue, E.J.; Feldman, M.F.; Haurat, M.F. Bacterial outer membrane vesicles: From discovery to applications. Annu. Rev. Microbiol. 2021, 75, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, A.; De Wever, O. Systemically circulating bacterial extracellular vesicles: Origin, fate, and function. Trends Microbiol. 2022, 30, 213–216. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.D.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer membrane vesicles of gram-negative bacteria: An outlook on biogenesis. Front. Microbiol. 2021, 12, 557902. [Google Scholar] [CrossRef]
- Juodeikis, R.; Carding, S.R. Outer membrane vesicles: Biogenesis, functions, and issues. Microbiol. Mol. Biol. Rev. 2022, 86, e0003222. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, S.Y.; Son, J.H.; Kim, S.I.; Lee, H.; Kim, S.; Lee, H.; Kim, S.; Shin, M.; Lee, J.C. Production of membrane vesicles by cultured with or without subinhibitory concentrations of antibiotics and their pathological effects on epithelial cells. Front. Cell Infect. Microbiol. 2019, 9, 295. [Google Scholar] [CrossRef] [PubMed]
- Devos, S.; Van Oudenhove, L.; Stremersch, S.; Van Putte, W.; De Rycke, R.; Van Driessche, G.; Vitse, J.; Raemdonck, K.; Derveese, B. The effect of imipenem and diffusible signaling factors on the secretion of outer membrane vesicles and associated Ax21 proteins in Stenotrophomonas maltophilia. Front. Microbiol. 2015, 6, 298. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. M100, 28th ed.; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Fang, Y.; Yang, G.; Wu, X.; Qin, B.; Xie, Y.; Zhuang, L. Sub-MIC antibiotics affect microbial ferrihydrite reduction by extracellular membrane vesicles. J. Hazard. Mater. 2023, 458, 131876. [Google Scholar] [CrossRef]
- Cho, H.; Sondak, T.; Kim, K.S. Characterization of increased extracellular vesicle-mediated tigecycline resistance in Acinetobacter baumannii. Pharmaceutics 2023, 15, 1251. [Google Scholar] [CrossRef] [PubMed]
- Devos, S.; Van Putte, W.; Vitse, J.; Van Driessche, G.; Stremersch, S.; Van Den Broek, W.; Raemdonck, K.; Braeckmans, K.; Stahlberg, H.; Kudryashev, M.; et al. Membrane vesicle secretion and prophage induction in multidrug-resistant Stenotrophomonas maltophilia in response to ciprofloxacin stress. Environ. Microbiol. 2017, 19, 3930–3937. [Google Scholar] [CrossRef]
- Macdonald, I.A.; Kuehn, M.J. Stress-induced outer membrane vesicle production by Pseudomonas aeruginosa. J. Bacteriol. 2013, 195, 2971–2981. [Google Scholar] [CrossRef]
- Siqueira, V.L.; Cardoso, R.F.; Caleffi-Ferracioli, K.R.; Scodro, R.B.; Fernandez, M.A.; Fiorini, A.; Ueda-Nakamura, T.; Dias-Filho, B.P.; Nakamura, C.V. Structural changes and differentially expressed genes in Pseudomonas aeruginosa exposed to meropenem-ciprofloxacin combination. Antimicrob. Agents Chemother. 2014, 58, 3957–3967. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Clarke, A.J.; Beveridge, T.J. Surface action of gentamicin on Pseudomonas aeruginosa. J. Bacteriol. 1993, 175, 5798–5805. [Google Scholar] [CrossRef]
- Andreoni, F.; Toyofuku, M.; Menzi, C.; Kalawong, R.; Shambat, S.M.; Francois, P.; Zinkernagel, A.; Eberl, L. Antibiotics stimulate formation of vesicles in Staphylococcus aureus in both phage-dependent and -independent fashions and via different routes. Antimicrob. Agents Chemother. 2019, 63, e01439-18. [Google Scholar] [CrossRef]
- Bauwens, A.; Kunsmann, L.; Karch, H.; Mellmann, A.; Bielaszewska, M. Antibiotic-mediated modulations of outer membrane vesicles in Enterohemorrhagic Escherichia coli O104:H4 and O157:H7. Antimicrob. Agents Chemother. 2017, 61, e00937-17. [Google Scholar] [CrossRef]
- Fulsundar, S.; Harms, K.; Flaten, G.E.; Johnsen, P.J.; Chopade, B.A.; Nielsen, K.M. Gene transfer potential of outer membrane vesicles of Acinetobacter baylyi and effects of stress on vesiculation. Appl. Environ. Microbiol. 2014, 80, 3469–3483. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Yang, A.; Liu, P.; Wang, Z.; Jian, Z.; Chen, X.; Yan, Q.; Liang, X.; Liu, W. Outer membrane vesicles transmitting mediate the emergence of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2023, 67, e0144422. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wen, R.; Mu, R.; Chen, X.; Ma, P.; Gu, K.; Huang, Z.; Ju, Z.; Lei, C.; Tang, Y.; et al. Outer membrane vesicles of Avian Pathogenic Escherichia coli mediate the horizontal transmission of blaCTX-M-55. Pathogens 2022, 11, 481. [Google Scholar] [CrossRef]
- Yang, J.; Jia, F.; Qiao, Y.; Hai, Z.; Zhou, X. Correlation between bacterial extracellular vesicles and antibiotics: A potentially antibacterial strategy. Microb. Pathog. 2023, 181, 106167. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Iida, K.I.; Takade, A.; Meno, Y.; Nair, G.B.; Yoshida, S. Release of Shiga toxin by membrane vesicles in Shigella dysenteriae serotype 1 strains and in vitro effects of antimicrobials on toxin production and release. Microbiol. Immunol. 2004, 48, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W.K. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef] [PubMed]
- Crispim, J.S.; Dias, R.S.; Laguardia, C.N.; Araújo, L.C.; da Silva, J.D.; Vidigal, P.M.P.; Sousa, M.P.; da Silva, C.C.; Santana, M.F.; de Paula, S.O. Prophages and their possible involvement in the horizontal transfer of genes by outer membrane vesicles. Gene 2019, 703, 50–57. [Google Scholar] [CrossRef]
- Kharina, A.; Podolich, O.; Faidiuk, I.; Zaika, S.; Haidak, A.; Kukharenko, O.; Zaets, I.; Tovkach, F.; Reva, O.; Kremenskoy, M.; et al. Temperate bacteriophages collected by outer membrane vesicles in Komagataeibacter intermedius. J. Basic Microbiol. 2015, 55, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, J.; Shin, S.G.; Hwang, S. Absolute and relative QPCR quantification of plasmid copy number in Escherichia coli. J. Biotechnol. 2006, 123, 273–280. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | MICs (μg/mL) for | |
|---|---|---|
| E. coli 47EC | ATCC25922 | |
| Rifaximin | 256.000 | 16.000 |
| Tigecycline | 16.000 | 0.125 |
| Ceftazidime | 0.250 | 0.250 |
| Ciprofloxacin | 1.000 | 0.008 |
| Meropenem | 0.030 | 0.031 |
| Chloramphenicol | 256.000 | 4.000 |
| Polymyxin B | 0.125 | 0.500 |
| Gentamicin | 8.000 | 1.000 |
| Mitomycin C | 16.000 | 1.000 |
| Antimicrobial | Copies of tet(X4) Gene in OMVs from E. coli 47EC at Different Conditions (Copies/µL) | ||
|---|---|---|---|
| Without Antimicrobial | 1/4 MIC | 1/2 MIC | |
| Control | 2.937 × 104 | ||
| Mitomycin C | 1.757 × 105 | 6.875 × 105 | |
| Ciprofloxacin | 1.657 × 105 | 5.769 × 105 | |
| Meropenem | 2.488 × 105 | 7.783 × 104 | |
| Tigecycline | 4.125 × 104 | 1.500 × 104 | |
| Ceftazidime | 3.226 × 105 | 5.890 × 105 | |
| Gentamicin | 5.471 × 105 | 2.478 × 105 | |
| Rifaximin | 1.755 × 103 | 2.867 × 104 | |
| Chloramphenicol | 3.067 × 105 | 1.384 × 105 | |
| Polymycin | 3.954 × 105 | 8.425 × 104 | |
| Gene No. | Gene Name | Product Name | Gene Start | Gene End | RNA-Seq Fold Change | Adjusted p Value |
|---|---|---|---|---|---|---|
| gene-PQQ28_RS02015 | rtcB | RNA-splicing ligase RtcB | 351,645 | 352,871 | 6.616 | 1.43 × 10−3 |
| gene-PQQ28_RS00805 | dinD | DNA damage-inducible protein D | 86,763 | 87,587 | 15.868 | 2.47 × 10−13 |
| gene-PQQ28_RS06340 | recA | DNA recombination and repair protein RecA | 1,229,187 | 1,230,248 | 28.095 | 5.40 × 10−14 |
| gene-PQQ28_RS06345 | recX | Recombination regulator RecX | 1,230,317 | 1,230,817 | 27.885 | 3.46 × 10−14 |
| gene-PQQ28_RS06590 | recN | DNA repair protein RecN | 1,270,949 | 1,272,610 | 51.591 | 3.16 × 10−5 |
| gene-PQQ28_RS10705 | ruvA | Holliday junction ATP-dependent DNA helicase RuvA | 2,197,626 | 2,198,237 | 2.2830 | 1.90 × 10−2 |
| gene-PQQ28_RS10770 | yebG | DNA damage-inducible protein YebG | 2,212,855 | 2,213,145 | 10.505 | 5.99 × 10−5 |
| gene-PQQ28_RS11320 | cho | Excinuclease Cho | 2,317,027 | 2,317,914 | 4.024 | 4.23 × 10−2 |
| gene-PQQ28_RS11380 | ydjM | Inner membrane protein YdjM | 2,329,376 | 2,329,966 | 4.493 | 4.49 × 10−2 |
| gene-PQQ28_RS14070 | umuC | DNA polymerase V catalytic protein | 2,872,722 | 2,873,990 | 6.838 | 7.53 × 10−7 |
| gene-PQQ28_RS14075 | umuD | Translesion error-prone DNA polymerase V autoproteolytic subunit | 2,873,990 | 2,874,409 | 12.589 | 8.69 × 10−11 |
| gene-PQQ28_RS15315 | dinI | DNA damage-inducible protein I | 3,067,149 | 3,067,394 | 8.974 | 4.42 × 10−5 |
| gene-PQQ28_RS16320 | sulA | Cell division inhibitor SulA | 3,263,018 | 3,263,527 | 32.872 | 8.06 × 10−7 |
| gene-PQQ28_RS17075 | dinG | ATP-dependent DNA helicase DinG | 3,436,699 | 3,438,849 | 3.112 | 1.16 × 10−3 |
| gene-PQQ28_RS17175 | uvrB | Excinuclease ABC subunit B | 3,455,919 | 3,457,940 | 5.136 | 4.98 × 10−6 |
| gene-PQQ28_RS19460 | yafP | GNAT family N-acetyltransferase | 3,933,638 | 3,934,090 | 8.469 | 5.61 × 10−3 |
| gene-PQQ28_RS19465 | dinB | DNA polymerase IV | 3,934,087 | 3,935,142 | 5.458 | 4.21 × 10−6 |
| gene-PQQ28_RS20795 | polB | DNA polymerase II | 4,218,200 | 4,220,551 | 7.983 | 1.56 × 10−8 |
| gene-PQQ28_RS23100 | ssb1 | Single-stranded DNA-binding protein | 4,690,573 | 4,691,109 | 3.367 | 1.96 × 10−2 |
| gene-PQQ28_RS23105 | uvrA | Excinuclease ABC subunit UvrA | 4,691,364 | 4,694,186 | 5.920 | 6.92 × 10−7 |
| gene-PQQ28_RS23255 | lexA | Transcriptional repressor LexA | 4,728,041 | 4,728,649 | 8.816 | 2.20 × 10−9 |
| gene-PQQ28_RS24565 | uvrD | DNA helicase II | 5,014,795 | 5,016,957 | 2.963 | 6.24 × 10−3 |
| gene-PQQ28_RS01710 | dcrB | Phage sensitivity protein DcrB | 287,181 | 287,738 | 2.643 | 4.40 × 10−3 |
| gene-PQQ28_RS14425 | PQQ28_RS14425 | Phage NinB-Orf DNA recombination | 2,924,894 | 2,925,334 | 586.660 | 4.67 × 10−2 |
| gene-PQQ28_RS14905 | PQQ28_RS14905 | Phage protein | 2,993,478 | 2,993,741 | 352.651 | 4.72 × 10−2 |
| Target Genes | Primers | Primer Sequences (5′-3′) | Length (bp) |
|---|---|---|---|
| recA | recA-F | GGCGTCGATATCGACAACCTG | 138 |
| recA-R | CGCTTTCGGCGTCAGTGC | ||
| cho | cho-F | GGATGAAGCCGCCATGCTAC | 94 |
| cho-R | GATTAATCGCGCTTCAAGG | ||
| lexA | lexA-F | TCATCCGTGATCACATCAGC | 113 |
| lexA-R | TGCCAGCGCCTTCAGATG | ||
| dcrB | dcrB-F | CATGCATGTCTGGTCCGACG | 133 |
| dcrB-R | TTGCAGCTGCGGATCGCG | ||
| PQQ28-RS14425 | PQQ28-RS14425-F | CACTACCCATCGACGACAAG | 98 |
| PQQ28-RS14425-R | ATCGTTCAGCATCGGCCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Li, J.; He, T.; Ji, X.; Wei, R.; Yu, M.; Wang, R. Sub-MIC Antibiotics Modulate Productions of Outer Membrane Vesicles in Tigecycline-Resistant Escherichia coli. Antibiotics 2024, 13, 276. https://doi.org/10.3390/antibiotics13030276
Li Q, Li J, He T, Ji X, Wei R, Yu M, Wang R. Sub-MIC Antibiotics Modulate Productions of Outer Membrane Vesicles in Tigecycline-Resistant Escherichia coli. Antibiotics. 2024; 13(3):276. https://doi.org/10.3390/antibiotics13030276
Chicago/Turabian StyleLi, Qianru, Jun Li, Tao He, Xing Ji, Ruicheng Wei, Meiling Yu, and Ran Wang. 2024. "Sub-MIC Antibiotics Modulate Productions of Outer Membrane Vesicles in Tigecycline-Resistant Escherichia coli" Antibiotics 13, no. 3: 276. https://doi.org/10.3390/antibiotics13030276
APA StyleLi, Q., Li, J., He, T., Ji, X., Wei, R., Yu, M., & Wang, R. (2024). Sub-MIC Antibiotics Modulate Productions of Outer Membrane Vesicles in Tigecycline-Resistant Escherichia coli. Antibiotics, 13(3), 276. https://doi.org/10.3390/antibiotics13030276