Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella enterica Motility, Biofilm Formation, and Gene Expression






Abstract
:1. Introduction
2. Results and Discussion
2.1. Antimicrobial Activity of Short Chain Fatty Acids
2.2. Effect of Short Chain Fatty Acids in Salmonella enterica Motility
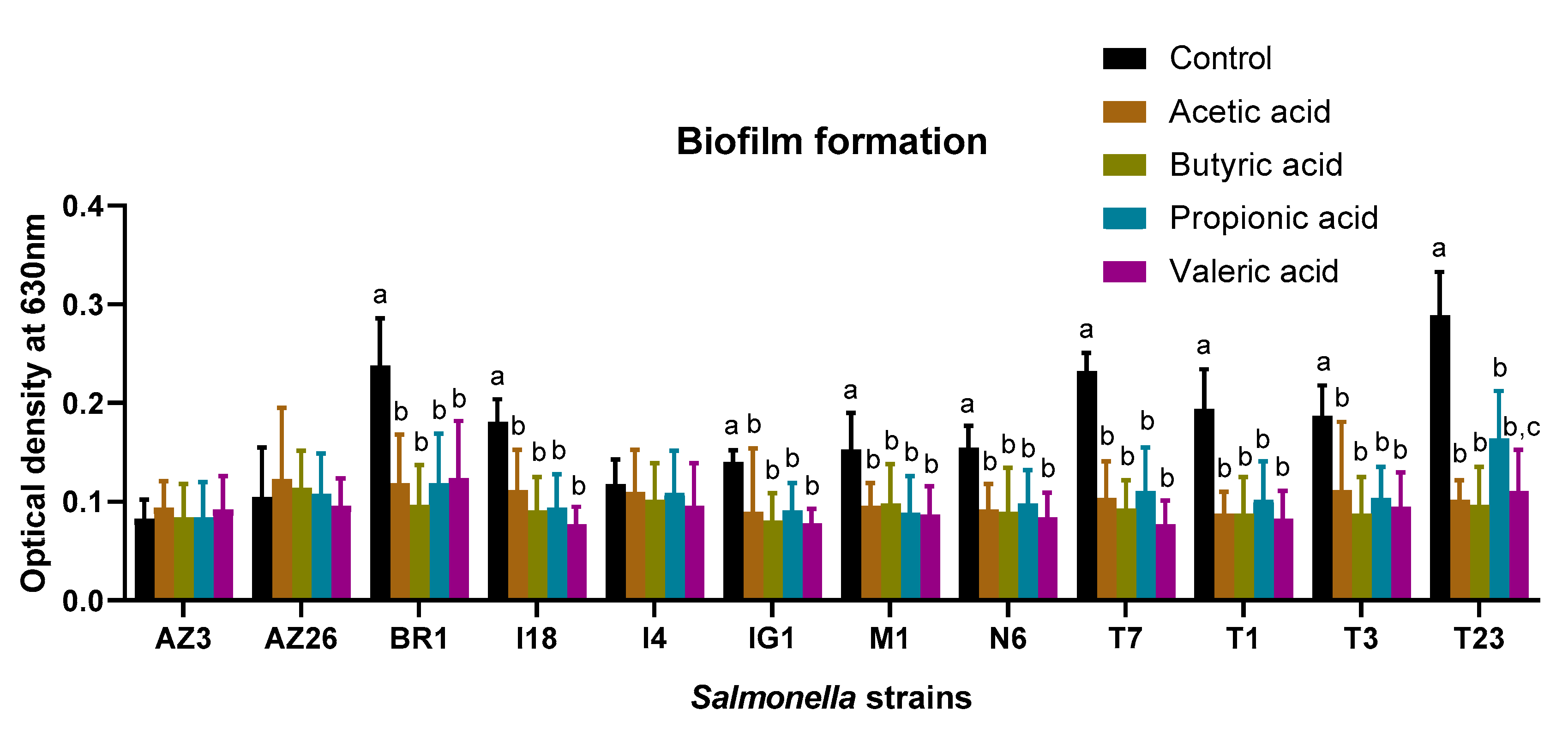
2.3. Effect of Short Chain Fatty Acids on Biofilm Formation
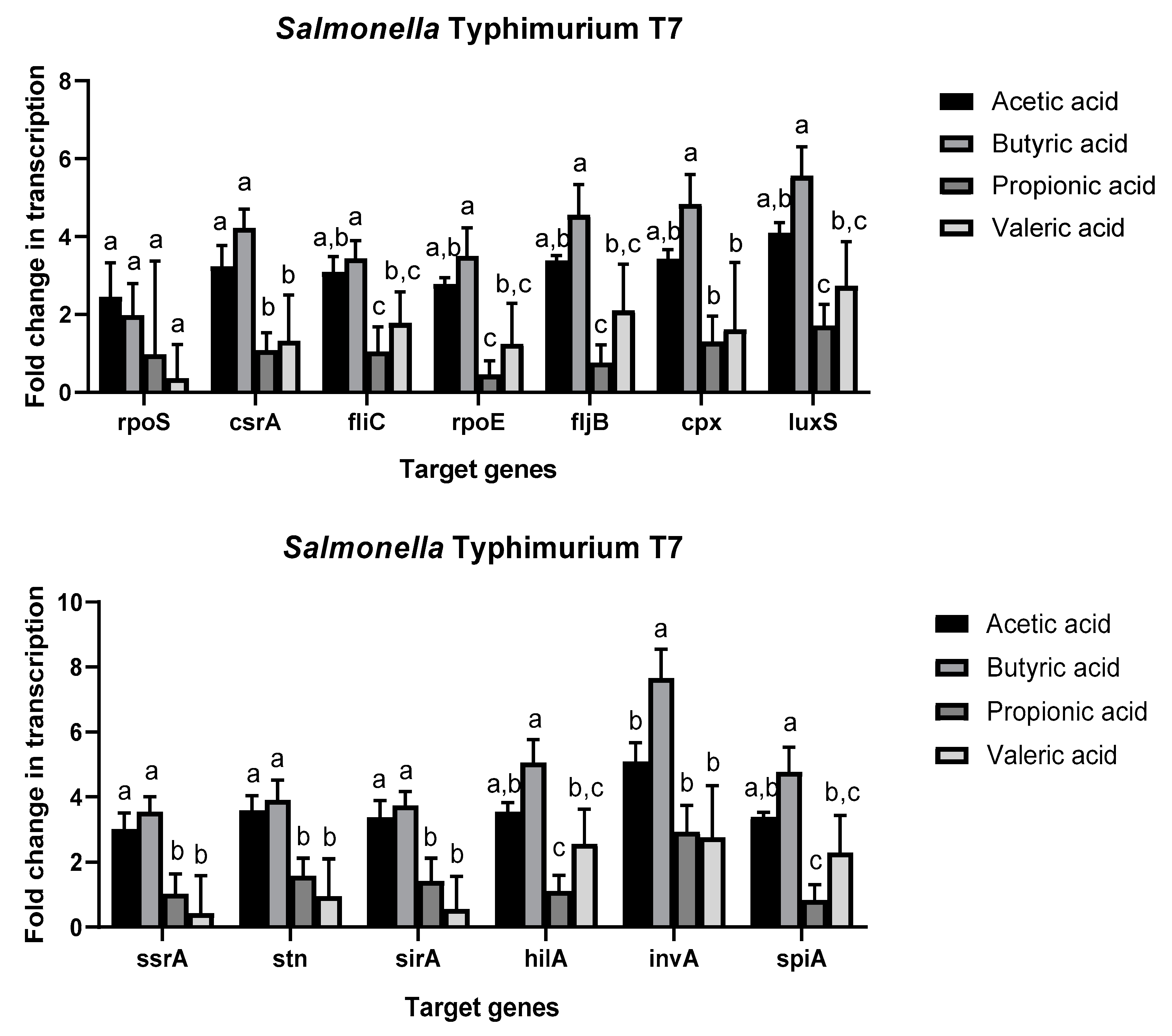
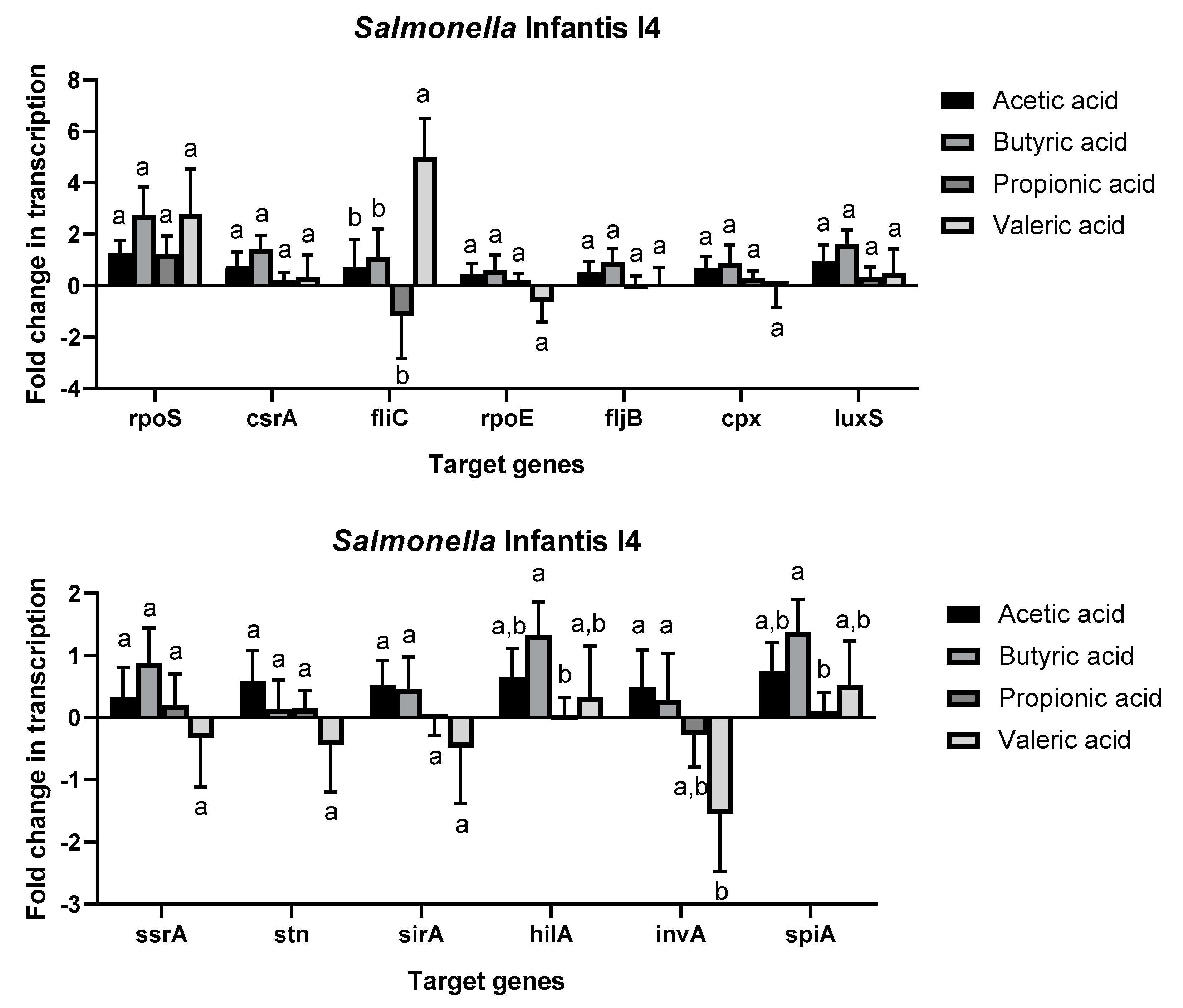
2.4. Effect of Short Chain Fatty Acids on Salmonella enterica Gene Expression
3. Materials and Methods
3.1. Salmonella enterica Strains and Short Chain Fatty Acids
3.2. Minimum Inhibitory Concentration and Minimum Biocidal Concentration of Short Chain Fatty Acids
3.3. Motility Assays
3.4. Biofilm Formation on Polystyrene
3.5. RNA Isolation and RT-qPCR
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, S.; Macfarlane, G.T. Session: Short-chain fatty acids. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.A.K.; Mullish, B.H.; Pechlivanis, A.; Liu, Z.; Brignardello, J.; Kao, D.; Holmes, E.; Li, J.V.; Clarke, T.B.; Thursz, M.R.; et al. Inhibiting Growth of Clostridioides difficile by Restoring Valerate, Produced by the Intestinal Microbiota. Gastroenterology 2018, 155, 1495–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Littman, D.R. The microbiome in infectious disease and inflammation. Ann. Rev. Immunol. 2012, 30, 759–795. [Google Scholar] [CrossRef] [Green Version]
- Ashida, H.; Ogawa, M.; Kim, M.; Mimuro, H.; Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nat. Chem. Biol. 2012, 8, 36–45. [Google Scholar] [CrossRef]
- Schulthess, J.; Pandey, S.; Capitani, M.; Rue-Albrecht, K.C.; Arnold, I.; Franchini, F.; Chomka, A.; Ilott, N.E.; Johnston, D.G.W.; Pires, E.; et al. The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity 2019, 50, 432–445. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, H.W.J.; Naylor, C.P.E.; MacFarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Repaske, D.R.; Adler, J. Change in intracellular pH of Escherichia coli mediates the chemotactic response to certain attractants and repellents. J. Bacteriol. 1981, 145, 1196–1208. [Google Scholar]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A Gut Commensal-Produced Metabolite Mediates Colonization Resistance to Salmonella Infection. Cell Host Microbe 2018, 24, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Rodríguez-Carrio, J.; Salazar, N.; Margolles, A.; González, S.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Suárez, A. Free fatty acids profiles are related to gut microbiota signatures and short-chain fatty acids. Front. Immunol. 2017, 8, 823. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Yao, Y.; Ju, S.Y. Short chain fatty acids and fecal microbiota abundance in humans with obesity: A systematic review and meta-analysis. Nutrients 2019, 11, 2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luethy, P.M.; Huynh, S.; Ribardo, D.A.; Winter, S.E.; Parker, C.T.; Hendrixson, D.R. Microbiota-derived short-chain fatty acids modulate expression of Campylobacter jejuni determinants required for commensalism and virulence. MBio 2017, 8, 00407-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, N.; Tashiro, K.; Kuhara, S.; Hayashi, T.; Sugimoto, N.; Tobe, T. Regulation of virulence by butyrate sensing in enterohaemorrhagic Escherichia coli. Microbiology 2009, 155, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.-C.; Garner, C.D.; Slauch, J.M.; Dwyer, Z.W.; Lawhon, S.D.; Frye, J.G.; Mcclelland, M.; Ahmer, B.M.M.; Altier, C. The intestinal fatty acid propionate inhibits Salmonella invasion through the post-translational control of HilD. Mol. Microbiol. 2013, 87, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Velge, P.; Bottreau, E.; Fievez, V.; Haesebrouck, F.; Ducatelle, R. Invasion of Salmonella enteritidis in avian intestinal epithelial cells in vitro is influenced by short-chain fatty acids. Int. J. Food Microbiol. 2003, 85, 237–248. [Google Scholar] [CrossRef]
- Lawhon, S.D.; Maurer, R.; Suyemoto, M.; Altier, C. Intestinal short-chain fatty acids alter Salmonella typhimurium invasion gene expression and virulence through BarA/SirA. Mol. Microbiol. 2002, 46, 1451–1464. [Google Scholar] [CrossRef]
- Palmieri, G.; Balestrieri, M.; Capuano, F.; Proroga, Y.T.R.; Pomilio, F.; Centorame, P.; Riccio, A.; Marrone, R.; Anastasio, A. Bactericidal and antibiofilm activity of bactenecin-derivative peptides against the food-pathogen Listeria monocytogenes: New perspectives for food processing industry. Int. J. Food Microbiol. 2018, 279, 33–42. [Google Scholar] [CrossRef]
- Agrillo, B.; Balestrieri, M.; Gogliettino, M.; Palmieri, G.; Moretta, R.; Proroga, Y.T.R.; Rea, I.; Cornacchia, A.; Capuano, F.; Smaldone, G.; et al. Functionalized polymeric materials with bio-derived antimicrobial peptides for “active” packaging. Int. J. Mol. Sci. 2019, 20, 601. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA, and ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar]
- Peng, M.; Biswas, D. Short chain and polyunsaturated fatty acids in host gut health and foodborne bacterial pathogen inhibition. Crit. Rev. Food Sci. Nutr. 2017, 57, 3987–4002. [Google Scholar] [CrossRef] [PubMed]
- Humayoun, S.B.; Hiott, L.M.; Gupta, S.K.; Barrett, J.B.; Woodley, T.A.; Johnston, J.J.; Jackson, C.R.; Frye, J.G. An assay for determining the susceptibility of Salmonella isolates to commercial and household biocides. PLoS ONE 2018, 13, e0209072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halstead, F.D.; Rauf, M.; Moiemen, N.S.; Bamford, A.; Wearn, C.M.; Fraise, A.P.; Lund, P.A.; Oppenheim, B.A.; Webber, M.A. The antibacterial activity of acetic acid against biofilm-producing pathogens of relevance to burns patients. PLoS ONE 2015, 10, e0136190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Effect of organic acids on biofilm formation and quorum signaling of pathogens from fresh fruits and vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Beier, R.C.; Harvey, R.B.; Hernandez, C.A.; Hume, M.E.; Andrews, K.; Droleskey, R.E.; Davidson, M.K.; Bodeis-Jones, S.; Young, S.; Duke, S.E.; et al. Interactions of organic acids with campylobacter coli from swine. PLoS ONE 2018, 13, e0202100. [Google Scholar] [CrossRef]
- Mine, S.; Boopathy, R. Effect of organic acids on shrimp pathogen, Vibrio harveyi. Curr. Microbiol. 2011, 63, 1–7. [Google Scholar] [CrossRef]
- Burt, S.A.; Adolfse, S.J.M.; Ahad, D.S.A.; Tersteeg-Zijderveld, M.H.G.; Jongerius-Gortemaker, B.G.M.; Post, J.A.; Brüggemann, H.; Santos, R.R. Cinnamaldehyde, Carvacrol and Organic Acids Affect Gene Expression of Selected Oxidative Stress and Inflammation Markers in IPEC-J2 Cells Exposed to Salmonella typhimurium. Phytother. Res. 2016, 30, 1988–2000. [Google Scholar] [CrossRef]
- Xu, J.; Koyanagi, Y.; Isogai, E.; Nakamura, S. Effects of fermentation products of the commensal bacterium Clostridium ramosum on motility, intracellular pH, and flagellar synthesis of enterohemorrhagic Escherichia coli. Arch. Microbiol. 2019, 201, 841–846. [Google Scholar] [CrossRef]
- Nakamura, S.; Kami-ike, N.; Yokota, J.-P.; Kudo, S.; Minamino, T.; Namba, K. Effect of Intracellular pH on the Torque-Speed Relationship of Bacterial Proton-Driven Flagellar Motor. J. Mol. Biol. 2009, 386, 332–338. [Google Scholar] [CrossRef]
- Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Salmonella and Campylobacter biofilm formation: A comparative assessment from farm to fork. J. Sci. Food Agric. 2018, 98, 4014–4032. [Google Scholar] [CrossRef]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Blana, V.; Georgomanou, A.; Giaouris, E. Assessing biofilm formation by Salmonella enterica serovar Typhimurium on abiotic substrata in the presence of quorum sensing signals produced by Hafnia alvei. Food Control 2017, 80, 83–91. [Google Scholar] [CrossRef]
- Gkana, E.; Giaouris, E.; Doulgeraki, A.; Kathariou, S.; Nychas, G.E. Biofilm formation by Salmonella Typhimurium and Staphylococcus aureus on stainless steel under either mono- or dual-species multi-strain conditions and resistance of sessile communities to sub-lethal chemical disinfection. Food Control 2017, 73, 838–846. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K. The talking language in some major Gram-negative bacteria. Arch. Microbiol. 2016, 198, 489–499. [Google Scholar] [CrossRef]
- Lianou, A.; Koutsoumanis, K.P. Strain variability of the behavior of foodborne bacterial pathogens: A review. Int. J. Food Microbiol. 2013, 167, 310–321. [Google Scholar] [CrossRef]
- Fàbrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [Green Version]
- International Standarization Organization. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection of Salmonella spp. (ISO 6579:2002); International Organization for Standardization: Geneva, Switzerland, 2003. [Google Scholar]
- Stepanovic, S.; Cirkovic, I.; Ranin, L.; Švabic-Vlahovic, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, B.Y.; Rayamahji, N.; Lee, H.S.; Jeon, W.J.; Choi, K.S.; Kweon, C.H.; Yoo, H.S. A multiplex real-time PCR for differential detection and quantification of Salmonella spp., Salmonella enterica serovar Typhimurium and Enteritidis in meats. J. Vet. Sci. 2009, 10, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Karavolos, M.H.; Bulmer, D.M.; Winzer, K.; Wilson, M.; Mastroeni, P.; Williams, P.; Khan, C.M.A. LuxS affects flagellar phase variation independently of quorum sensing in Salmonella enterica serovar typhimurium. J. Bacteriol. 2008, 190, 769–771. [Google Scholar] [CrossRef] [Green Version]
- Salaheen, S.; Jaiswal, E.; Joo, J.; Peng, M.; Ho, R.; OConnor, D.; Adlerz, K.; Aranda Espinoza, J.; Biswas, D. Bioactive extracts from berry byproducts on the pathogenicity of Salmonella Typhimurium. Int. J. Food Microbiol. 2016, 237, 128–135. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, X. Thermal resistance and gene expression of both desiccation-adapted and rehydrated Salmonella enterica serovar Typhimurium cells in aged broiler litter. Appl. Environ. Microbiol. 2017, 83, e00367-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Dong, Y.; Wang, G.; Xu, X.; Zhou, G. Effect of growth media on gene expression levels in Salmonella Typhimurium biofilm formed on stainless steel surface. Food Control 2016, 59, 546–552. [Google Scholar] [CrossRef]
- Aurass, P.; Düvel, J.; Karste, S.; Nübel, U.; Rabsch, W.; Flieger, A. glnA truncation in Salmonella enterica results in a small colony variant phenotype, attenuated host cell entry, and reduced expression of flagellin and SPI-1- associated effector genes. Appl. Environ. Microbiol. 2018, 84, e01838-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Jiang, Y.; Liu, X.; Qian, W.; Xu, X.; Zhou, G. Behavior variability of Salmonella enterica isolates from meat-related sources. LWT Food Sci. Technol. 2016, 73, 375–382. [Google Scholar] [CrossRef]
- Liu, J.; Zhai, L.; Lu, W.; Lu, Z.; Bie, X. Amino acid decarboxylase-dependent acid tolerance, selected phenotypic, and virulence gene expression responses of Salmonella enterica serovar Heidelberg. Food Res. Int. 2017, 92, 33–39. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Motility (mm) | ||||||
|---|---|---|---|---|---|---|
| Strain | Code | Control (n = 3) Mean ± SD | Acetic Acid (n = 3) Mean ± SD | Butyric Acid (n = 3) Mean ± SD | Propionic Acid (n = 3) Mean ± SD | Valeric Acid (n = 3) Mean ± SD |
| S. enterica subsp. arizonae | AZ3 | 16.00 ± 1.73 a | 2.33 ± 1.53 c | 6.00 ± 1.73 b,c | 5.00 ± 1.00 b,c | 8.67 ± 1.15 b |
| S. enterica subsp. arizonae | AZ26 | 17.00 ± 2.00 a | 2.00 ± 1.00 c | 5.33 ± 1.53 b,c | 5.67 ± 2.10 b,c | 8.00 ± 2.00 b |
| S. Bredeney | BR1 | 15.33 ± 1.15 a | 2.00 ± 1.73 c | 4.67 ± 1.15 c | 3.00 ± 1.00 c | 8.33 ± 1.53 b |
| S. Infantis | I18 | 15.33 ± 2.31 a | 1.67 ± 1.15 c | 3.67 ± 1.15 c | 3.67 ± 0.57 c | 8.67 ± 1.15 b |
| S. Infantis | I4 | 15.33 ± 1.53 a | 3.00 ± 1.73 c | 5.67 ± 1.15 b,c | 2.67 ± 1.53 c | 7.33 ± 2.08 b |
| S. Isangi | IG1 | 15.67 ± 2.08 a | 1.67 ± 0.58 c | 4.00 ± 1.00 c | 4.67 ± 1.15 c | 9.00 ± 1.73 b |
| S. Montevideo | M1 | 20.00 ± 2.00 a | 1.33 ± 0.58 c | 6.33 ± 2.08 b | 5.33 ± 0.58 b | 8.33 ± 0.58 b |
| S. Newport | N6 | 18.33 ± 1.53 a | 2.67 ± 1.53 d | 7.00 ± 1.73 b,c | 4.00 ± 2.00 c,d | 10.00 ± 2.00 b |
| S. Typhimurium | T1 | 20.00 ± 1.00 a | 3.67 ± 1.15 c | 4.67 ± 1.53 c | 6.00 ± 1.73 c | 10.00 ± 1.00 b |
| S. Typhimurium | T3 | 17.77 ± 2.52 a | 2.67 ± 0.58 c | 4.33 ± 1.53 b,c | 4.00 ± 2.65 b,c | 8.00 ± 1.00 b |
| S. Typhimurium | T7 | 19.33 ± 2.08 a | 3.33 ± 1.15 c | 5.33 ± 1.53 c | 3.33 ± 1.15 c | 10.67 ± 1.53 b |
| S. Typhimurium | T23 | 19.67 ± 2.10 a | 2.33 ± 2.31 c | 6.00 ± 1.00 b,c | 3.67 ± 2.10 c | 9.33 ± 2.52 b |
| Total | - | 17.36 ± 2.41 a | 2.43 ± 1.36 d | 4.12 ± 1.60 c | 4.24 ± 1.69 c | 8.45 ± 1.92 b |
| Strain | Code | Source |
|---|---|---|
| S. enterica subsp. arizonae | AZ3 | Poultry farm |
| S. enterica subsp. arizonae | AZ26 | Poultry farm |
| S. Bredeney | BR1 | Poultry farm |
| S. Infantis | I18 | Poultry farm |
| S. Infantis | I4 | Poultry farm |
| S. Isangi | IG1 | Poultry farm |
| S. Montevideo | M1 | Poultry farm |
| S. Newport | N6 | Poultry farm |
| S. Typhimurium | T1 | Chicken meat |
| S. Typhimurium | T3 | Chicken meat |
| S. Typhimurium | T7 | Poultry farm |
| S. Typhimurium | T23 | Poultry farm |
| Target Genes | Sequence (5′–3′) | Reference |
|---|---|---|
| 16s rRNA | F: AGGCCTTCGGGTTGTAAAGT R: GTTAGCCGGTGCTTCTTCTG | [39] |
| luxS | F: ATGCCATTATTAGATAGCTT R: GAGATGGTCGCGCATAAAGCCAGC | [40] |
| hilA | F: AATGGTCACAGGCTGAGGTG R: ACATCGTCGCGACTTGTGAA | [41] |
| invA | F: CGCGCTTGATGAGCTTTACC R: CTCGTAATTCGCCGCCATTG | [41] |
| rpoS | F: CAAGGGGAAATCCGTAAACCC R: GCCAATGGTGCCGAGTATC | [42] |
| csrA | F: CTGGACTGCTGGGATTTTTC R: CATGATTGGCGATGAGGTC | [43] |
| fliC | F: CTCGGCTACTGGTCTTGGTG R: CCGTAACGGTAACTTTGGCG | [44] |
| ssrA | F: CGGCTGGTATTCTTGTAAGGGT R: AAGCAGACACAAATTCGCAAG | [45] |
| stn | F: CAACCAGATAGTAAAGACCG R: ATTAGCGTAGAGGCAAAAGA | [46] |
| fadA | F: ATCTCTCCGCCCACTTAATGCGTA R: AGCCTTGCTCCAGCGTTTGTTGTA | [42] |
| sirA | F: CCAGCTACTTTCGCAGCAA R: AACACGTTGTAACGCGGTTG | [41] |
| spiA | F: AGGCGCTTGATATGTGC R: GCAGGCTCCGGAATTTTAGG | [46] |
| cpx | F: CATTTAACGACCGCGAGCTG R: ACCCGGATTAAGGCTTAGCG | [44] |
| rpoE | F: CACCTTACGGGAGCTGGATG R: GAAGATACGTGAACGCACCG | [44] |
| flijB | F: ATGGTACTACACTGGATGTATCG R: GTAAAGCCACCAATAGTAAC | [44] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamas, A.; Regal, P.; Vázquez, B.; Cepeda, A.; Franco, C.M. Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella enterica Motility, Biofilm Formation, and Gene Expression. Antibiotics 2019, 8, 265. https://doi.org/10.3390/antibiotics8040265
Lamas A, Regal P, Vázquez B, Cepeda A, Franco CM. Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella enterica Motility, Biofilm Formation, and Gene Expression. Antibiotics. 2019; 8(4):265. https://doi.org/10.3390/antibiotics8040265
Chicago/Turabian StyleLamas, Alexandre, Patricia Regal, Beatriz Vázquez, Alberto Cepeda, and Carlos Manuel Franco. 2019. "Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella enterica Motility, Biofilm Formation, and Gene Expression" Antibiotics 8, no. 4: 265. https://doi.org/10.3390/antibiotics8040265
APA StyleLamas, A., Regal, P., Vázquez, B., Cepeda, A., & Franco, C. M. (2019). Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella enterica Motility, Biofilm Formation, and Gene Expression. Antibiotics, 8(4), 265. https://doi.org/10.3390/antibiotics8040265