

New Insights into the Biological Functions of Essential TsaB/YeaZ Protein in Staphylococcus aureus



Abstract
:1. Introduction
2. Results
2.1. The Deletion of TsaB/YeaZ Leads to Unusual Cellular Morphologies
2.2. Identification of Genes That Are Differentially Expressed during the Downregulation of TsaB/YeaZ
2.3. The Depletion of TsaB/YeaZ Alters Bacterial Autolysis and Cell Wall Recycling
2.4. The Down Regulation of TsaB/YeaZ Affects the Transcriptions of Some tRNA Genes
2.5. The Depletion of TsaB/YeaZ Affects the Transcriptions of Multiple Genes Encoding Virulence Factors
2.6. Identify the Enriched Biological Pathways That Are Significantly Affected by TsaB/YeaZ
2.7. Identify Proteins That Potentially Interact with TsaB/YeaZ in S. aureus
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
4.2. Scanning Electron Microscopy (SEM)
4.3. Transmission Electron Microscopy (TEM)
4.4. RNA Isolation and Purification
4.5. RNA Sequencing (RNA-seq) and Data Analysis
4.5.1. RNA Sequencing
4.5.2. Differential Gene Expression Analysis
4.5.3. KEGG Pathway and Gene Ontology (GO) Biological Process (BP) Functional Enrichment Analysis
4.5.4. Protein and Protein Interaction (PPI) Analysis of TsaB/YeaZ Gene and Interest Trend Genes Using the String 11.5 Database
4.6. Semi-Quantitative Real-Time RT-PCR (qPCR) Analysis
4.7. Triton X-100-Induced Autolysis Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006-0008. [Google Scholar] [CrossRef] [PubMed]
- Vecchietti, D.; Ferrara, S.; Rusmini, R.; Macchi, R.; Milani, M.; Bertoni, G. Crystal structure of YeaZ from Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2016, 470, 460–465. [Google Scholar] [CrossRef]
- Liberati, N.T.; Urbach, J.M.; Miyata, S.; Lee, D.G.; Drenkard, E.; Wu, G.; Villanueva, J.; Wei, T.; Ausubel, F.M. An ordered, nonredundant library of Pseudomonas aeruginosa strain PA14 transposon insertion mutants. Proc. Natl. Acad. Sci. USA 2006, 103, 2833–2838. [Google Scholar] [CrossRef]
- Zalacain, M.; Biswas, S.; Ingraham, K.A.; Ambrad, J.; Bryant, A.; Chalker, A.F.; Iordanescu, S.; Fan, J.; Fan, F.; Lunsford, R.D.; et al. A global approach to identify novel broad-spectrum antibacterial targets among proteins of unknown function. J. Mol. Microbiol. Biotechnol. 2003, 6, 109–126. [Google Scholar] [CrossRef]
- Lei, T.; Yang, J.; Ji, Y. Determination of essentiality and regulatory function of staphylococcal YeaZ in branched-chain amino acid biosynthesis. Virulence 2015, 6, 75–84. [Google Scholar] [CrossRef]
- Thiaville, P.C.; El Yacoubi, B.; Köhrer, C.; Thiaville, J.J.; Deutsch, C.; Iwata-Reuyl, D.; Bacusmo, J.M.; Armengaud, J.; Bessho, Y.; Wetzel, C.; et al. Essentiality of threonylcarbamoyladenosine (t(6)A), a universal tRNA modification, in bacteria. Mol. Microbiol. 2015, 98, 1199–1221. [Google Scholar] [CrossRef]
- Zhang, W.; Collinet, B.; Perrochia, L.; Durand, D.; van Tilbeurgh, H. The ATP-mediated formation of the YgjD-YeaZ-YjeE complex is required for the biosynthesis of tRNA t6A in Escherichia coli. Nucleic Acids Res. 2015, 43, 1804–1817. [Google Scholar] [CrossRef]
- Deutsch, C.; El Yacoubi, B.; de Crécy-Lagard, V.; Iwata-Reuyl, D. Biosynthesis of Threonylcarbamoyl Adenosine (t6A), a Universal tRNA Nucleoside. J. Biol. Chem. 2012, 287, 13666–13673. [Google Scholar] [CrossRef]
- Bacusmo, J.M.; Orsini, S.S.; Hu, J.; DeMott, M.; Thiaville, P.C.; Elfarash, A.; Paulines, M.J.; Rojas-Benítez, D.; Meineke, B.; Deutsch, C.; et al. The t6A modification acts as a positive determinant for the anticodon nuclease PrrC, and is distinctively nonessential in Streptococcus mutans. RNA Biol. 2018, 15, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Britton, T.A.; Guo, H.; Ji, Y. Interaction between two essential, conserved bacterial proteins YeaZ and glycoprotease as a potential antibacterial target in multi-drug-resistant Staphylococcus aureus. Sci. Prog. 2020, 103, 003685041989052. [Google Scholar] [CrossRef]
- Lei, T.; Yang, J.; Zheng, L.; Markowski, T.; Witthuhn, B.A.; Ji, Y. The essentiality of staphylococcal Gcp is independent of its repression of branched-chain amino acids biosynthesis. PLoS ONE 2012, 7, e46836. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Liang, X.; Yang, J.; Yan, M.; Zheng, L.; Walcheck, B.; Ji, Y. The C-terminal domain of the novel essential protein Gcp is critical for interaction with another essential protein YeaZ of Staphylococcus aureus. PLoS ONE 2011, 6, e20163. [Google Scholar] [CrossRef]
- Nichols, C.E.; Johnson, C.; Lockyer, M.; Charles, I.G.; Lamb, H.K.; Hawkins, A.R.; Stammers, D.K. Structural characterization of Salmonella typhimurium YeaZ, an M22 O-sialoglycoprotein endopeptidase homolog. Proteins 2006, 64, 111–123. [Google Scholar] [CrossRef]
- Jeudy, S.; Stelter, M.; Coutard, B.; Kahn, R.; Abergel, C. Preliminary crystallographic analysis of the Escherichia coli YeaZ protein using the anomalous signal of a gadolinium derivative. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Handford, J.I.; Ize, B.; Buchanan, G.; Butland, G.P.; Greenblatt, J.; Emili, A.; Palmer, T. Conserved network of proteins essential for bacterial viability. J. Bacteriol. 2009, 191, 4732–4749. [Google Scholar] [CrossRef]
- Zhao, R.; Chen, J.; Wang, Y.; Li, Y.; Kong, X.; Han, Y. Proteolytic activity of Vibrio harveyi YeaZ is related with resuscitation on the viable but non-culturable state. Lett. Appl. Microbiol. 2020, 71, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; McMullan, D.; Jaroszewski, L.; Krishna, S.S.; Elsliger, M.A.; Yeh, A.P.; Abdubek, P.; Astakhova, T.; Axelrod, H.L.; Carlton, D.; et al. Structure of an essential bacterial protein YeaZ (TM0874) from Thermotoga maritima at 2.5 A resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1230–1236. [Google Scholar] [CrossRef]
- Aydin, I.; Saijo-Hamano, Y.; Namba, K.; Thomas, C.; Roujeinikova, A. Structural analysis of the essential resuscitation promoting factor YeaZ suggests a mechanism of nucleotide regulation through dimer reorganization. PLoS ONE 2011, 6, e23245. [Google Scholar] [CrossRef]
- Nichols, C.E.; Lamb, H.K.; Thompson, P.; Omari, K.E.; Lockyer, M.; Charles, I.; Hawkins, A.R.; Stammers, D.K. Crystal structure of the dimer of two essential Salmonella typhimurium proteins, YgjD & YeaZ and calorimetric evidence for the formation of a ternary YgjD-YeaZ-YjeE complex. Protein Sci. 2013, 22, 628–640. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Wang, Y.; Ma, D.; Rui, W. The effects of the recombinant YeaZ of Vibrio harveyi on the resuscitation and growth of soil bacteria in extreme soil environment. PeerJ 2020, 8, e10342. [Google Scholar] [CrossRef]
- Panutdaporn, N.; Kawamoto, K.; Asakura, H.; Makino, S.I. Resuscitation of the viable but non-culturable state of Salmonella enterica serovar Oranienburg by recombinant resuscitation-promoting factor derived from Salmonella Typhimurium strain LT2. Int. J. Food Microbiol. 2006, 106, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Shleeva, M.; Mukamolova, G.V.; Young, M.; Williams, H.D.; Kaprelyants, A.S. Formation of ‘non-culturable’ cells of Mycobacterium smegmatis in stationary phase in response to growth under suboptimal conditions and their Rpf-mediated resuscitation. Microbiology 2004, 150, 1687–1697. [Google Scholar] [CrossRef]
- Ravagnani, A.; Finan, C.L.; Young, M. A novel firmicute protein family related to the actinobacterial resuscitation-promoting factors by non-orthologous domain displacement. BMC Genom. 2005, 6, 39. [Google Scholar] [CrossRef]
- Bae, T.; Banger, A.K.; Wallace, A.; Glass, E.M.; Aslund, F.; Schneewind, O.; Missiakas, D.M. Staphylococcus aureus virulence genes identified by bursa aurealis mutagenesis and nematode killing. Proc. Natl. Acad. Sci. USA 2004, 101, 12312–12317. [Google Scholar] [CrossRef]
- Chaudhuri, R.R.; Allen, A.G.; Owen, P.J.; Shalom, G.; Stone, K.; Harrison, M.; A Burgis, T.; Lockyer, M.; Garcia-Lara, J.; Foster, S.J.; et al. Comprehensive identification of essential Staphylococcus aureus genes using Transposon-Mediated Differential Hybridisation (TMDH). BMC Genom. 2009, 10, 291. [Google Scholar] [CrossRef] [PubMed]
- Borisova, M.; Gaupp, R.; Duckworth, A.; Schneider, A.; Dalügge, D.; Mühleck, M.; Deubel, D.; Unsleber, S.; Yu, W.; Muth, G.; et al. Peptidoglycan Recycling in Gram-Positive Bacteria Is Crucial for Survival in Stationary Phase. mBio 2016, 7, e00923-16. [Google Scholar] [CrossRef] [PubMed]
- Ranjit, D.K.; Endres, J.L.; Bayles, K.W. Staphylococcus aureus CidA and LrgA proteins exhibit holin-like properties. J. Bacteriol. 2011, 193, 2468–2476. [Google Scholar] [CrossRef]
- Endres, J.L.; Chaudhari, S.S.; Zhang, X.; Prahlad, J.; Wang, S.-Q.; Foley, L.A.; Luca, S.; Bose, J.L.; Thomas, V.C.; Bayles, K.W. The Staphylococcus aureus CidA and LrgA Proteins Are Functional Holins Involved in the Transport of By-Products of Carbohydrate Metabolism. mBio 2021, 13, e0282721. [Google Scholar] [CrossRef]
- Murphy, F.V., IV; Ramakrishnan, V.; Malkiewicz, A.; Agris, P.F. The role of modifications in codon discrimination by tRNA(Lys)UUU. Nat. Struct. Mol. Biol. 2004, 11, 1186–1191. [Google Scholar] [CrossRef]
- White, M.L.; Hough-Neidig, A.; Khan, S.J.; Eswara, P.J. MraZ Transcriptionally Controls the Critical Level of FtsL Required for Focusing Z-Rings and Kickstarting Septation in Bacillus subtilis. J. Bacteriol. 2022, 204, e0024322. [Google Scholar] [CrossRef]
- Yarwood, J.M.; McCormick, J.K.; Schlievert, P.M. Identification of a novel two-component regulatory system that acts in global regulation of virulence factors of Staphylococcus aureus. J. Bacteriol. 2001, 183, 1113–1123. [Google Scholar] [CrossRef]
- Pragman, A.A.; Ji, Y.; Schlievert, P.M. Repression of Staphylococcus aureus SrrAB using inducible antisense srrA alters growth and virulence factor transcript levels. Biochemistry 2007, 46, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Cleverley, R.M.; Rismondo, J.; Lockhart-Cairns, M.P.; Van Bentum, P.T.; Egan, A.J.; Vollmer, W.; Halbedel, S.; Baldock, C.; Breukink, E.; Lewis, R.J. Subunit Arrangement in GpsB, a Regulator of Cell Wall Biosynthesis. Microb. Drug Resist. 2016, 22, 446–460. [Google Scholar] [CrossRef]
- Wang, B.; Duan, J.; Jin, Y.; Zhan, Q.; Xu, Y.; Zhao, H.; Wang, X.; Rao, L.; Guo, Y.; Yu, F. Functional Insights of MraZ on the Pathogenicity of Staphylococcus aureus. Infect. Drug Resist. 2021, 14, 4539–4551. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Yu, C.; Bayles, K.; Lasa, I.; Ji, Y. Conditional mutation of an essential putative glycoprotease eliminates autolysis in Staphylococcus aureus. J. Bacteriol. 2007, 189, 2734–2742. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Kluj, R.M.; Mühleck, M.; Walter, A.; Unsleber, S.; Hottmann, I.; Borisova, M. Bacteria’s different ways to recycle their own cell wall. Int. J. Med. Microbiol. 2019, 309, 151326. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, R.C.; Liu, R.; Burgin, D.J.; Otto, M. Understanding mechanisms of virulence in MRSA: Implications for antivirulence treatment strategies. Expert Rev. Anti-Infect. Ther. 2023, 21, 911–928. [Google Scholar] [CrossRef]
- Nikolic, P.; Mudgil, P. The Cell Wall, Cell Membrane and Virulence Factors of Staphylococcus aureus and Their Role in Antibiotic Resistance. Microorganisms 2023, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.S.; Cunneen, T.; Lee, C.Y. Sequence analysis and molecular characterization of genes required for the biosynthesis of type 1 capsular polysaccharide in Staphylococcus aureus. J. Bacteriol. 1994, 176, 7005–7016. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.L. Effects of overproduction of single-stranded DNA-binding protein on RecA protein-dependent processes in Escherichia coli. J. Mol. Biol. 1987, 194, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Katz, C.; Cohen-Or, I.; Gophna, U.; Ron, E.Z. The ubiquitous conserved glycopeptidase Gcp prevents accumulation of toxic glycated proteins. mBio 2010, 1, e00195-10. [Google Scholar] [CrossRef] [PubMed]
- Bergmiller, T.; Peña-Miller, R.; Boehm, A.; Ackermann, M. Single-cell time-lapse analysis of depletion of the universally conserved essential protein YgjD. BMC Microbiol. 2011, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Cassels, R.; Oliva, B.; Knowles, D. Occurrence of the regulatory nucleotides ppGpp and pppGpp following induction of the stringent response in staphylococci. J. Bacteriol. 1995, 177, 5161–5165. [Google Scholar] [CrossRef] [PubMed]
- Lange, R.; Hengge-Aronis, R. Growth phase-regulated expression of bolA and morphology of stationary-phase Escherichia coli cells are controlled by the novel sigma factor sigma S. J. Bacteriol. 1991, 173, 4474–4481. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, G.; Ron, E.Z.; Glaser, G. ppGpp-mediated regulation of DNA replication and cell division in Escherichia coli. Curr. Microbiol. 1995, 30, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Metzger, S.; Schreiber, G.; Aizenman, E.; Cashel, M.; Glaser, G. Characterization of the relA1 mutation and a comparison of relA1 with new relA null alleles in Escherichia coli. J. Biol. Chem. 1989, 264, 21146–21152. [Google Scholar] [CrossRef]
- Hashimoto, C.; Sakaguchi, K.; Taniguchi, Y.; Honda, H.; Oshima, T.; Ogasawara, N.; Kato, J.-I. Effects on transcription of mutations in ygjD, yeaZ, and yjeE genes, which are involved in a universal tRNA modification in Escherichia coli. J. Bacteriol. 2011, 193, 6075–6079. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Sen, S.; Sinha, A.; Wilkinson, B.J.; Heinrichs, D.E. The role of two branched-chain amino acid transporters in Staphylococcus aureus growth, membrane fatty acid composition and virulence. Mol. Microbiol. 2016, 102, 850–864. [Google Scholar] [CrossRef] [PubMed]
- Brinsmade, S.R.; Kleijn, R.J.; Sauer, U.; Sonenshein, A.L. Regulation of CodY activity through modulation of intracellular branched-chain amino acid pools. J. Bacteriol. 2010, 192, 6357–6368. [Google Scholar] [CrossRef] [PubMed]
- Grandoni, J.A.; Zahler, S.A.; Calvo, J.M. Transcriptional regulation of the ilv-leu operon of Bacillus subtilis. J. Bacteriol. 1992, 174, 3212–3219. [Google Scholar] [CrossRef] [PubMed]
- Shivers, R.P.; Sonenshein, A.L. Bacillus subtilis ilvB operon: An intersection of global regulons. Mol. Microbiol. 2005, 56, 1549–1559. [Google Scholar] [CrossRef]
- Lincoln, R.A.; Leigh, J.A.; Jones, N.C. The amino acid requirements of Staphylococcus aureus isolated from cases of bovine mastitis. Vet. Microbiol. 1995, 45, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Onoue, Y.; Mori, M. Amino acid requirements for the growth and enterotoxin production by Staphylococcus aureus in chemically defined media. Int. J. Food Microbiol. 1997, 36, 77–82. [Google Scholar] [CrossRef]
- Majerczyk, C.D.; Dunman, P.M.; Luong, T.T.; Lee, C.Y.; Sadykov, M.R.; Somerville, G.A.; Bodi, K.; Sonenshein, A.L. Direct targets of CodY in Staphylococcus aureus. J. Bacteriol. 2010, 192, 2861–2877. [Google Scholar] [CrossRef]
- Ludwig, H.; Meinken, C.; Matin, A.; Stülke, J. Insufficient expression of the ilv-leu operon encoding enzymes of branched-chain amino acid biosynthesis limits growth of aBacillus subtilis ccpAmutant. J. Bacteriol. 2002, 184, 5174–5178. [Google Scholar] [CrossRef]
- Tojo, S.; Satomura, T.; Morisaki, K.; Deutscher, J.; Hirooka, K.; Fujita, Y. Elaborate transcription regulation of the Bacillus subtilis ilv-leu operon involved in the biosynthesis of branched-chain amino acids through global regulators of CcpA, CodY and TnrA. Mol. Microbiol. 2005, 56, 1560–1573. [Google Scholar] [CrossRef]
- Pohl, K.; Francois, P.; Stenz, L.; Schlink, F.; Geiger, T.; Herbert, S.; Goerke, C.; Schrenzel, J.; Wolz, C. CodY in Staphylococcus aureus: A regulatory link between metabolism and virulence gene expression. J. Bacteriol. 2009, 191, 2953–2963. [Google Scholar] [CrossRef]
- Gutiérrez-Preciado, A.; Henkin, T.M.; Grundy, F.J.; Yanofsky, C.; Merino, E. Biochemical features and functional implications of the RNA-based T-box regulatory mechanism. Microbiol. Mol. Biol. Rev. 2009, 73, 36–61. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Haldeman, M.T.; Hornblow, G.M.; Ward, J.M.; Chalker, A.F.; Henkin, T.M. The Staphylococcus aureus ileS gene, encoding isoleucyl-tRNA synthetase, is a member of the T-box family. J. Bacteriol. 1997, 179, 3767–3772. [Google Scholar] [CrossRef] [PubMed]
- Reiß, S.; Pané-Farré, J.; Fuchs, S.; François, P.; Liebeke, M.; Schrenzel, J.; Lindequist, U.; Lalk, M.; Wolz, C.; Hecker, M.; et al. Global analysis of the Staphylococcus aureus response to mupirocin. Antimicrob. Agents Chemother. 2012, 56, 787–804. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Zhang, Y.; Yang, J.; Silverstein, K.; Ji, Y. Complete Genome Sequence of Hospital-Acquired Methicillin-Resistant Staphylococcus aureus Strain WCUH29. Microbiol. Resour. Announc. 2019, 8, e00551-19. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Yang, J.; Becker, A.; Ji, Y. Identification of Target Genes Mediated by Two-Component Regulators of Staphylococcus aureus Using RNA-seq Technology. Methods Mol. Biol. 2020, 2069, 125–138. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Fold Change (Decrease) a |
|---|---|
| tsaB/yeaZ | 1.99 ± 0.52 |
| cidA | 1.88 ± 0.25 |
| murQ | 2.75 ± 0.48 |
| femA | 1.61 ± 0.19 |
| lytM | 1.97 ± 0.08 |
| lytN | 2.04 ± 0.31 |
| atl | 2.64 ± 0.46 |
| srrA | 3.28 ± 0.35 |
| hlgC | 3.06 ± 0.48 |
| lukH | 16.75 ± 2.32 |
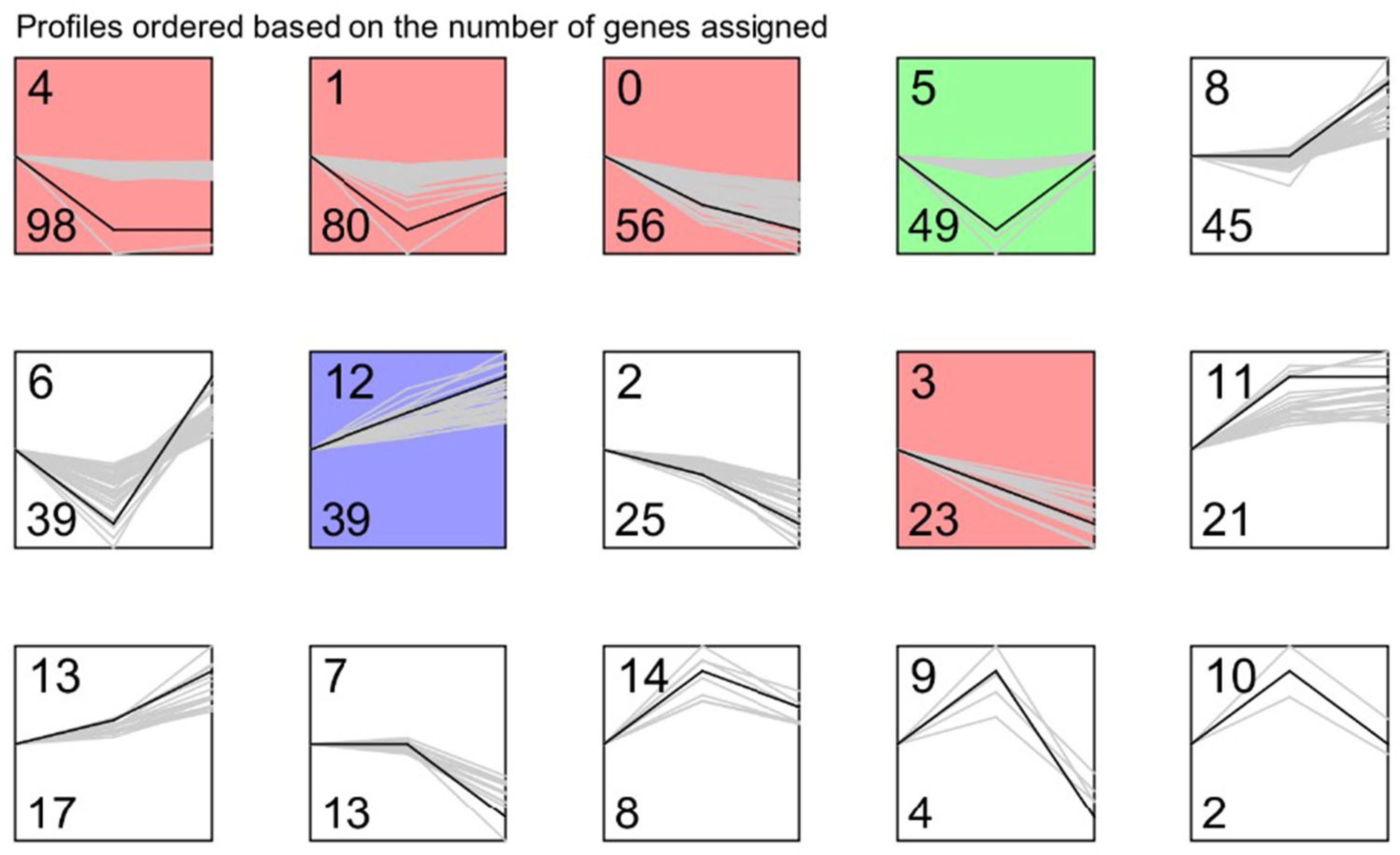
| Profile | Count | Trend | Class |
|---|---|---|---|
| 0 | 56 | Down-down | Interest trend class I gene |
| 2 | 25 | Down-down | |
| 3 | 23 | Down-down | |
| 12 | 39 | Up-up | |
| 13 | 17 | Up-up | |
| 9 | 4 | Up-down | Interest trend class II gene |
| 10 | 2 | Up-down | |
| 14 | 8 | Up-down | |
| 1 | 80 | Down-up | |
| 5 | 49 | Down-up | |
| 6 | 39 | Down-up | |
| 4 | 98 | Down-flat | Interest trend class III gene |
| 11 | 21 | Up-flat | |
| 8 | 45 | Flat-up | Interest trend class IV gene |
| 7 | 13 | Flat-down |
| Pathway ID | Description of Pathway | Count | p-Value |
|---|---|---|---|
| ko01110 | Biosynthesis of secondary metabolites | 21 | 5.55 × 10−8 |
| ko01120 | Microbial metabolism in diverse environments | 14 | 8.07 × 10−5 |
| ko02010 | ABC transporters | 16 | 0.004460232 |
| ko00561 | Glycerolipid metabolism | 5 | 0.008120074 |
| ko00450 | Selenocompound metabolism | 3 | 0.009847925 |
| ko00910 | Nitrogen metabolism | 4 | 0.019201212 |
| ko00562 | Inositol phosphate metabolism | 2 | 0.03031329 |
| ko02024 | Quorum sensing | 8 | 0.043943233 |
| ko01230 | Biosynthesis of amino acids | 14 | 0.044417679 |
| Gene Name | Gene orf Number | Degree | Profile | Type |
|---|---|---|---|---|
| tpi | E5491_RS04085 | 9 | 3 | down-down |
| thrB | E5491_RS06900 | 6 | 0 | down-down |
| leuC | E5491_RS11545 | 5 | 0 | down-down |
| dapH | E5491_RS07270 | 4 | 0 | down-down |
| tsaB/yeaZ | E5491_RS11500 | 4 | ||
| groEL | E5491_RS11380 | 4 | 2 | down-down |
| yidC | E5491_RS11755 | 4 | 13 | up-up |
| ilvA | E5491_RS11555 | 4 | 0 | down-down |
| dapB | E5491_RS07265 | 3 | 0 | down-down |
| metE | E5491_RS01730 | 3 | 3 | down-down |
| plsY | E5491_RS07035 | 3 | 13 | up-up |
| Group | IPTG – | IPTG + | DEG_Up | DEG_Down | Total |
|---|---|---|---|---|---|
| yeaZ mutant OD 0.2 | yeaZ_02 | yeaZ 02 | 292 | 98 | 390 |
| yeaZ mutant OD 0.5 | yeaZ_05 | yeaZ 05 | 97 | 118 | 215 |
| yeaZ mutant OD 1.0 | yeaZ_10 | yeaZ 10 | 241 | 126 | 367 |
| Control OD 0.2 | C_02 | C02 | 2 | 3 | 5 |
| Control OD 0.5 | C_05 | C05 | 0 | 0 | 0 |
| Control OD 1.0 | C_10 | C10 | 0 | 0 | 0 |
| Primer Name | Oligo Sequences (5’-3’) |
|---|---|
| yeaZRTfor243 | ACTGCTTGTCGTCTTGCATC |
| yeaZRTrev370 | AACGCTAAAACATTAGCGTATGCGTTAG |
| cidA RT For | GTCTTTTTCTTCATACCGTCAGT |
| cidA RT Rev | TCATTCATAAGCGTCTACACCT |
| murQ RT For | GCTATGACGATGGCTGTAGAAG |
| murQ RT Rev | CACTCGCGGCAATTCCTATAA |
| femA RT For | TCATCGATTACAGACGAAGACAC |
| femA RT Rev | TCTTTTAGTTTAGACGGCGCAACC |
| lytM-F1 | GCAGGAGATAACAATGACTACACAG |
| lytM-R1 | GCTGTCGCTTTACTTGCTGAT |
| lytN RT For | AGCTGAACCTGGGGACTTAG |
| lytN RT Rev | CAACTTTATGTGCAACCTCTGC |
| atl RT For | GCTGGTTATAGTTTAGTTGATGATG |
| atl RT Rev | GGTTGTGCTGAAGCGCTAAAAG |
| srrAB-RT.2A | TGCCTGAAATGGATGGTATCC |
| srrAB-RT.2B | AACACGGTTTGTTTCTTCACC |
| hlgC RT For | CCCTCTTGCCAATCCGTTATTA |
| hlgC RT Rev | ATATCGCTTCCTTTACCGATGTC |
| lukH RT For | CCACTTCTTACTAATGCTGGGT |
| lukH RT Rev | TGACTTGAAGTATGGTGGAGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Lei, T.; Yang, J.; Wang, Y.; Wang, Y.; Ji, Y. New Insights into the Biological Functions of Essential TsaB/YeaZ Protein in Staphylococcus aureus. Antibiotics 2024, 13, 393. https://doi.org/10.3390/antibiotics13050393
Guo H, Lei T, Yang J, Wang Y, Wang Y, Ji Y. New Insights into the Biological Functions of Essential TsaB/YeaZ Protein in Staphylococcus aureus. Antibiotics. 2024; 13(5):393. https://doi.org/10.3390/antibiotics13050393
Chicago/Turabian StyleGuo, Haiyong, Ting Lei, Junshu Yang, Yue Wang, Yifan Wang, and Yinduo Ji. 2024. "New Insights into the Biological Functions of Essential TsaB/YeaZ Protein in Staphylococcus aureus" Antibiotics 13, no. 5: 393. https://doi.org/10.3390/antibiotics13050393
APA StyleGuo, H., Lei, T., Yang, J., Wang, Y., Wang, Y., & Ji, Y. (2024). New Insights into the Biological Functions of Essential TsaB/YeaZ Protein in Staphylococcus aureus. Antibiotics, 13(5), 393. https://doi.org/10.3390/antibiotics13050393