Size and Flexibility Define the Inhibition of the H3N2 Influenza Endonuclease Enzyme by Calix[n]arenes

 ,
,  , , ,
, , , 

Abstract
:1. Introduction
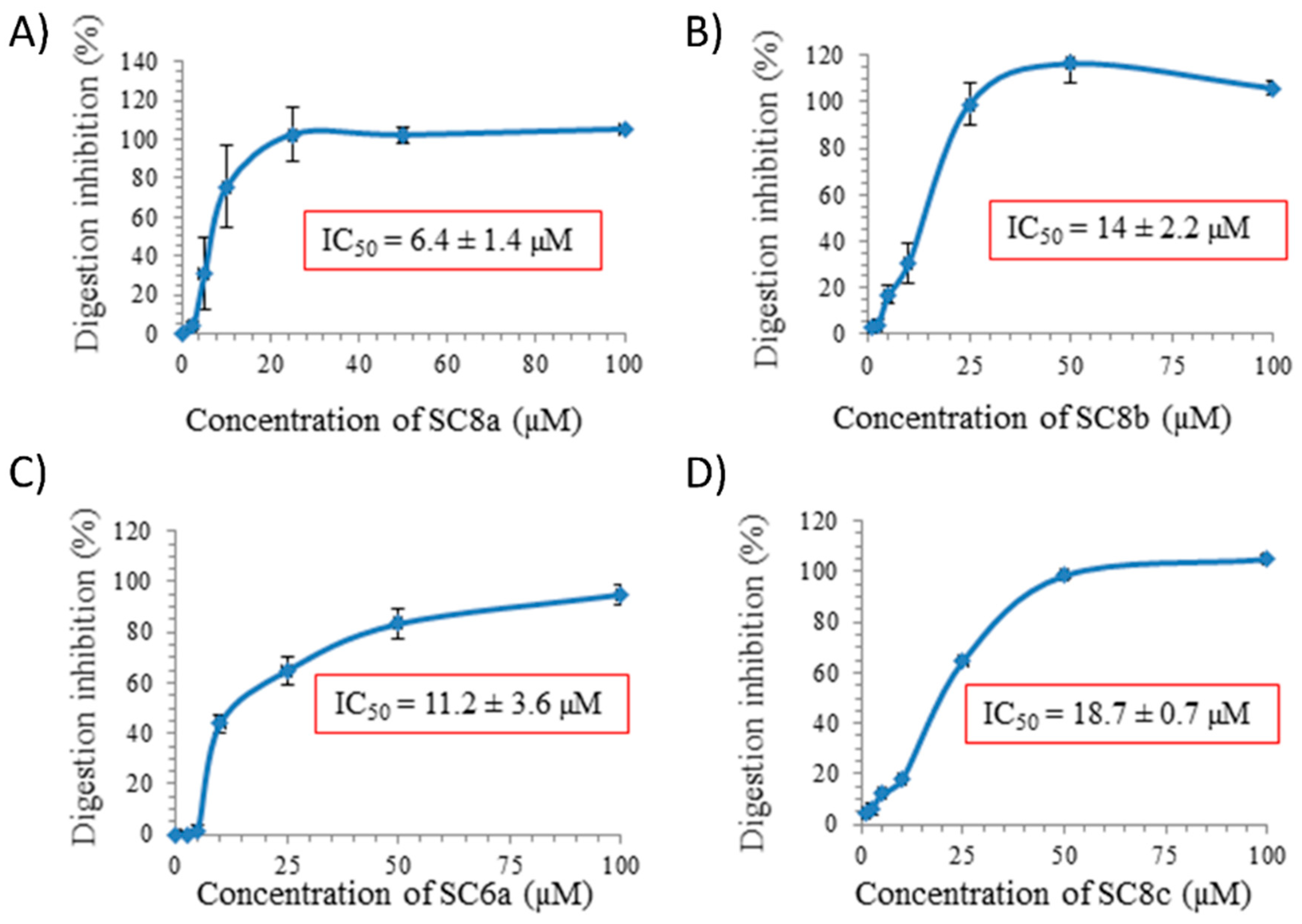
2. Results
3. Discussion
4. Materials and Methods
4.1. Synthesis and Characterization of Calix[n]arenes
4.2. Over-Expression and Purification of PA Endonuclease
4.3. PA Endonuclease Activity
4.4. PA Endonuclease Inhibition Assay for Macrocyclic Molecules
4.5. Molecular Modeling Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Virus Investigation Team; Dawood, F.S.; Jain, S.; Finelli, L.; Shaw, M.W.; Lindstrom, S.; Garten, R.J.; Gubareva, L.V.; Xu, X.; Bridges, C.B.; et al. Novel Swine-Origin Influenza A (H1N1). N. Engl. J. Med. 2009, 360, 2605–2615. [Google Scholar]
- Fraser, C.; Donnelly, C.; Cauchemez, S.; Hanage, W.; Van Kerkhove, M.; Hollingsworth, T.; Griffin, J.; Baggaley, R.; Jenkins, H.; Lyons, E.; et al. Pandemic Potential of a Strain of Influenza A (H1N1): Early Findings. Science 2009, 324, 1557–1561. [Google Scholar] [CrossRef] [Green Version]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018. [Google Scholar] [CrossRef] [PubMed]
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef]
- Rehwinkel, J.; Tan, C.P.; Goubau, D.; Schulz, O.; Pichlmair, A.; Bier, K.; Robb, N.; Vreede, F.; Barclay, W.; Fodor, E.; et al. RIG-I detects viral genomic RNA during negative-strand RNA virus infection. Cell 2010, 140, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Ernst, W.A.; Kim, H.J.; Tumpey, T.M.; Jansen, A.D.A.; Tai, W.; Cramer, D.V.; Adler-Moore, J.P.; Fujii, G. Protection against H1, H5, H6 and H9 influenza A infection with liposomal matrix 2 epitope vaccines. Vaccine 2006, 24, 5158–5168. [Google Scholar] [CrossRef] [PubMed]
- Deyde, V.M.; Xu, X.; Bright, R.A.; Shaw, M.; Smith, C.B.; Zhang, Y.; Shu, Y.; Gubareva, L.V.; Cox, N.J.; Klimov, A.I. Surveillance of resistance to adamantanes among influenza A(H3N2) and A(H1N1) viruses isolated worldwide. J. Infect. Dis. 2007, 196, 249–257. [Google Scholar] [CrossRef]
- Schaduangrat, N.; Phanich, J.; Rungrotmongkol, T.; Lerdsamran, H.; Puthavathana, P.; Ubol, S. The significance of naturally occurring neuraminidase quasispecies of H5N1 avian influenza virus on resistance to oseltamivir: A point of concern. J. Gen. Virol. 2016, 97, 1311–1323. [Google Scholar] [CrossRef]
- Dubois, R.M.; Slavish, P.J.; Baughman, B.M.; Yun, M.K.; Bao, J.; Webby, R.J.; Webb, T.R.; White, S.W. Structural and biochemical basis for development of influenza virus inhibitors targeting the PA endonuclease. PLoS Pathog. 2012, 8, e1002830. [Google Scholar] [CrossRef]
- Ju, H.; Zhang, J.; Huang, B.; Kang, D.; Huang, B.; Liu, X.; Zhan, P. Inhibitors of influenza virus polymerase acidic (PA) Endonuclease: Contemporary developments and perspectives. J. Med. Chem. 2017, 60, 3533–3551. [Google Scholar] [CrossRef]
- Yuan, S.; Wen, L.; Zhou, J. Inhibitors of Influenza A Virus Polymerase. ACS Infect. Dis. 2018, 4, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.; Reina, N. Baloxavir marboxil: Un potente inhibidor de la endonucleasa cap-dependiente de los virus gripales. Rev. Esp. Quimioter. 2019, 32, 1–5. [Google Scholar] [PubMed]
- Hayden, F.G.; Sugaya, N.; Hirotsu, N.; Lee, N.; de Jong, M.D.; Hurt, A.C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Shaw, M.L. Baloxavir marboxil: The new influenza drug on the market. Curr. Opin. Virol. 2019, 35, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Shoji, M.; Takahashi, E.; Hatakeyama, D.; Iwai, Y.; Morita, Y.; Shirayama, R.; Echigo, N.; Kido, H.; Nakamura, S.; Mashino, T.; et al. Anti-Influenza Activity of C60 Fullerene Derivatives. PLoS ONE 2013, 8, e66337. [Google Scholar] [CrossRef]
- Tauran, Y.; Anjard, C.; Kim, B.J.; Rhimi, M.; Coleman, A.W. Large negatively charged organic host molecules as inhibitors of endonuclease enzymes. Chem. Comm. 2014, 50, 11404–11406. [Google Scholar] [CrossRef] [PubMed]
- Pingoud, A.; Jeltsch, A. Structure and function of type II restriction endonucleases. Nucl. Acids Res. 2001, 29, 3705–3727. [Google Scholar] [CrossRef]
- Hileman, R.E.; Fromm, J.R.; Weiler, J.M.; Linhardt, R.J. Glycosaminoglycan-protein interactions: Definition of consensus sites in glycosaminoglycan binding proteins. BioEssays 1998, 20, 156–167. [Google Scholar] [CrossRef]
- Govindaraju, A.; Cortez, J.D.; Reveal, B.; Christensen, S.M. Endonuclease domain of non-LTR retrotransposons: Loss-of-function mutants and modeling of the R2Bm endonuclease. Nucl. Acids Res. 2016, 44, 3276–3287. [Google Scholar] [CrossRef]
- Pingoud, A.; Wilson, G.G.; Wende, W. Type II restriction endonucleases—A historical perspective and more. Nucl. Acids. Res. 2014, 42, 7489–7527. [Google Scholar] [CrossRef]
- Da Silva, E.; Ficheux, D.; Coleman, A.W. Anti-thrombotic activity of water-soluble calix[n]arenes. J. Incl. Phen. Macro. Chem. 2005, 52, 201–206. [Google Scholar] [CrossRef]
- Moussa, A.; Coleman, A.W.; Shahgaldian, P.; Da Silva, E.; Martin, A.; Lazar, A.; Leclere, E.; Dupin, M.; Perron, H. Process for detecting PrP using a macrocyclic adjuvant ligand. FR Patent Application No 20060019311, 26 January 2006. [Google Scholar]
- Coleman, A.W.; Perret, F.; Cecillon, S.; Moussa, A.; Martin, A.; Dupin, M.; Perron, H. Enhanced detection of the pathogenic prion protein by its supramolecular association with para-sulfonato-calix[n]arene derivatives. New J. Chem. 2007, 31, 711–717. [Google Scholar] [CrossRef]
- Cecillon, S.; Coleman, A.W.; Eveno-Nobile, A.; Perron, H.; Rodrigue, M. Method for Detecting Aggregate-Forming Circulating Protein Forms and Agent for Capturing Formed Aggregates. U.S. Patent Application No. US8158441B2, 17 April 2012. [Google Scholar]
- Perret, F.; Lazar, A.N.; Coleman, A.W. Biochemistry of the para-sulfonato-calix[n]arenes. Chem. Commun. 2006, 23, 2425–2438. [Google Scholar] [CrossRef]
- Moriya, T.; Saito, K.; Kurita, H.; Matsumoto, K.; Otake, T.; Mori, H.; Morimoto, M.; Ueba, N.; Kunita, N. A new candidate for an anti-HIV-1 agent: Modified cyclodextrin sulfate (mCDS71). J. Med. Chem. 1993, 36, 1674–1677. [Google Scholar] [CrossRef] [PubMed]
- Perret, F.; Coleman, A.W. Biochemistry of anionic calixarenes. Chem. Commun. 2011, 47, 7303–7319. [Google Scholar] [CrossRef]
- Mallipeddi, P.L.; Kumar, G.; White, S.W.; Webb, T.R. Recent advances in computer-aided drug design as applied to anti-influenza drug discovery. Curr. Top. Med. Chem. 2014, 14, 1875–1889. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.W.; Bott, S.G.; David Morley, S.; Mitchell Means, C.; Robinson, K.D.; Zhang, H.; Atwood, J.A. Novel Layer Structure of Sodium Calix [4] arenesulfonate Complexes—A Class of Organic Clay Mimics? Angew. Chem. 1988, 27, 1361–1362. [Google Scholar] [CrossRef]
- Danylyuk, O.; Suwinska, K. Solid-state interactions of calixarenes with biorelevant molecules. Chem. Commun. 2009, 5799–5813. [Google Scholar] [CrossRef]
- Da Silva, E.; Memmi, L.; Coleman, A.W.; Rather, B.; Zaworotko, M.J. Synthesis and solid-state structures of mono-functionalised para-H-calix-[6]-arenes. J. Supramol. Chem. 2002, 1, 135–138. [Google Scholar] [CrossRef]
- Perret, F.; Bonnard, V.; Danylyuk, O.; Suwinska, K.; Coleman, A.W. Conformational extremes in the supramolecular assemblies of para-sulfonato-calix [8] arene. New J. Chem. 2006, 30, 987–990. [Google Scholar] [CrossRef]
- Danylyuk, O.; Butkiewicz, H.; Coleman, A.W.; Suwinska, K. Host-guest complexes of local anesthetics with cucurbit [6] uril and para-sulfonatocalix [8] arene in the solid state. J. Mol. Struct. 2017, 1150, 28–36. [Google Scholar] [CrossRef]
- Leśniewska, B.; Coleman, A.W.; Tauran, Y.; Perret, F.; Suwińska, K. Pseudopolymorphs—A variety of self-organization of para-sulfonato-calix [8] arene and phenanthroline in the solid state. CrystEngComm 2016, 18, 8858–8870. [Google Scholar] [CrossRef]
- Dias, A.; Bouvier, D.; Crepin, T.; McCarthy, A.A.; Hart, D.J.; Baudin, F.; Cusack, S.; Ruigrok, R.W.H. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. Nature 2009, 458, 914–918. [Google Scholar] [CrossRef]
- Hu, Y.; Musharrafieh, R.; Ma, C.; Zhang, J.; Smee, D.F.; DeGrado, W.F.; Wang, J. An M2-V27A channel blocker demonstrates potent in vitro and in vivo antiviral activities against amantadine-sensitive and -resistant influenza A viruses. Antivir. Res. 2017, 140, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraris, O.; Kessler, N.; Lina, B. Sensitivity of influenza viruses to zanamivir and oseltamivir: A study performed on viruses circulating in France prior to the introduction of neuraminidase inhibitors in clinical practice. Antivir. Res. 2005, 68, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Noshi, T.; Kitano, M.; Taniguchi, K.; Yamamoto, A.; Omoto, S.; Baba, K.; Hashimoto, T.; Ishida, K.; Kushima, Y.; Hattori, K.; et al. In Vitro characterization of baloxavir acid, a first-in-class cap-dependent endonuclease inhibitor of the influenza virus polymerase PA subunit. Antivir. Res. 2018, 160, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Credille, C.V.; Dick, B.L.; Morrison, C.N.; Stokes, R.W.; Adamek, R.N.; Wu, N.C.; Wilson, I.A.; Cohen, S.M. Structure-Activity Relationships in Metal-Binding Pharmacophores for Influenza Endonuclease. J. Med. Chem. 2018, 61, 10206–10217. [Google Scholar] [CrossRef] [PubMed]
- Selkti, M.; Coleman, A.W.; Nicolis, I.; Douteau-Guével, N.; Villain, F.; Tomas, A.; de Rango, C. The first example of a substrate spanning the calix [4] arene bilayer: The solid state complex of p-sulfonatocalix [4] arene with L-lysine. Chem. Commun. 2000, 161–162. [Google Scholar] [CrossRef]
- Lazar, A.; Da Silva, E.; Navaza, A.; Barbey, C.; Coleman, A.W. A new packing motif for para-sulfonatocalix [4] arene: The solid state structure of the para-sulfonatocalix [4] arene D-arginine complex. Chem. Commun. 2004, 2162–2163. [Google Scholar] [CrossRef]
- Yousaf, A.; Abd Hamid, S.; Bunnori, N.M.; Ishola, A.A. Applications of calixarenes in cancer chemotherapy: Facts and perspectives. Drug Des. Devel. Ther. 2015, 9, 2831–2838. [Google Scholar]
- Tauran, Y.; Coleman, A.W.; Perret, F.; Kim, B. Cellular and in Vivo Biological Activities of the Calix [n] arenes. Curr. Org. Chem. 2015, 19, 2250–2270. [Google Scholar] [CrossRef]
- Gutsche, C.D.; Lin, L.G. Calixarene 12: The Synthesis of Functionalized Calixarene. Tetrahedron. 1986, 42, 1633–1640. [Google Scholar] [CrossRef]
- Coleman, A.W.; Jebors, S.; Cecillon, S.; Perret, P.; Garin, D.; Marti-Battle, D.; Moulin, M. Toxicity and biodistribution of para-sulfonato-calix [4] arene in mice. New J. Chem. 2008, 32, 780–782. [Google Scholar] [CrossRef]
- Shinkai, S.; Arimura, T.; Arak, K.; Kawabata, H.J. Syntheses and aggregation properties of new water-soluble calix [n] arenes. J. Chem. Soc. Perkin Trans. 1989, 1, 2039–2045. [Google Scholar] [CrossRef]
- Hwang, K.M.; Qi, Y.M.; Liu, S.Y.; Lee, T.C.; Choy, W.; Chen, J. Antithrombotic Treatment with Calixarene Compounds. U.S. Patent Application No. US5409959A, 25 April 1995. [Google Scholar]
- Shinkai, S.; Mori, S.; Koreishi, H.; Tsubakiand, T.; Manabe, O. Hexasulfonated calix [6] arene derivatives: A new class of catalysts, surfactants, and host molecules. J. Am. Chem. Soc. 1986, 108, 2409–2416. [Google Scholar] [CrossRef] [PubMed]
- Kalchenko, V.I.; Rudkevich, D.M.; Markovskii, L.N. Reactive exophosphorylated benzo-crown esters. Zhurnal Obshchei Khimii 1990, 60, 2813–2814. [Google Scholar]
- Zahid, A.; Jamil, W.; Begum, R. Method Development and Validation of SDS-PAGE for Quality Control Testing of Pegylated Interferon Alpha-2a. IOSR J. Pharm. Biol. 2014, 9, 32–36. [Google Scholar] [CrossRef]
- Navarro-Fernandez, J.; Pérez-Sánchez, H.; Martinez-Martinez, I.; Meliciani, I.; Guerrero, J.A.; Vicente, V.; Corral, J.; Wenzel, W.J. In silico discovery of a compound with nanomolar affinity to antithrombin causing partial activation and increased heparin affinity. Med. Chem. 2012, 55, 6403–6412. [Google Scholar] [CrossRef]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Case, D.A.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Greene, D.; Homeyer, N.; et al. AMBER 2017; University of California: San Francisco, CA, USA, 2017. Available online: http://ambermd.org/doc12/Amber17.pdf (accessed on 31 May 2017).
- Stroganov, O.V.; Novikov, F.N.; Stroylov, V.S.; Kulkov, V.; Chilov, G.G. Lead finder: An approach to improve accuracy of protein-ligand docking, binding energy estimation, and virtual screening. J. Chem. Inf. Model. 2008, 48, 2371–2385. [Google Scholar] [CrossRef]
- Sánchez-Linares, I.; Pérez-Sánchez, H.; Cecilia, J.; García, J. High-throughput parallel blind virtual screening using BINDSURF. BMC Bioinf. 2012, 13, S13. [Google Scholar] [CrossRef]
- Zhao, Y.; Truhlar, D.G. Density functionals with broad applicability in chemistry. Acc. Chem. Res. 2008, 41, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-class functionals and 12 other functionals. Theor. Chem. Acc. 2008, 120, 215–241. [Google Scholar]
- Boys, S.; Bernardi, F. The calculation of small molecular interactions by the differences of separate total energies. Some procedures with reduced errors. Mol. Phys. 1970, 19, 553–566. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Janesko, B.G.; Hratchian, H.P.; Ortiz, J.V.; et al. Gaussian 09, Revision E.01, Fox, Gaussian, Inc.: Wallingford, CT, USA, 2009.
- Tapia-Abellán, A.; Angosto-Bazarra, D.; Martínez-Banaclocha, H.; de Torre-Minguela, C.; Cerón-Carrasco, J.P.; Pérez-Sánchez, H.; Arostegui, J.I.; Pelegrin, P. MCC950 closes the active conformation of NLRP3 to an inactive state. Nat. Chem. Biol. 2019, 15, 560–564. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Molecules | Inhibition Activity IC50 (μM) | Reference | Available Commercially (Trade Name) |
|---|---|---|---|---|
| M2 Channel | Amantadine | 15.7 | [36] | Yes (Flumadine) |
| Rimantadine | 10.8 | Yes (Symmetrel) | ||
| Neuraminidase | Zanamivir | 0.002 | [37] | Yes (Relenza) |
| Oseltamivir carboxylate | 0.00067 | Yes (Tamiflu) | ||
| PA endonuclease | Baloxavir Acid | 0.00023 | [38] | Yes (Xofluza) |
| 4-dioxo-4-phenylbutanoic acid | 21.3 | [10] | No | |
| 5-hydroxy-4-pyridone-3-carboxy acid | 3.23 | No | ||
| 5-hydroxypyrimidin-4(3H)-one | 0.15 | No | ||
| SC4a | >100 | No | ||
| SC6a | 11.2 ± 3.6 | |||
| SC8a | 6.4 ± 1.4 | |||
| SC4b | >100 | |||
| SC6b | >100 | |||
| SC8b | 14.0 ± 2.2 | |||
| SC4c | >100 | |||
| SC6c | >100 | |||
| SC8c | 18.7 ± 0.7 | |||
| C4diP | >100 | |||
| CD | >100 |
| Type of Interactions | Interacting Residues |
|---|---|
| Hydrophobic | TYR24, TYR111 |
| Pi Stacking | TYR24, HIS41 |
| Hydrogen Bonds | THR32, ASN33, LYS34, VAL103, ARG106, TYR111, GLU114, SER175, ALA179 |
| Salt Bridges | HIS41, LYS115, LYS118 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tauran, Y.; Cerón-Carrasco, J.P.; Rhimi, M.; Perret, F.; Kim, B.; Collard, D.; Coleman, A.W.; Pérez-Sánchez, H. Size and Flexibility Define the Inhibition of the H3N2 Influenza Endonuclease Enzyme by Calix[n]arenes. Antibiotics 2019, 8, 73. https://doi.org/10.3390/antibiotics8020073
Tauran Y, Cerón-Carrasco JP, Rhimi M, Perret F, Kim B, Collard D, Coleman AW, Pérez-Sánchez H. Size and Flexibility Define the Inhibition of the H3N2 Influenza Endonuclease Enzyme by Calix[n]arenes. Antibiotics. 2019; 8(2):73. https://doi.org/10.3390/antibiotics8020073
Chicago/Turabian StyleTauran, Yannick, José Pedro Cerón-Carrasco, Moez Rhimi, Florent Perret, Beomjoon Kim, Dominique Collard, Anthony W. Coleman, and Horacio Pérez-Sánchez. 2019. "Size and Flexibility Define the Inhibition of the H3N2 Influenza Endonuclease Enzyme by Calix[n]arenes" Antibiotics 8, no. 2: 73. https://doi.org/10.3390/antibiotics8020073
APA StyleTauran, Y., Cerón-Carrasco, J. P., Rhimi, M., Perret, F., Kim, B., Collard, D., Coleman, A. W., & Pérez-Sánchez, H. (2019). Size and Flexibility Define the Inhibition of the H3N2 Influenza Endonuclease Enzyme by Calix[n]arenes. Antibiotics, 8(2), 73. https://doi.org/10.3390/antibiotics8020073