Phenotypic and Genotypic Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) Related to Persistent Endovascular Infection


Abstract
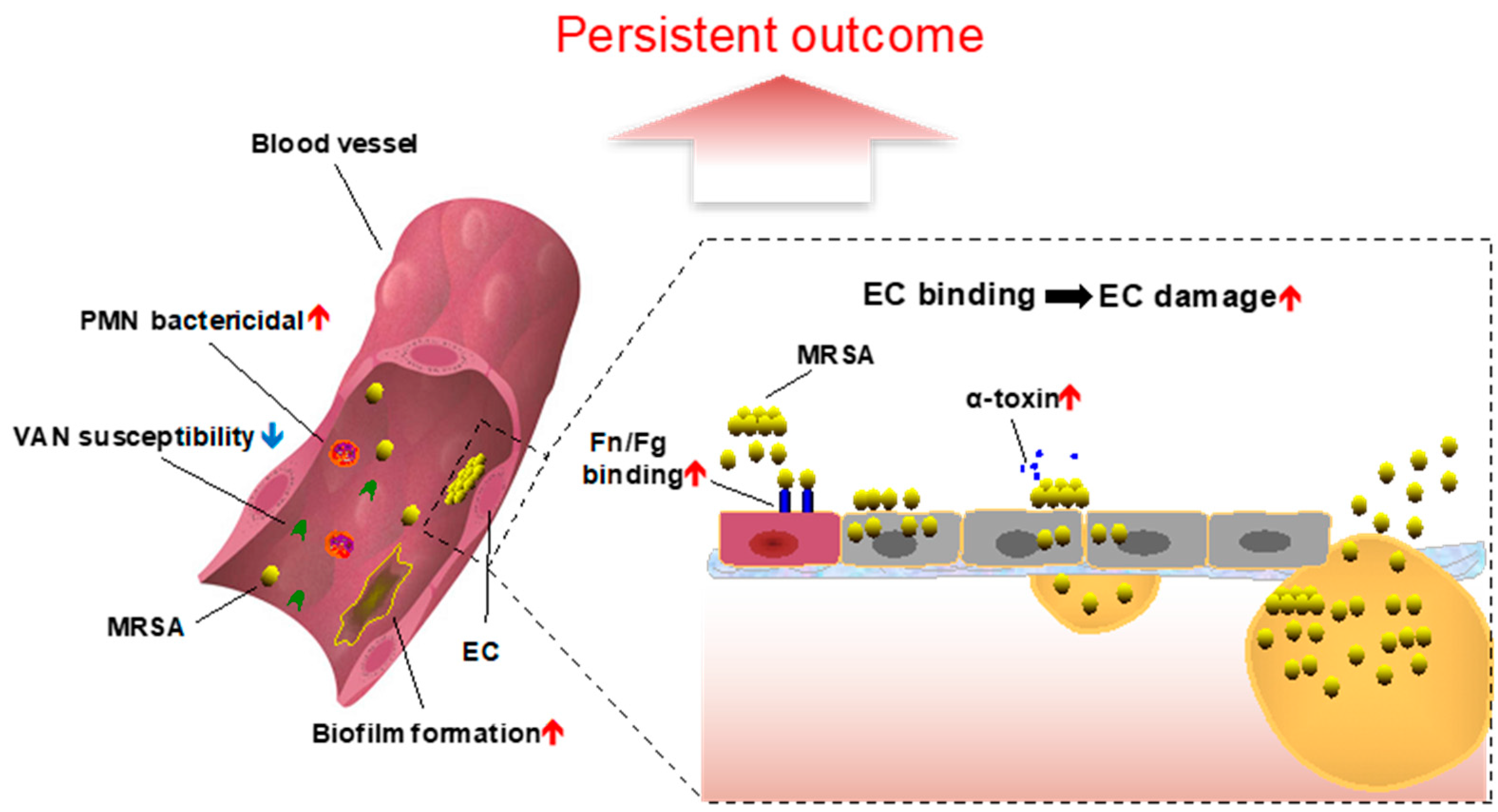
:1. Introduction
2. Genotypic Characteristics of MRSA Persistent Bacteremia
2.1. Standard Molecular Typing
2.2. Screening of Virulence Genes
2.3. Activation of Global Regulons
3. Phenotypic Characteristics of MRSA Persistent Bacteremia
3.1. Pathogen-Drug Interactions
3.1.1. VAN Susceptibility In Vitro vs. In Vivo
3.1.2. VAN-Killing Activity and Affinity for MRSA
3.1.3. MRSA Biofilm Formation
3.2. Pathogen-Host Interactions
3.2.1. Host Defense Peptide (HDP) Susceptibility
3.2.2. Survival in the Face of Polymorphonuclear Leukocytes (PMNs)
3.2.3. Adherence to Host Endothelial Cells and Substrates
3.2.4. Endothelial Cell Invasion and Damage
3.2.5. VAN Efficacy in IE Models
4. Treatment Strategies for Persistent MRSA Endovascular Infection
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Fowler, V.G., Jr.; Miro, J.M.; Hoen, B.; Cabell, C.H.; Abrutyn, E.; Rubinstein, E.; Corey, G.R.; Spelman, D.; Bradley, S.F.; Barsic, B.; et al. Staphylococcus aureus endocarditis: A consequence of medical progress. JAMA 2005, 293, 3012–3021. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, E. Staphylococcus aureus bacteremia with known sources. Int. J. Antimicrob. Agents 2008, 32, S18–S20. [Google Scholar] [CrossRef]
- Chang, F.Y.; Peacock, J.E., Jr.; Musher, D.M.; Triplett, P.; MacDonald, B.B.; Mylotte, J.M.; O’Donnell, A.; Wagener, M.M.; Yu, V.L. Staphylococcus aureus bacteremia: Recurrence and the impact of antibiotic treatment in a prospective multicenter study. Medicine 2003, 82, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Menichetti, F. Current and emerging serious Gram-positive infections. Clin. Microbiol. Infect. 2005, 11, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Q.; Fowler, V.G.; Yeaman, M.R.; Perdreau-Remington, F.; Kreiswirth, B.N.; Bayer, A.S. Phenotypic and genotypic characteristics of persistent methicillin-resistant Staphylococcus aureus bacteremia in vitro and in an experimental endocarditis model. J. Infect. Dis. 2009, 199, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.; Huang, J.; Jin, N.; Noskin, G.A.; Zembower, T.R.; Bolon, M. Persistent Staphylococcus aureus bacteremia: An analysis of risk factors and outcomes. Arch. Intern. Med. 2007, 167, 1861–1867. [Google Scholar] [CrossRef]
- Fowler, V.G., Jr.; Sakoulas, G.; McIntyre, L.M.; Meka, V.G.; Arbeit, R.D.; Cabell, C.H.; Stryjewski, M.E.; Eliopoulos, G.M.; Reller, L.B.; Corey, G.R.; et al. Persistent bacteremia due to methicillin-resistant Staphylococcus aureus infection is associated with agr dysfunction and low-level in vitro resistance to thrombin-induced platelet microbicidal protein. J. Infect. Dis. 2004, 190, 1140–1149. [Google Scholar] [CrossRef]
- Khatib, R.; Johnson, L.B.; Fakih, M.G.; Riederer, K.; Khosrovaneh, A.; Shamse Tabriz, M.; Sharma, M.; Saeed, S. Persistence in Staphylococcus aureus bacteremia: incidence, characteristics of patients and outcome. Scand. J. Infect. Dis. 2006, 38, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Bayer, A.S.; Fowler, V.G., Jr.; McKinnell, J.A.; Abdel Hady, W.; Sakoulas, G.; Yeaman, M.R.; Xiong, Y.Q. Combinatorial phenotypic signatures distinguish persistent from resolving methicillin-resistant Staphylococcus aureus bacteremia isolates. Antimicrob. Agents Chemother. 2011, 55, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Chen, L.; Bayer, A.S.; Hady, W.A.; Kreiswirth, B.N.; Xiong, Y.Q. Relationship of agr expression and function with virulence and vancomycin treatment outcomes in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 5631–5639. [Google Scholar] [CrossRef]
- Fowler, V.G., Jr.; Nelson, C.L.; McIntyre, L.M.; Kreiswirth, B.N.; Monk, A.; Archer, G.L.; Federspiel, J.; Naidich, S.; Remortel, B.; Rude, T.; et al. Potential associations between hematogenous complications and bacterial genotype in Staphylococcus aureus infection. J. Infect. Dis. 2007, 196, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Lalani, T.; Federspiel, J.J.; Boucher, H.W.; Rude, T.H.; Bae, I.G.; Rybak, M.J.; Tonthat, G.T.; Corey, G.R.; Stryjewski, M.E.; Sakoulas, G.; et al. Associations between the genotypes of Staphylococcus aureus bloodstream isolates and clinical characteristics and outcomes of bacteremic patients. J. Clin. Microbiol. 2008, 46, 2890–2896. [Google Scholar] [CrossRef]
- Abdelhady, W.; Chen, L.; Bayer, A.S.; Seidl, K.; Yeaman, M.R.; Kreiswirth, B.N.; Xiong, Y.Q. Early agr activation correlates with vancomycin treatment failure in multi-clonotype MRSA endovascular infections. J. Antimicrob. Chemother. 2015, 70, 1443–1452. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [Green Version]
- Feil, E.J.; Li, B.C.; Aanensen, D.M.; Hanage, W.P.; Spratt, B.G. eBURST: Inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 2004, 186, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- Nienaber, J.J.; Sharma Kuinkel, B.K.; Clarke-Pearson, M.; Lamlertthon, S.; Park, L.; Rude, T.H.; Barriere, S.; Woods, C.W.; Chu, V.H.; Marin, M.; et al. Methicillin-susceptible Staphylococcus aureus endocarditis isolates are associated with clonal complex 30 genotype and a distinct repertoire of enterotoxins and adhesins. J. Infect. Dis. 2011, 204, 704–713. [Google Scholar] [CrossRef]
- Shopsin, B.; Gomez, M.; Montgomery, S.O.; Smith, D.H.; Waddington, M.; Dodge, D.E.; Bost, D.A.; Riehman, M.; Naidich, S.; Kreiswirth, B.N. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 1999, 37, 3556–3563. [Google Scholar] [PubMed]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef]
- Dunman, P.M.; Murphy, E.; Haney, S.; Palacios, D.; Tucker-Kellogg, G.; Wu, S.; Brown, E.L.; Zagursky, R.J.; Shlaes, D.; Projan, S.J. Transcription profiling-based identification of Staphylococcus aureus genes regulated by the agr and/or sarA Loci. J. Bacteriol. 2001, 183, 7341–7353. [Google Scholar] [CrossRef]
- Francois, P.; Koessler, T.; Huyghe, A.; Harbarth, S.; Bento, M.; Lew, D.; Etienne, J.; Pittet, D.; Schrenzel, J. Rapid Staphylococcus aureus agr type determination by a novel multiplex real-time quantitative PCR assay. J. Clin. Microbiol. 2006, 44, 1892–1895. [Google Scholar] [CrossRef] [PubMed]
- Moise-Broder, P.A.; Sakoulas, G.; Eliopoulos, G.M.; Schentag, J.J.; Forrest, A.; Moellering, R.C., Jr. Accessory gene regulator group II polymorphism in methicillin-resistant Staphylococcus aureus is predictive of failure of vancomycin therapy. Clin. Infect. Dis. 2004, 38, 1700–1705. [Google Scholar] [CrossRef]
- Hernandez, D.; Seidl, K.; Corvaglia, A.R.; Bayer, A.S.; Xiong, Y.Q.; Francois, P. Genome sequences of sequence type 45 (ST45) persistent methicillin-resistant Staphylococcus aureus (MRSA) bacteremia strain 300-169 and ST45 resolving MRSA bacteremia strain 301-188. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Abdelhady, W.; Donegan, N.P.; Seidl, K.; Cheung, A.; Zhou, Y.F.; Yeaman, M.R.; Bayer, A.S.; Xiong, Y.Q. Role of purine biosynthesis in persistent methicillin-resistant Staphylococcus aureus (MRSA) infection. J. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Q.; Willard, J.; Yeaman, M.R.; Cheung, A.L.; Bayer, A.S. Regulation of Staphylococcus aureus alpha-toxin gene (hla) expression by agr, sarA, and sae in vitro and in experimental infective endocarditis. J. Infect. Dis. 2006, 194, 1267–1275. [Google Scholar] [CrossRef]
- Abdelhady, W.; Bayer, A.S.; Seidl, K.; Nast, C.C.; Kiedrowski, M.R.; Horswill, A.R.; Yeaman, M.R.; Xiong, Y.Q. Reduced vancomycin susceptibility in an in vitro catheter-related biofilm model correlates with poor therapeutic outcomes in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.S.; Prasad, R.; Chandra, J.; Koul, A.; Smriti, M.; Varma, A.; Skurray, R.A.; Firth, N.; Brown, M.H.; Koo, S.P.; et al. In vitro resistance of Staphylococcus aureus to thrombin-induced platelet microbicidal protein is associated with alterations in cytoplasmic membrane fluidity. Infect. Immun. 2000, 68, 3548–3553. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Q.; Sharma-Kuinkel, B.K.; Casillas-Ituarte, N.N.; Fowler, V.G., Jr.; Rude, T.; DiBartola, A.C.; Lins, R.D.; Abdel-Hady, W.; Lower, S.K.; Bayer, A.S. Endovascular infections caused by methicillin-resistant Staphylococcus aureus are linked to clonal complex-specific alterations in binding and invasion domains of fibronectin-binding protein A as well as the occurrence of fnbB. Infect. Immun. 2015, 83, 4772–4780. [Google Scholar] [CrossRef] [PubMed]
- Seidl, K.; Bayer, A.S.; McKinnell, J.A.; Ellison, S.; Filler, S.G.; Xiong, Y.Q. In vitro endothelial cell damage is positively correlated with enhanced virulence and poor vancomycin responsiveness in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Cell Microbiol. 2011, 13, 1530–1541. [Google Scholar] [CrossRef]
- Bae, T.; Baba, T.; Hiramatsu, K.; Schneewind, O. Prophages of Staphylococcus aureus Newman and their contribution to virulence. Mol. Microbiol. 2006, 62, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Gonzalez, S.; Quiles-Puchalt, N.; Gutierrez, D.; Penades, J.R.; Garcia, P.; Rodriguez, A. Lysogenization of Staphylococcus aureus RN450 by phages varphi11 and varphi80alpha leads to the activation of the SigB regulon. Sci. Rep. 2018, 8, 12662. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yeo, W.S.; Bae, T. The SaeRS Two-component system of Staphylococcus aureus. Genes 2016, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pfortner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PloS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of Biochemistry: Life at the Molecular Level, 3rd ed.; Wiley: Hoboken, NJ, USA, 2008. [Google Scholar]
- Yamaoka, T.; Kondo, M.; Honda, S.; Iwahana, H.; Moritani, M.; Ii, S.; Yoshimoto, K.; Itakura, M. Amidophosphoribosyltransferase limits the rate of cell growth-linked de novo purine biosynthesis in the presence of constant capacity of salvage purine biosynthesis. J. Biol. Chem. 1997, 272, 17719–17725. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Entenza, J.M.; Giachino, P. Influence of a functional sigB operon on the global regulators sar and agr in Staphylococcus aureus. J. Bacteriol. 2001, 183, 5171–5179. [Google Scholar] [CrossRef]
- Steinhuber, A.; Goerke, C.; Bayer, M.G.; Doring, G.; Wolz, C. Molecular architecture of the regulatory locus sae of Staphylococcus aureus and its impact on expression of virulence factors. J. Bacteriol. 2003, 185, 6278–6286. [Google Scholar] [CrossRef]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef]
- Lemaire, S.; Kosowska-Shick, K.; Julian, K.; Tulkens, P.M.; Van Bambeke, F.; Appelbaum, P.C. Activities of antistaphylococcal antibiotics towards the extracellular and intraphagocytic forms of Staphylococcus aureus isolates from a patient with persistent bacteraemia and endocarditis. Clin. Microbiol. Infect. 2008, 14, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.; Stinear, T.P.; Grayson, M.L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef]
- Rose, W.E.; Knier, R.M.; Hutson, P.R. Pharmacodynamic effect of clinical vancomycin exposures on cell wall thickness in heterogeneous vancomycin-intermediate Staphylococcus aureus. J. Antimicrob. Chemother. 2010, 65, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 207–228. [Google Scholar] [PubMed]
- Abdelhady, W.; Bayer, A.S.; Seidl, K.; Moormeier, D.E.; Bayles, K.W.; Cheung, A.; Yeaman, M.R.; Xiong, Y.Q. Impact of vancomycin on sarA-mediated biofilm formation: role in persistent endovascular infections due to methicillin-resistant Staphylococcus aureus. J. Infect. Dis. 2014, 209, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Bayer, A.S.; Koo, S.P.; Foss, W.; Sullam, P.M. Platelet microbicidal proteins and neutrophil defensin disrupt the Staphylococcus aureus cytoplasmic membrane by distinct mechanisms of action. J. Clin. Investig. 1998, 101, 178–187. [Google Scholar] [CrossRef]
- Cole, J.N.; Nizet, V. Bacterial evasion of host antimicrobial peptide defenses. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Dhawan, V.K.; Yeaman, M.R.; Bayer, A.S. Influence of in vitro susceptibility phenotype against thrombin-induced platelet microbicidal protein on treatment and prophylaxis outcomes of experimental Staphylococcus aureus endocarditis. J. Infect. Dis. 1999, 180, 1561–1568. [Google Scholar] [CrossRef]
- Peschel, A.; Jack, R.W.; Otto, M.; Collins, L.V.; Staubitz, P.; Nicholson, G.; Kalbacher, H.; Nieuwenhuizen, W.F.; Jung, G.; Tarkowski, A.; et al. Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor MprF is based on modification of membrane lipids with l-lysine. J. Exp. Med. 2001, 193, 1067–1076. [Google Scholar] [CrossRef]
- Richards, R.L.; Haigh, R.D.; Pascoe, B.; Sheppard, S.K.; Price, F.; Jenkins, D.; Rajakumar, K.; Morrissey, J.A. Persistent Staphylococcus aureus isolates from two independent cases of bacteremia display increased bacterial fitness and novel immune evasion phenotypes. Infect. Immun. 2015, 83, 3311–3324. [Google Scholar] [CrossRef]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Geiger, T.; Francois, P.; Liebeke, M.; Fraunholz, M.; Goerke, C.; Krismer, B.; Schrenzel, J.; Lalk, M.; Wolz, C. The stringent response of Staphylococcus aureus and its impact on survival after phagocytosis through the induction of intracellular PSMs expression. PLoS Pathog. 2012, 8, e1003016. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.; Joo, H.S.; Chatterjee, S.S.; Otto, M. Phenol-soluble modulins--critical determinants of staphylococcal virulence. FEMS Microbiol. Rev. 2014, 38, 698–719. [Google Scholar] [CrossRef]
- Chorianopoulos, E.; Bea, F.; Katus, H.A.; Frey, N. The role of endothelial cell biology in endocarditis. Cell Tissue Res. 2009, 335, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Piroth, L.; Que, Y.A.; Widmer, E.; Panchaud, A.; Piu, S.; Entenza, J.M.; Moreillon, P. The fibrinogen- and fibronectin-binding domains of Staphylococcus aureus fibronectin-binding protein A synergistically promote endothelial invasion and experimental endocarditis. Infect. Immun. 2008, 76, 3824–3831. [Google Scholar] [CrossRef]
- Proctor, R.A.; Balwit, J.M.; Vesga, O. Variant subpopulations of Staphylococcus aureus as cause of persistent and recurrent infections. Infect. Agents Dis. 1994, 3, 302–312. [Google Scholar]
- Zhang, Y. Persisters, persistent infections and the Yin-Yang model. Emerg. Microbes. Infect. 2014, 3, e3. [Google Scholar] [CrossRef] [PubMed]
- Garzoni, C.; Kelley, W.L. Staphylococcus aureus: New evidence for intracellular persistence. Trends Microbiol. 2009, 17, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34, table of contents. [Google Scholar] [CrossRef] [PubMed]
- Kullar, R.; Sakoulas, G.; Deresinski, S.; van Hal, S.J. When sepsis persists: A review of MRSA bacteraemia salvage therapy. J. Antimicrob. Chemother. 2016, 71, 576–586. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, S.H.; Kim, M.J.; Lee, Y.M.; Park, S.Y.; Moon, S.M.; Park, K.H.; Chong, Y.P.; Lee, S.O.; Choi, S.H.; et al. Efficacy of linezolid-based salvage therapy compared with glycopeptide-based therapy in patients with persistent methicillin-resistant Staphylococcus aureus bacteremia. J. Infect. 2012, 65, 505–512. [Google Scholar] [CrossRef]
- Ruggero, M.A.; Peaper, D.R.; Topal, J.E. Telavancin for refractory methicillin-resistant Staphylococcus aureus bacteremia and infective endocarditis. Infect. Dis. 2015, 47, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Barber, K.E.; Smith, J.R.; Ireland, C.E.; Boles, B.R.; Rose, W.E.; Rybak, M.J. Evaluation of ceftaroline alone and in combination against biofilm-producing methicillin-resistant Staphylococcus aureus with reduced susceptibility to daptomycin and vancomycin in an in vitro pharmacokinetic/pharmacodynamic model. Antimicrob. Agents Chemother. 2015, 59, 4497–4503. [Google Scholar] [CrossRef] [PubMed]
- Trotonda, M.P.; Manna, A.C.; Cheung, A.L.; Lasa, I.; Penades, J.R. SarA positively controls bap-dependent biofilm formation in Staphylococcus aureus. J. Bacteriol. 2005, 187, 5790–5798. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cheung, A.; Bayer, A.S.; Chen, L.; Abdelhady, W.; Kreiswirth, B.N.; Yeaman, M.R.; Xiong, Y.Q. The global regulon sarA regulates beta-lactam antibiotic resistance in methicillin-resistant Staphylococcus aureus in vitro and in endovascular infections. J. Infect. Dis. 2016, 214, 1421–1429. [Google Scholar] [CrossRef]
- Arya, R.; Ravikumar, R.; Santhosh, R.S.; Princy, S.A. SarA based novel therapeutic candidate against Staphylococcus aureus associated with vascular graft infections. Front. Microbiol. 2015, 6, 416. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Characteristic | PB Isolates (Frequency) | RB Isolates (Frequency) | References |
|---|---|---|---|
| Genotypic | |||
| Identical PFGE cluster | 85% | 44% | [9] |
| MLST | CC5 (53%), CC30 (21%–48%) | CC30 (18%) | [7,13,18] |
| spa type | 16 (21%) | 16 (18%) | [7,13] |
| SCCmec type | II (21%) | II (18%) | [7,13] |
| agr type | II (68.9%), III (21%) | II (27.8%), III (18%) | [7,23] |
| Screen of virulence genes | cna (21%), cap8 (21%), tst-1 (21%) | sdrD (47%), sdrE (47%), cap5 (21%), pvl (56%) | [7,14,18] |
| Prophage | 2 | 1 | [24] |
| Early on-set activation of global regulons/genes in PB vs. RB | agr RNAIII, sigB, sarA, sae, cap5 | [15,25,26] | |
| Higher gene expression in PB vs. RB | Purine-, amino acid-, carbohydrates- biosynthesis pathway, psms | [25] unpublished | |
| Lower gene expression in PB vs. RB | Histidine biosynthesis pathway genes | [25] | |
| Phenotypic | |||
| Pathogen-Drug interactions (PB vs. RB) | |||
| Survival with VAN exposure | > | [27] | |
| VAN binding | < | [27] | |
| In vitro growth rate | > | [25] | |
| Biofilm formation ± sub-MIC VAN (under static conditions) | > | [27] | |
| Biofilm formation (under flow conditions) | > | [27] | |
| Pathogen-Host interactions (PB vs. RB) | |||
| Survival with HDPs exposure | > | [7,11] | |
| Membrane fluidity | > | [7,28] | |
| Survival with PMNs exposure | > | unpublished | |
| Adherence to Fn and Fg | > | [29] | |
| Fn-FnBPs binding | > | [29] | |
| EC damage | > | [30] | |
| α-, δ-hemolysin | > | [9,12] | |
| VAN effectiveness in an IE model | < | [7,12,15] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yeaman, M.R.; Bayer, A.S.; Xiong, Y.Q. Phenotypic and Genotypic Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) Related to Persistent Endovascular Infection. Antibiotics 2019, 8, 71. https://doi.org/10.3390/antibiotics8020071
Li L, Yeaman MR, Bayer AS, Xiong YQ. Phenotypic and Genotypic Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) Related to Persistent Endovascular Infection. Antibiotics. 2019; 8(2):71. https://doi.org/10.3390/antibiotics8020071
Chicago/Turabian StyleLi, Liang, Michael R. Yeaman, Arnold S. Bayer, and Yan Q. Xiong. 2019. "Phenotypic and Genotypic Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) Related to Persistent Endovascular Infection" Antibiotics 8, no. 2: 71. https://doi.org/10.3390/antibiotics8020071
APA StyleLi, L., Yeaman, M. R., Bayer, A. S., & Xiong, Y. Q. (2019). Phenotypic and Genotypic Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) Related to Persistent Endovascular Infection. Antibiotics, 8(2), 71. https://doi.org/10.3390/antibiotics8020071