
Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli


Abstract
:1. Introduction
2. Results
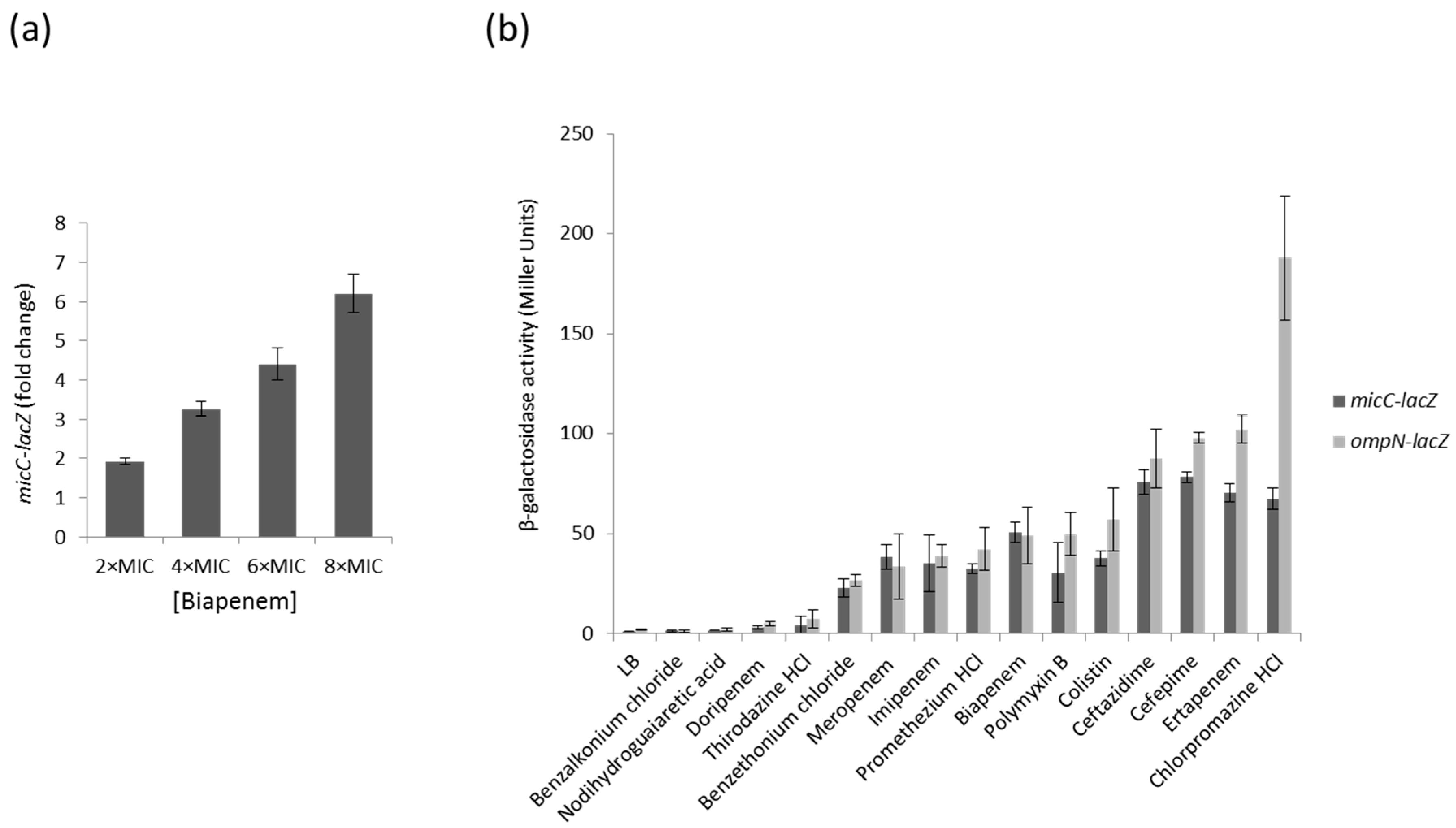
2.1. Screening of MicC and OmpN Inducing Conditions Using LacZ Transcriptional Fusions and BiologTM Plates
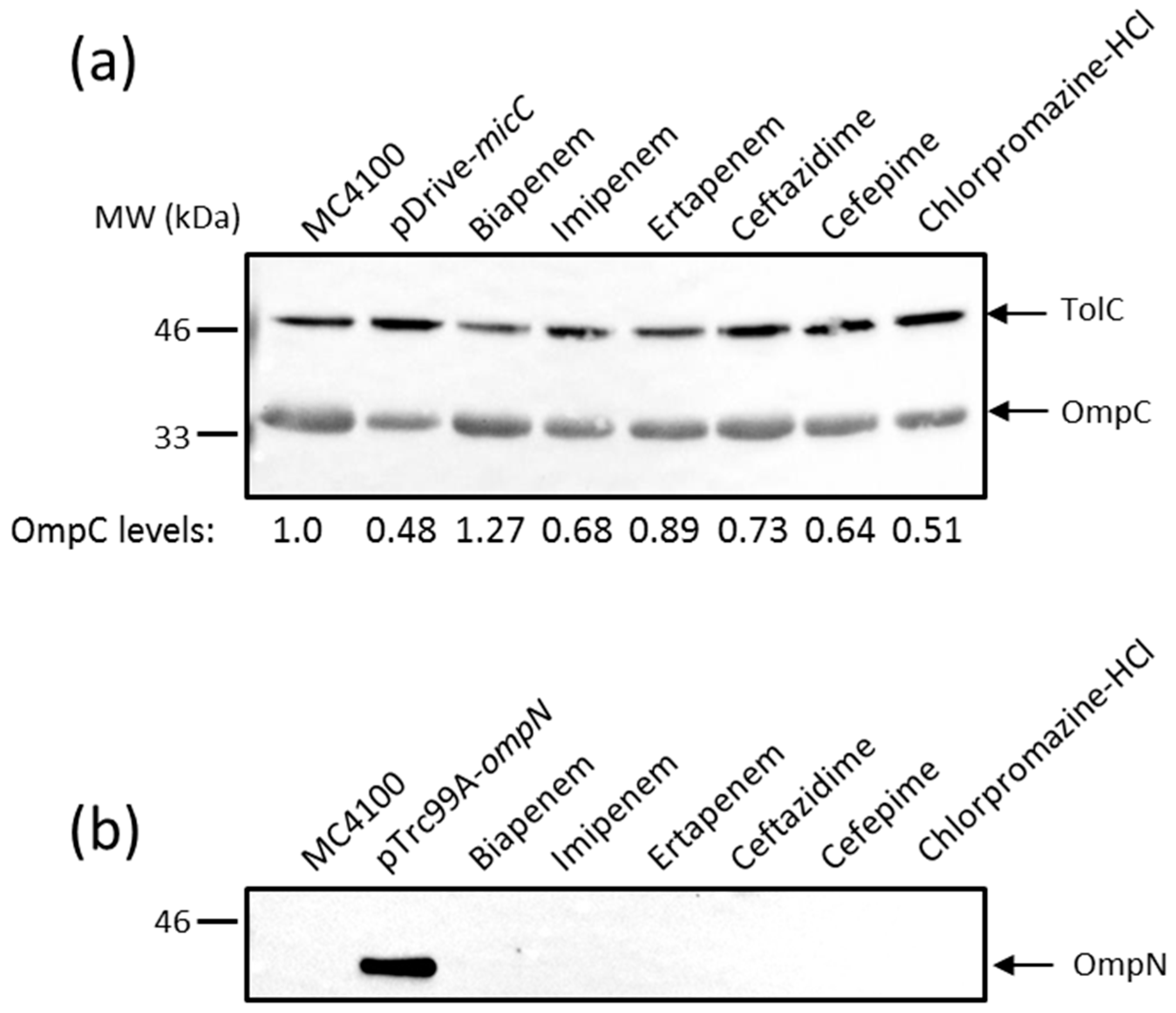
2.2. Effects of MicC and OmpN Inducing Conditions on the Expression Levels of OmpC and OmpN
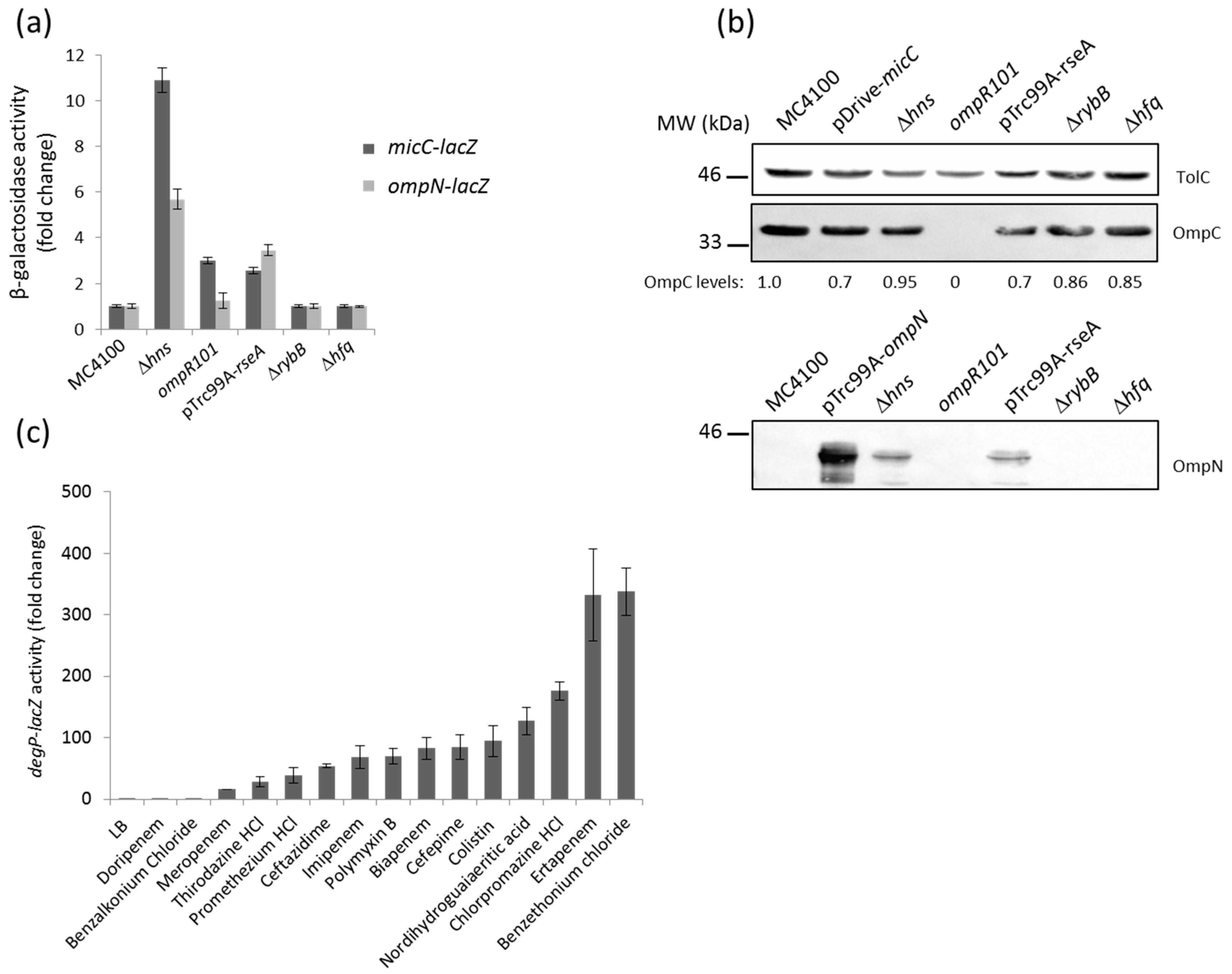
2.3. Identification of Genetic Factors That Impact on MicC and OmpN Expression
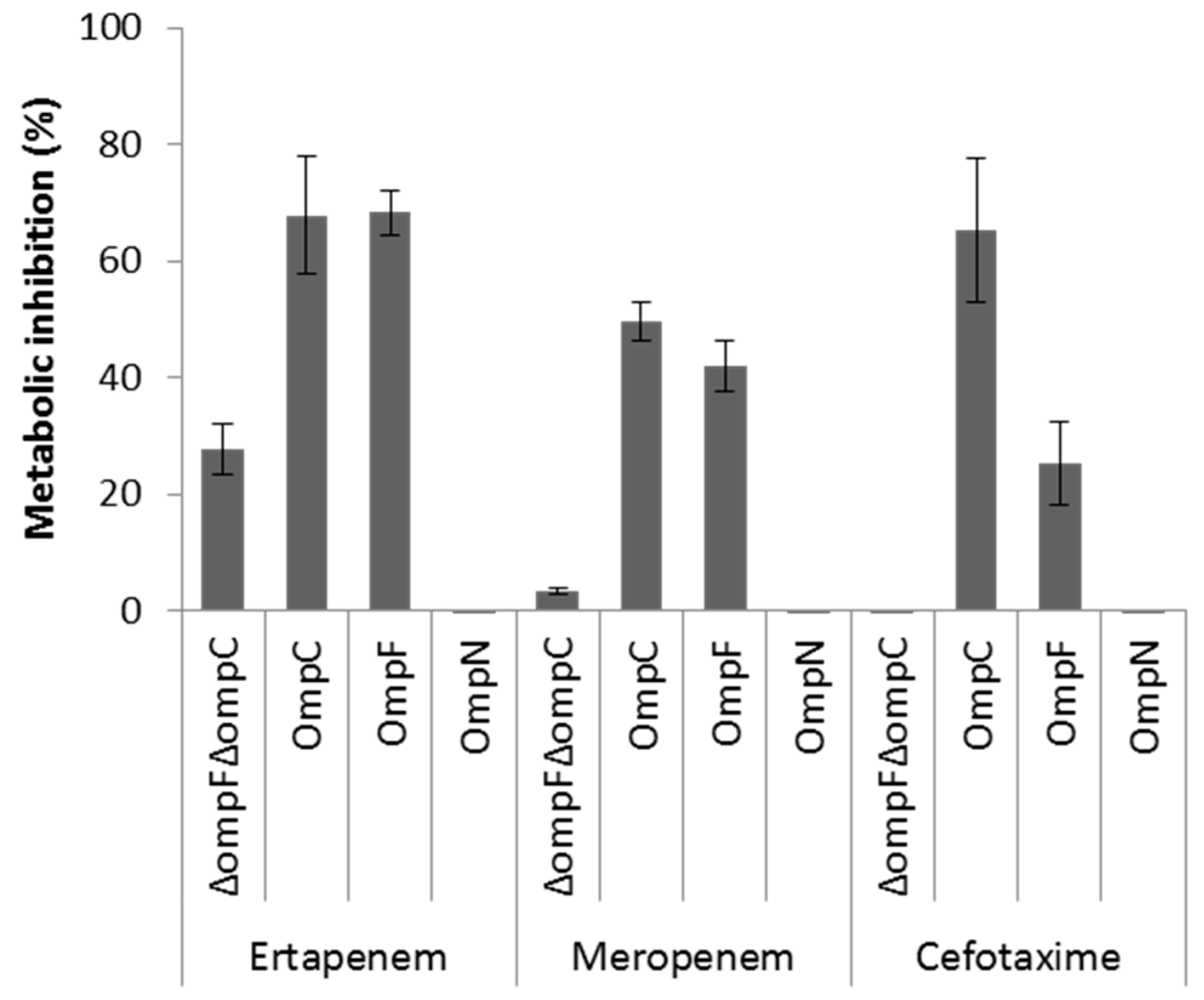
2.4. Role of OmpN in Antibiotic Translocation
3. Discussion
4. Materials and Methods
4.1. Plasmids and Bacterial Strains
4.2. Plasmid Construction
4.3. β-Galactosidase Assays
4.4. Determination of Minimal Inhibitory Concentrations (MIC)
4.5. Preparation of the Microtiter Plates for β-Galactosidase Assays
4.6. Preparation of OM Extracts
4.7. SDS-PAGE and Western Blot Analysis
4.8. Whole Cell-Based Viability Assay
4.9. Colicin Killing Assays
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- The Review on Antimicrobial Resistance Chaired by Jim O’Neill. Tackling Drug-Resistant Infections Globally: Final Report and Recommandations. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 3 March 2016).
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance. Available online: http://apps.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf (accessed on 3 March 2016).
- National Institute of Allergy and Infectious Diseases (NIAID). NIAID’s Antibacterial Resistance Program: Currtent Status and Future Directions. Available online: https://www.niaid.nih.gov/sites/default/files/arstrategicplan2014.pdf (accessed on 3 March 2016).
- Stavenger, R.A.; Winterhalter, M. Translocation project: How to get good drugs into bad bugs. Sci. Transl. Med. 2014, 6, 228ed7. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Masi, M.; Réfregiers, M.; Pos, K.M.; Pagès, J.-M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Pagès, J.-M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Pagès, J.-M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Bolla, J.-M.; James, C.E.; Lavigne, J.-P.; Chevalier, J.; Garnotel, E.; Molitor, A. Membrane permeability and regulation of drug “influx and efflux” in enterobacterial pathogens. Curr. Drug Targets 2008, 9, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Pratt, L.A.; Hsing, W.; Gibson, K.E.; Silhavy, T.J. From acids to osmZ: Multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol. Microbiol. 1996, 20, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Guest, R.L.; Raivio, T.L. Role of the Gram-negative envelope stress response in the presence of antimicrobial agents. Trends Microbiol. 2016, 24, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Gerken, H.; Charlson, E.S.; Cicirelli, E.M.; Kenney, L.J.; Misra, R. MzrA: A novel modulator of the EnvZ/OmpR two-component regulon. Mol. Microbiol. 2009, 72, 1408–1422. [Google Scholar] [CrossRef] [PubMed]
- Philippe, N.; Maigre, L.; Santini, S.; Pinet, E.; Claverie, J.-M.; Davin-Régli, A.V.; Pagès, J.-M.; Masi, M. In vivo evolution of bacterial resistance in two cases of Enterobacter aerogenes infections during treatment with imipenem. PLoS ONE 2015, 10, e0138828. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Chou, M.Y.; Inouye, M. A unique mechanism regulating gene expression: Translational inhibition by a complementary RNA transcript (micRNA). Proc. Natl. Acad. Sci. USA 1984, 81, 1966–1970. [Google Scholar] [CrossRef] [PubMed]
- Delihas, N. Discovery and characterization of the first non-coding RNA that regulates gene expression, micF RNA: A historical perspective. World J. Biol. Chem. 2015, 6, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M. The first demonstration of RNA interference to inhibit mRNA function. Gene 2016, 592, 332–333. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Delihas, N. micF RNA binds to the 5′ end of ompF mRNA and to a protein from Escherichia coli. Biochemistry 1990, 29, 9249–9256. [Google Scholar] [CrossRef] [PubMed]
- Delihas, N.; Forst, S. MicF: An antisense RNA gene involved in response of Escherichia coli to global stress factors. J. Mol. Biol. 2001, 313, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ramani, N.; Hedeshian, M.; Freundlich, M. micF antisense RNA has a major role in osmoregulation of OmpF in Escherichia coli. J. Bacteriol. 1994, 176, 5005–5010. [Google Scholar] [CrossRef] [PubMed]
- Chou, J.H.; Greenberg, J.T.; Demple, B. Posttranscriptional repression of Escherichia coli OmpF protein in response to redox stress: Positive control of the micF antisense RNA by the soxRS locus. J. Bacteriol. 1993, 175, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; McMurry, L.M.; Levy, S.B. MarA locus causes decreased expression of OmpF porin in multiple-antibiotic-resistant (Mar) mutants of Escherichia coli. J. Bacteriol. 1988, 170, 5416–5422. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, A.; Blyn, L.B.; Storz, G. MicC, a second small-RNA regulator of Omp protein expression in Escherichia coli. J. Bacteriol. 2004, 186, 6689–6697. [Google Scholar] [CrossRef] [PubMed]
- Prilipov, A.; Phale, P.S.; Koebnik, R.; Widmer, C.; Rosenbusch, J.-P. Identification and characterization of two quiescent porin genes, nmpC and ompN, in Escherichia coli BE. J. Bacteriol. 1998, 180, 3388–3392. [Google Scholar] [PubMed]
- Johansen, J.; Rasmussen, A.A.; Overgaard, M.; Valentin-Hansen, P. Conserved small non-coding RNAs that belong to the sigmaE regulon: Role in down-regulation of outer membrane proteins. J. Mol. Biol. 2006, 364, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Douchin, V.; Bohn, C.; Bouloc, P. Down-regulation of porins by a small RNA bypasses the essentiality of the regulated intramembrane proteolysis protease RseP in Escherichia coli. J. Biol. Chem. 2006, 281, 12253–12259. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Keller, M.; Vuong, P.; Misra, R. Novel mechanism of Escherichia coli porin regulation. J. Bacteriol. 2006, 188, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Papenfort, K. Small non-coding RNAs and the bacterial outer membrane. Curr. Opin. Microbiol. 2006, 9, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Guillier, M.; Gottesman, S.; Storz, G. Modulating the outer membrane with small RNAs. Genes Dev. 2006, 20, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Bollet, C.; Chevalier, J.; Malléa, M.; Pagès, J.-M.; Davin-Regli, A. mar operon involved in multidrug resistance of Enterobacter aerogenes. Antimicrob. Agents Chemother. 2002, 46, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Chevalier, J.; Bollet, C.; Pages, J.-M.; Davin-Regli, A. RamA is an alternate activator of the multidrug resistance cascade in Enterobacter aerogenes. Antimicrob. Agents Chemother. 2004, 48, 2518–2523. [Google Scholar] [CrossRef] [PubMed]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.M.; Rhodius, V.A.; Gottesman, S. SigmaE regulates and is regulated by a small RNA in Escherichia coli. J. Bacteriol. 2007, 189, 4243–4256. [Google Scholar] [CrossRef] [PubMed]
- Skovierova, H.; Rowley, G.; Rezuchova, B.; Homerova, D.; Lewis, C.; Roberts, M.; Kormanec, J. Identification of the sigmaE regulon of Salmonella enterica serovar Typhimurium. Microbiology 2006, 152, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Johansen, J.; Eriksen, M.; Kallipolitis, B.; Valentin-Hansen, P. Down-regulation of outer membrane proteins by noncoding RNAs: Unraveling the cAMP-CRP- and sigmaE-dependent CyaR-OmpX regulatory case. J. Mol. Biol. 2008, 383, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Pfeiffer, V.; Lucchini, S.; Sonawane, A.; Hinton, J.C.; Vogel, J. Systematic deletion of Salmonella small RNA genes identifies CyaR, a conserved CRP-dependent riboregulator of OmpX synthesis. Mol. Microbiol. 2008, 68, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Mandin, P.; Gottesman, S. Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA. EMBO J. 2010, 29, 3094–3107. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.; Storz, G. Reprogramming of anaerobic metabolism by the FnrS small RNA. Mol. Microbiol. 2010, 75, 1215–1231. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Six, D.A.; Lee, H.J.; Raetz, C.R.; Gottesman, S. Complex transcriptional and post-transcriptional regulation of an enzyme for lipopolysaccharide modification. Mol. Microbiol. 2013, 89, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.S.; Papenfort, K.; Berger, A.A.; Vogel, J. A conserved RpoS-dependent small RNA controls the synthesis of major porin OmpD. Nucleic Acids Res. 2012, 40, 3623–3640. [Google Scholar] [CrossRef] [PubMed]
- Grabowicz, M.; Koren, D.; Silhavy, T.J. The CpxQ sRNA negatively regulates Skp to prevent mistargeting of β-barrel outer membrane proteins into the cytoplasmic membrane. mBio 2016, 7, e00312-16. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Vogel, J. A 3′ UTR-derived small RNA provides the regulatory noncoding arm of the inner membrane stress response. Mol. Cell 2016, 61, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Snyder, W.B.; Davis, L.J.; Danese, P.N.; Cosma, C.L.; Silhavy, T.J. Overproduction of NlpE, a new outer membrane lipoprotein, suppresses the toxicity of periplasmic LacZ by activation of the Cpx signal transduction pathway. J. Bacteriol. 1995, 177, 4216–4223. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, G.; Blankschien, M.; Herman, C.; Gross, C.A.; Rhodius, V.A. Regulon and promoter analysis of the E. coli heat-shock factor, sigma32, reveals a multifaceted cellular response to heat stress. Genes Dev. 2006, 20, 1776–1789. [Google Scholar] [CrossRef] [PubMed]
- Gogol, E.B.; Rhodius, V.A.; Papenfort, K.; Vogel, J.; Gross, C.A. Small RNAs endow a transcriptional activator with essential repressor functions for single-tier control of a global stress regulon. Proc. Natl. Acad. Sci. USA 2011, 108, 12875–12880. [Google Scholar] [CrossRef] [PubMed]
- Raivio, T.L.; Silhavy, T.J. Transduction of envelope stress in Escherichia coli by the Cpx two-component system. J. Bacteriol. 1997, 179, 7724–7733. [Google Scholar] [CrossRef] [PubMed]
- Viveiros, M.; Dupont, M.; Rodrigues, L.; Couto, I.; Davin-Regli, A.; Martins, M.; Pagès, J.-M.; Amaral, L. Antibiotic stress, genetic response and altered permeability of E. coli. PLoS ONE 2007, 2, e365. [Google Scholar] [CrossRef] [PubMed]
- Deighan, P.; Free, A.; Dorman, C.J. A role for the Escherichia coli H-NS-like protein StpA in OmpF porin expression through modulation of micF RNA stability. Mol. Microbiol. 2000, 38, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ueguchi, C.; Mizuno, T. H-NS regulates OmpF expression through micF antisense RNA in Escherichia coli. J. Bacteriol. 1996, 178, 3650–3653. [Google Scholar] [CrossRef] [PubMed]
- Hommais, F.; Krin, E.; Laurent-Winter, C.; Soutourina, O.; Malpertuy, A.; Le Caer, J.P.; Danchin, A.; Bertin, P. Large-scale monitoring of pleiotropic regulation of gene expression by the prokaryotic nucleoid-associated protein, H-NS. Mol. Microbiol. 2001, 40, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Doménech-Sánchez, A.; Hernández-Allés, S.; Martínez-Martínez, L.; Benedí, V.J.; Albertí, S. Identification and characterization of a new porin gene of Klebsiella pneumoniae: Its role in beta-lactam antibiotic resistance. J. Bacteriol. 1999, 181, 2726–2732. [Google Scholar] [PubMed]
- Housden, N.G.; Kleanthous, C. Colicin translocation across the Escherichia coli outer membrane. Biochem. Soc. Trans. 2012, 40, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Wassarman, K.M.; Rosenow, C.; Tjaden, B.C.; Storz, G.; Gottesman, S. Global analysis of small RNA and mRNA targets of Hfq. Mol. Microbiol. 2003, 50, 1111–1124. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Rosner, J.L.; Martin, R.G.; Solé, M.; Vila, J. SoxS-dependent coregulation of ompN and ydbK in a multidrug-resistant Escherichia coli strain. FEMS Microbiol. Lett. 2012, 332, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Raivio, T.L.; Silhavy, T.J. Periplasmic stress and ECF sigma factors. Annu. Rev. Microbiol. 2001, 55, 591–624. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Pfeiffer, V.; Mika, F.; Lucchini, S.; Hinton, J.C.; Vogel, J. SigmaE-dependent small RNAs of Salmonella respond to membrane stress by accelerating global omp mRNA decay. Mol. Microbiol. 2006, 62, 1674–1688. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Pagès, J.-M.; Villard, C.; Pradel, E. The eefABC multidrug efflux pump operon is repressed by H-NS in Enterobacter aerogenes. J. Bacteriol. 2005, 187, 3894–3897. [Google Scholar] [CrossRef] [PubMed]
- Casadaban, M.J. Transposition and fusion of the lac genes to selected promoters in Escherichia coli using bacteriophage lambda and Mu. J. Mol. Biol. 1976, 104, 541–555. [Google Scholar] [CrossRef]
- Sarma, V.; Reeves, P. Genetic locus (ompB) affecting a major outer-membrane protein in Escherichia coli K-12. J. Bacteriol. 1977, 132, 23–27. [Google Scholar] [PubMed]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [PubMed]
- Klein, G.; Lindner, B.; Brade, H.; Raina, S. Molecular basis of lipopolysaccharide heterogeneity in Escherichia coli: Envelope stress-responsive regulators control the incorporation of glycoforms with a third 3-deoxy-α-d-manno-oct-2-ulosonic acid and rhamnose. J. Biol. Chem. 2011, 286, 42787–42807. [Google Scholar] [CrossRef] [PubMed]
- Bertin, P.; Terao, E.; Lee, E.H.; Lejeune, P.; Colson, C.; Danchin, A.; Collatz, E. The H-NS protein is involved in the biogenesis of flagella in Escherichia coli. J. Bacteriol. 1994, 176, 5537–5540. [Google Scholar] [CrossRef] [PubMed]
- George, A.M.; Levy, S.B. Amplifiable resistance to tetracycline, chloramphenicol, and other antibiotics in Escherichia coli: Involvement of a non-plasmid-determined efflux of tetracycline. J. Bacteriol. 1983, 155, 531–540. [Google Scholar] [PubMed]
- Rosner, J.L.; Slonczewski, J.L. Dual regulation of inaA by the multiple antibiotic resistance (mar) and superoxide (soxRS) stress response systems of Escherichia coli. J. Bacteriol. 1994, 176, 6262–6269. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Simonet, V.; Malle, M.; Fourel, D.; Bolla, J.-M.; Pages, J.-M. Crucial domains are conserved in Enterobacteriaceae porins. FEMS Microbiol. Lett. 1996, 136, 91–97. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Description | Source or Reference |
|---|---|---|
| E. coli strains | ||
| MC4100 | F− [araD139]B/r Δ(argF-lac)169 λ− e14 flhD5301 Δ(fruK-yeiR)725(fruA25) relA1 rpsL150(StrR) rbsR22 Δ(fimB-fimE)632(::IS1) deoC1 | [60] |
| MH1160 | MC4100 ompR101 | [61] |
| TR49 | MC4100 λRS88[degP–lacZ] | [47] |
| W3110 | F− λ− IN(rrnD-rrnE)1 rph-1 | [62] |
| SR8265 | W3110 rybB< >aph, KanR, source for P1 transduction | [63] |
| PS2209 | W3110 ΔlacZ169 | [64] |
| PS2652 | ΔlacZ169 zch-506::TnlO hns-1001::Tnseq1, KanR, source for P1 transduction | [64] |
| AG100 | F− glnX44(AS) galK2(Oc) rpsL704(StrR) xylA5 mtl-1 argE3(Oc) thiE1 tfr-3 | [65] |
| CH164 | AG100 marA zdd-230::Tn9, CamR, source for P1 transduction | [66] |
| BW25113 | F− Δ(araD–araB)567 ΔlacZ4787(::rrnB-3) λ− rph-1 Δ(rhaD–rhaB)568 hsdR514 | [67] |
| JW4130 | BW25113 hfq::kan, KanR, source for P1 transduction | GE Healthcare |
| SD01 | MC4100 ΔrybB | This study |
| SD02 | MC4100 marA zdd-230::Tn9, CamR, | This study |
| SD03 | MC4100 Δhfq | This study |
| SD04 | MC4100 Δhns | This study |
| SD05 | MC4100 ΔrpoS | This study |
| W3110ΔompF | W3110 ompF::kan | M.G. Page |
| W3110ΔompC | W3110 ompC::kan | M.G. Page |
| W3110ΔompFΔompC | W3110 ΔompFΔompC | M.G. Page |
| Plasmids | ||
| pDrive | PCR cloning vector; AmpR, KanR | Qiagen |
| pRC1 | pDrive containing Enterobacter aerogenes MarA | [31] |
| pRC2 | pDrive containing Enterobacter aerogenes RamA | [32] |
| pSD01 | pDrive encoding MicC sRNA | This study |
| pFus2K | Cloning vector with promoter-less lacZ, KanR | [59] |
| pSD02 | pFus2K containing the micC–lacZ fusion | This study |
| pSD03 | pFus2K containing the ompN–lacZ fusion | This study |
| pTrc99A | Expression vector with the inducible PTRC promoter, AmpR | Pharmacia |
| pSD04 | pTrc99A containing OmpN | This study |
| pSD05 | pTrc99A containing RseA | This study |
| pBAD24 | Expression vector with the inducible PBAD promoter, AmpR | [68] |
| pBAD24-NlpE | pBAD24 containing NlpE | M. Masi |
| pBAD33 | Expression vector with the inducible PBAD promoter, CamR | [68] |
| pBAD33-CpxA* | pBAD33 containing an autoactivated (*) CpxA | M. Masi |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dam, S.; Pagès, J.-M.; Masi, M. Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli. Antibiotics 2017, 6, 33. https://doi.org/10.3390/antibiotics6040033
Dam S, Pagès J-M, Masi M. Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli. Antibiotics. 2017; 6(4):33. https://doi.org/10.3390/antibiotics6040033
Chicago/Turabian StyleDam, Sushovan, Jean-Marie Pagès, and Muriel Masi. 2017. "Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli" Antibiotics 6, no. 4: 33. https://doi.org/10.3390/antibiotics6040033
APA StyleDam, S., Pagès, J. -M., & Masi, M. (2017). Dual Regulation of the Small RNA MicC and the Quiescent Porin OmpN in Response to Antibiotic Stress in Escherichia coli. Antibiotics, 6(4), 33. https://doi.org/10.3390/antibiotics6040033