More Than a Pore: A Current Perspective on the In Vivo Mode of Action of the Lipopeptide Antibiotic Daptomycin


Abstract
:1. Introduction
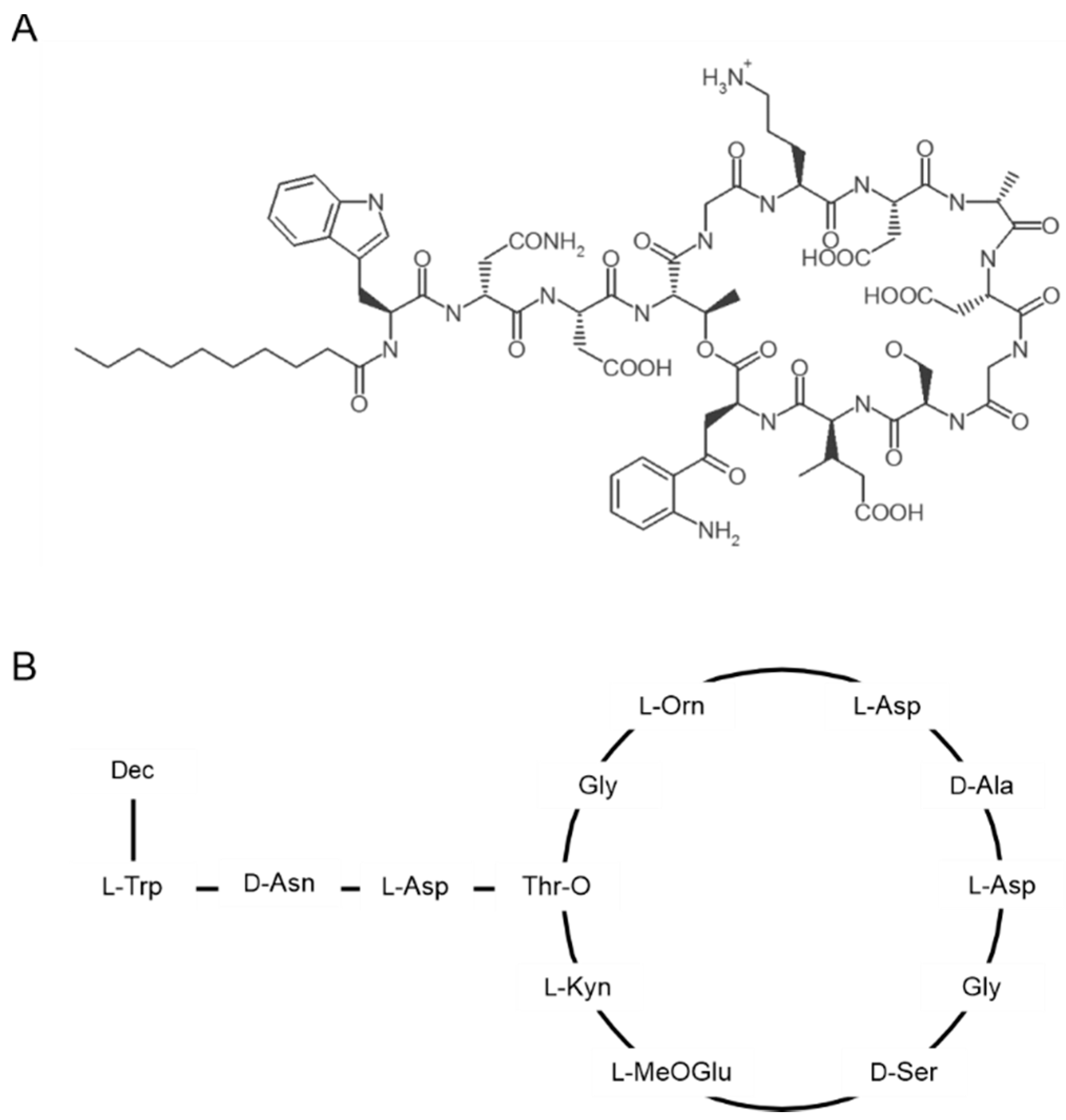
2. Structure and Oligomerization of Daptomycin
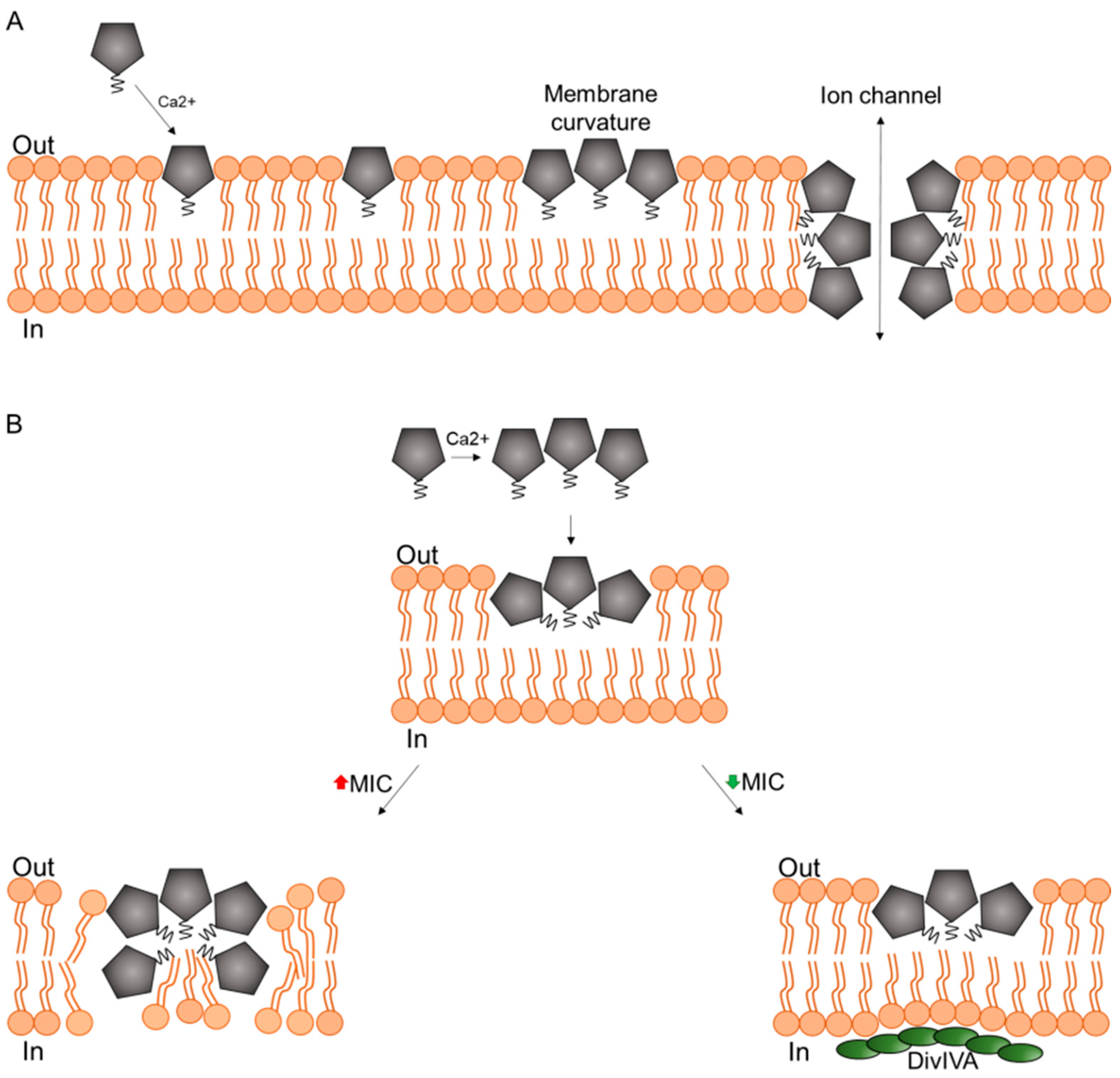
3. Mechanism of Action in Model Membranes

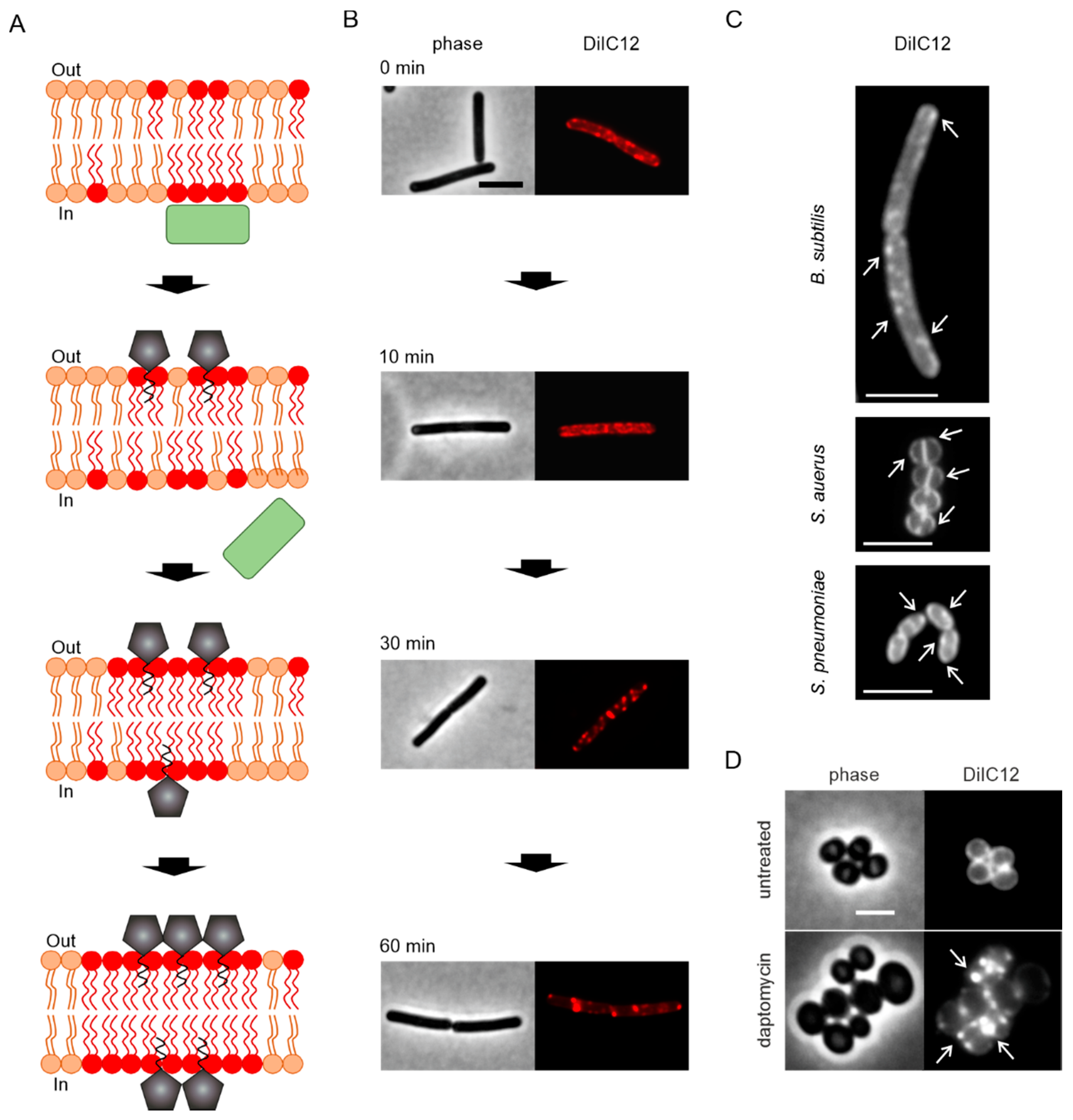
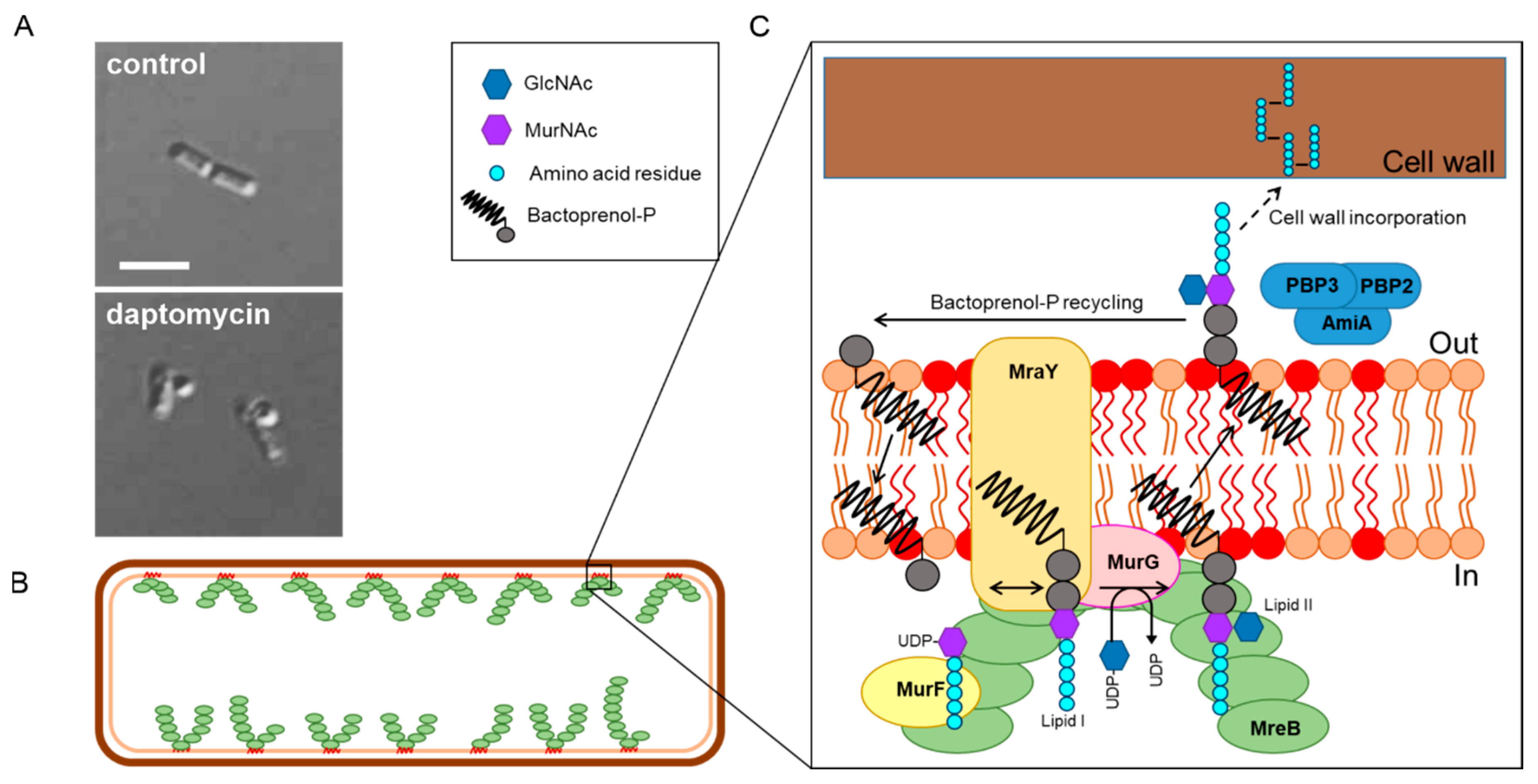
4. Pore Formation In Vivo
5. Is It Cell Wall Synthesis after All?
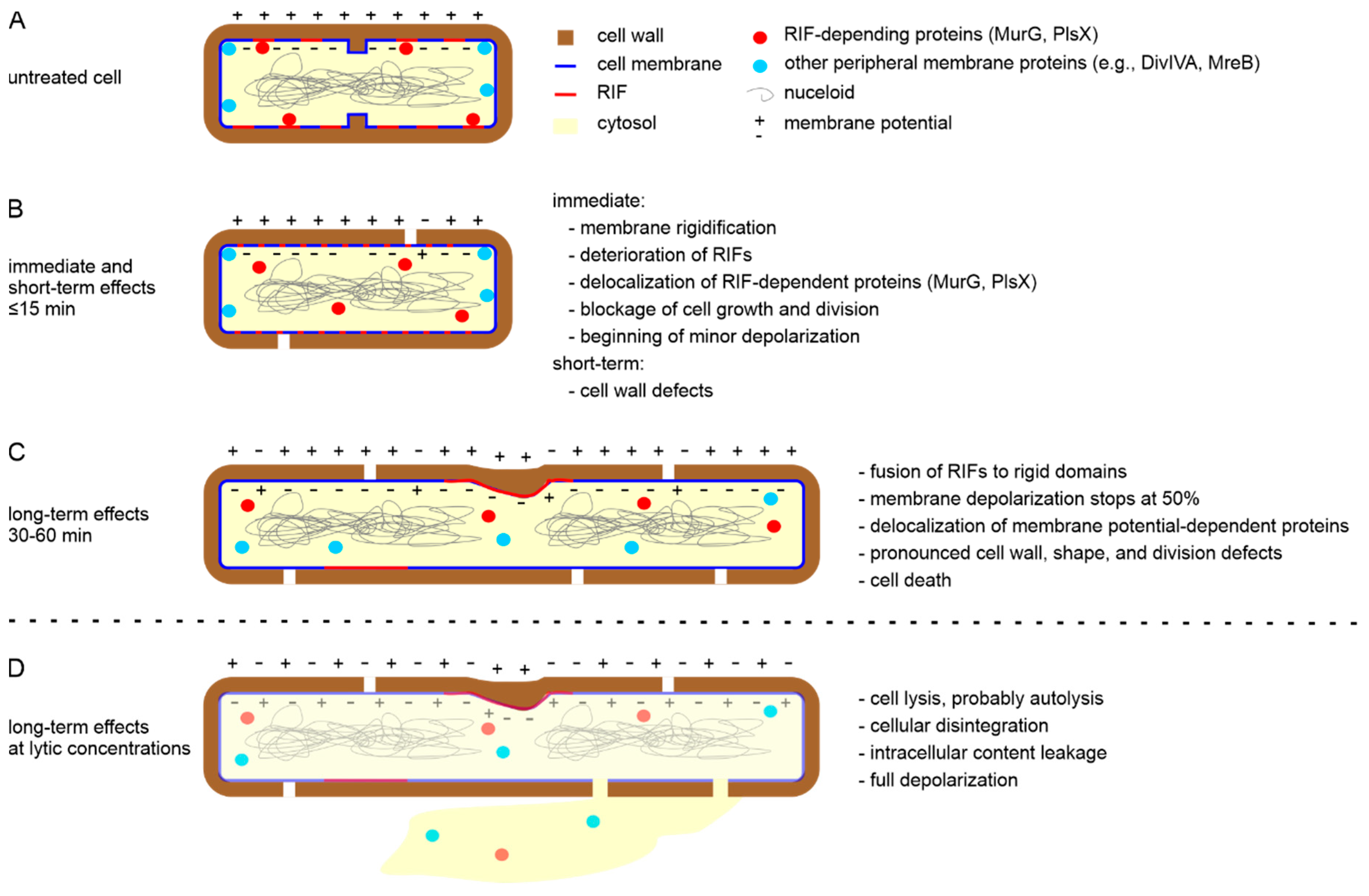
6. A New In Vivo Mode of Action Model
7. More to Discover

8. Lessons from Daptomycin Resistance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Mutated Gene | Gene Function | References |
|---|---|---|---|
| B. subtilis | pgsA | PG synthase | [25,43] |
| mprF | lysinylation of PG to lysyl-PG | [43] | |
| liaSR | cell envelope stress response | [25,43,92,137] | |
| S. aureus | pgsA | PG synthase | [44] |
| mprF | lysinylation of PG to lysyl-PG | [26,42] | |
| cls | cardiolipin synthase | [44,123,124,125,126] | |
| walKR | cell wall and membrane homeostasis | [37,138,139,140] | |
| dtlABCD | D-analylation of cell wall teichoic acids | [141,142,143,144,145,146] | |
| graRS | upregulation of dtl operon | [116,147] | |
| vraSR | cell envelope stress response | [52,148] | |
| SAOUHSC_00362 | hypothetical lipoprotein | [149] | |
| SAOUHSC_02441 | alkaline shock protein | [149] | |
| E. faecalis | cls | cardiolipin synthase | [127,128,129] |
| gdpD | glycerophosphoryl diester phosphodiesterase | [127,128] | |
| liaSR | cell envelope stress response | [127,128,150] | |
| E. faecium | cls | cardiolipin synthase | [127,130,131,132,133] |
| walKR | cell wall and membrane homeostasis | [133] | |
| liaSR | cell envelope stress response | [127,130,151,152,153] |
9. Closing the Gap between In Vitro and In Vivo
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Eliopoulos, G.M.; Willey, S.; Reiszner, E.; Spitzer, P.G.; Caputo, G.; Moellering, R.C.J. In vitro and in vivo activity of LY 146032, a new cyclic lipopeptide antibiotic. Antimicrob. Agents Chemother. 1986, 30, 532–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debono, M.; Barnhart, M.; Carrell, C.B.; Hoffmann, J.A.; Occolowitz, J.L.; Abbott, B.J.; Fukuda, D.S.; Hamill, R.L.; Biemann, K.; Herlihy, W.C. A21978C, a complex of new acidic peptide antibiotics: Isolation, chemistry, and mass spectral structure elucidation. J. Antibiot. 1987, 40, 761–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echevarria, K.; Datta, P.; Cadena, J.; Lewis, J.S. 2nd Severe myopathy and possible hepatotoxicity related to daptomycin. J. Antimicrob. Chemother. 2005, 55, 599–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauermann, R.; Rothenburger, M.; Graninger, W.; Joukhadar, C. Daptomycin: A review 4 years after first approval. Pharmacology 2008, 81, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H.; Miao, V.; Wrigley, S.K. Natural products to drugs: Daptomycin and related lipopeptide antibiotics. Nat. Prod. Rep. 2005, 22, 717–741. [Google Scholar] [CrossRef] [PubMed]
- Pirri, G.; Giuliani, A.; Nicoletto, S.F.; Pizzuto, L.; Rinaldi, A.C. Lipopeptides as anti-infectives: A practical perspective. Cent. Eur. J. Biol. 2009, 4, 258–273. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting bacterial membrane function: An underexploited mechanism for treating persistent infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, C.; Haldar, J. Membrane-Active Small Molecules: Designs Inspired by Antimicrobial Peptides. ChemMedChem 2015, 10, 1606–1624. [Google Scholar] [CrossRef]
- Allen, N.E.; Hobbs, J.N.; Alborn, W.E.J. Inhibition of peptidoglycan biosynthesis in gram-positive bacteria by LY146032. Antimicrob. Agents Chemother. 1987, 31, 1093–1099. [Google Scholar] [CrossRef] [Green Version]
- Huber, F.M.; Pieper, R.L.; Tietz, A.J. The formation of daptomycin by supplying decanoic acid to Streptomyces roseosporus cultures producing the antibiotic complex A21978C. J. Biotechnol. 1988, 7, 283–292. [Google Scholar] [CrossRef]
- Grunewald, J.; Sieber, S.A.; Mahlert, C.; Linne, U.; Marahiel, M.A.; Grünewald, J.; Sieber, S.A.; Mahlert, C.; Linne, U.; Marahiel, M.A. Synthesis and Derivatization of Daptomycin: A Chemoenzymatic Route to Acidic Lipopeptide Antibiotics. J. Am. Chem. Soc. 2004, 126, 17025–17031. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.; Barnawi, G.; Beriashvili, D.; Palmer, M.; Taylor, S.D. The effect of replacing the ester bond with an amide bond and of overall stereochemistry on the activity of daptomycin. Bioorg. Med. Chem. 2019, 27, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Hojati, Z.; Milne, C.; Harvey, B.; Gordon, L.; Borg, M.; Flett, F.; Wilkinson, B.; Sidebottom, P.J.; Rudd, B.A.M.; Hayes, M.A.; et al. Structure, biosynthetic origin, and engineered biosynthesis of calcium-dependent antibiotics from Streptomyces coelicolor. Chem. Biol. 2002, 9, 1175–1187. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Sun, T.-L.; Sun, Y.; Huang, H.W. Interaction of daptomycin with lipid bilayers: A lipid extracting effect. Biochemistry 2014, 53, 5384–5392. [Google Scholar] [CrossRef] [Green Version]
- Scott, W.R.P.; Baek, S.-B.; Jung, D.; Hancock, R.E.W.; Straus, S.K. NMR structural studies of the antibiotic lipopeptide daptomycin in DHPC micelles. Biochim. Biophys. Acta 2007, 1768, 3116–3126. [Google Scholar] [CrossRef] [Green Version]
- Rotondi, K.S.; Gierasch, L.M. A well-defined amphipathic conformation for the calcium-free cyclic lipopeptide antibiotic, daptomycin, in aqueous solution. Biopolymers 2005, 80, 374–385. [Google Scholar] [CrossRef]
- Ball, L.-J.; Goult, C.M.; Donarski, J.A.; Micklefield, J.; Ramesh, V. NMR structure determination and calcium binding effects of lipopeptide antibiotic daptomycin. Org. Biomol. Chem. 2004, 2, 1872–1878. [Google Scholar] [CrossRef]
- Jung, D.; Rozek, A.; Okon, M.; Hancock, R.E.W. Structural transitions as determinants of the action of the calcium-dependent antibiotic daptomycin. Chem. Biol. 2004, 11, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-T.; Hung, W.-C.; Hsieh, M.-H.; Chen, H.; Chang, Y.-Y.; Huang, H.W. Molecular State of the Membrane-Active Antibiotic Daptomycin. Biophys. J. 2017, 113, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.W.; Jung, D.; Calhoun, J.R.; Lear, J.D.; Okon, M.; Scott, W.R.P.; Hancock, R.E.W.; Straus, S.K. Effect of divalent cations on the structure of the antibiotic daptomycin. Eur. Biophys. J. 2008, 37, 421–433. [Google Scholar] [CrossRef]
- Muraih, J.K.; Pearson, A.; Silverman, J.; Palmer, M. Oligomerization of daptomycin on membranes. Biochim. Biophys. Acta 2011, 1808, 1154–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraih, J.K.; Palmer, M. Estimation of the subunit stoichiometry of the membrane-associated daptomycin oligomer by FRET. Biochim. Biophys. Acta 2012, 1818, 1642–1647. [Google Scholar] [CrossRef] [Green Version]
- Muraih, J.K.; Harris, J.; Taylor, S.D.; Palmer, M. Characterization of daptomycin oligomerization with perylene excimer fluorescence: Stoichiometric binding of phosphatidylglycerol triggers oligomer formation. Biochim. Biophys. Acta 2012, 1818, 673–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreutzberger, M.A.; Pokorny, A.; Almeida, P.F. Daptomycin-Phosphatidylglycerol Domains in Lipid Membranes. Langmuir 2017, 33, 13669–13679. [Google Scholar] [CrossRef] [PubMed]
- Hachmann, A.B.; Sevim, E.; Gaballa, A.; Popham, D.L.; Antelmann, H.; Helmann, J.D. Reduction in membrane phosphatidylglycerol content leads to daptomycin resistance in Bacillus subtilis. Antimicrob. Agents Chemother. 2011, 55, 4326–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, A.S.; Schneider, T.; Sahl, H.-G. Mechanisms of daptomycin resistance in Staphylococcus aureus: Role of the cell membrane and cell wall. Ann. N. Y. Acad. Sci. 2013, 1277, 139–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-J.; Mishra, N.N.; Rubio, A.; Bayer, A.S. Causal role of single nucleotide polymorphisms within the mprF gene of Staphylococcus aureus in daptomycin resistance. Antimicrob. Agents Chemother. 2013, 57, 5658–5664. [Google Scholar] [CrossRef] [Green Version]
- Rubio, A.; Moore, J.; Varoglu, M.; Conrad, M.; Chu, M.; Shaw, W.; Silverman, J.A. LC-MS/MS characterization of phospholipid content in daptomycin-susceptible and -resistant isolates of Staphylococcus aureus with mutations in mprF. Mol. Membr. Biol. 2012, 29, 1–8. [Google Scholar] [CrossRef]
- Andra, J.; Goldmann, T.; Ernst, C.M.; Peschel, A.; Gutsmann, T. Multiple peptide resistance factor (MprF)-mediated Resistance of Staphylococcus aureus against antimicrobial peptides coincides with a modulated peptide interaction with artificial membranes comprising lysyl-phosphatidylglycerol. J. Biol. Chem. 2011, 286, 18692–18700. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C.M.; Peschel, A. Broad-spectrum antimicrobial peptide resistance by MprF-mediated aminoacylation and flipping of phospholipids. Mol. Microbiol. 2011, 80, 290–299. [Google Scholar] [CrossRef]
- Nishi, H.; Komatsuzawa, H.; Fujiwara, T.; McCallum, N.; Sugai, M. Reduced content of lysyl-phosphatidylglycerol in the cytoplasmic membrane affects susceptibility to moenomycin, as well as vancomycin, gentamicin, and antimicrobial peptides, in Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 4800–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staubitz, P.; Neumann, H.; Schneider, T.; Wiedemann, I.; Peschel, A. MprF-mediated biosynthesis of lysylphosphatidylglycerol, an important determinant in staphylococcal defensin resistance. FEMS Microbiol. Lett. 2004, 231, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.N.; Yang, S.-J.; Chen, L.; Muller, C.; Saleh-Mghir, A.; Kuhn, S.; Peschel, A.; Yeaman, M.R.; Nast, C.C.; Kreiswirth, B.N.; et al. Emergence of daptomycin resistance in daptomycin-naive rabbits with methicillin-resistant Staphylococcus aureus prosthetic joint infection is associated with resistance to host defense cationic peptides and mprF polymorphisms. PLoS ONE 2013, 8, e71151. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-J.; Xiong, Y.Q.; Dunman, P.M.; Schrenzel, J.; Francois, P.; Peschel, A.; Bayer, A.S. Regulation of mprF in daptomycin-nonsusceptible Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2009, 53, 2636–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slavetinsky, C.J.; Peschel, A.; Ernst, C.M. Alanyl-phosphatidylglycerol and lysyl-phosphatidylglycerol are translocated by the same MprF flippases and have similar capacities to protect against the antibiotic daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 3492–3497. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C.M.; Kuhn, S.; Slavetinsky, C.J.; Krismer, B.; Heilbronner, S.; Gekeler, C.; Kraus, D.; Wagner, S.; Peschel, A. The lipid-modifying multiple peptide resistance factor is an oligomer consisting of distinct interacting synthase and flippase subunits. MBio 2015, 6, e02340-14. [Google Scholar] [CrossRef] [Green Version]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 2137–2145. [Google Scholar] [CrossRef] [Green Version]
- Bayer, A.S.; Mishra, N.N.; Sakoulas, G.; Nonejuie, P.; Nast, C.C.; Pogliano, J.; Chen, K.-T.; Ellison, S.N.; Yeaman, M.R.; Yang, S.-J. Heterogeneity of mprF sequences in methicillin-resistant Staphylococcus aureus clinical isolates: Role in cross-resistance between daptomycin and host defense antimicrobial peptides. Antimicrob. Agents Chemother. 2014, 58, 7462–7467. [Google Scholar] [CrossRef] [Green Version]
- Bayer, A.S.; Mishra, N.N.; Chen, L.; Kreiswirth, B.N.; Rubio, A.; Yang, S.-J. Frequency and Distribution of Single-Nucleotide Polymorphisms within mprF in Methicillin-Resistant Staphylococcus aureus Clinical Isolates and Their Role in Cross-Resistance to Daptomycin and Host Defense Antimicrobial Peptides. Antimicrob. Agents Chemother. 2015, 59, 4930–4937. [Google Scholar] [CrossRef] [Green Version]
- Cameron, D.R.; Mortin, L.I.; Rubio, A.; Mylonakis, E.; Moellering, R.C.J.; Eliopoulos, G.M.; Peleg, A.Y. Impact of daptomycin resistance on Staphylococcus aureus virulence. Virulence 2015, 6, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C.M.; Staubitz, P.; Mishra, N.N.; Yang, S.-J.; Hornig, G.; Kalbacher, H.; Bayer, A.S.; Kraus, D.; Peschel, A. The bacterial defensin resistance protein MprF consists of separable domains for lipid lysinylation and antimicrobial peptide repulsion. PLoS Pathog. 2009, 5, e1000660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, A.; Conrad, M.; Haselbeck, R.J.; Kedar, G.C.; Brown-Driver, V.; Finn, J.; Silverman, J.A. Regulation of mprF by antisense RNA restores daptomycin susceptibility to daptomycin-resistant isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 364–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hachmann, A.-B.; Angert, E.R.; Helmann, J.D. Genetic Analysis of Factors Affecting Susceptibility of Bacillus subtilis to Daptomycin. Antimicrob. Agents Chemother. 2009, 53, 1598–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, A.Y.; Miyakis, S.; Ward, D.V.; Earl, A.M.; Rubio, A.; Cameron, D.R.; Pillai, S.; Moellering, R.C.J.; Eliopoulos, G.M. Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS ONE 2012, 7, e28316. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.P.; Mariner, K.R.; Chopra, I.; O’Neill, A.J. The target of daptomycin is absent from Escherichia coli and other gram-negative pathogens. Antimicrob. Agents Chemother. 2013, 57, 637–639. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.F.; Savage, P.B.; Epand, R.M. Bacterial lipid composition and the antimicrobial efficacy of cationic steroid compounds (Ceragenins). Biochim. Biophys. Acta 2007, 1768, 2500–2509. [Google Scholar] [CrossRef] [Green Version]
- Straus, S.K.; Hancock, R.E.W. Mode of action of the new antibiotic for Gram-positive pathogens daptomycin: Comparison with cationic antimicrobial peptides and lipopeptides. Biochim. Biophys. Acta 2006, 1758, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Kinouchi, H.; Onishi, M.; Kamimori, H. Lipid membrane-binding properties of daptomycin using surface plasmon resonance. Anal. Sci. 2013, 29, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Beriashvili, D.; Taylor, R.; Kralt, B.; Abu Mazen, N.; Taylor, S.D.; Palmer, M. Mechanistic studies on the effect of membrane lipid acyl chain composition on daptomycin pore formation. Chem. Phys. Lipids 2018, 216, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Muraih, J.K.; Tishbi, N.; Herskowitz, J.; Victor, R.L.; Silverman, J.; Uwumarenogie, S.; Taylor, S.D.; Palmer, M.; Mintzer, E. Cardiolipin prevents membrane translocation and permeabilization by daptomycin. J. Biol. Chem. 2014, 289, 11584–11591. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.D.; Palmer, M. The action mechanism of daptomycin. Bioorg. Med. Chem. 2016, 24, 6253–6268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.R.; Bayer, A.S.; Arias, C.A. Mechanism of Action and Resistance to Daptomycin in Staphylococcus aureus and Enterococci. Cold Spring Harb. Perspect. Med. 2016, 6, a026997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Muraih, J.K.; MacCormick, B.; Silverman, J.; Palmer, M. Daptomycin forms cation- and size-selective pores in model membranes. Biochim. Biophys. Acta 2014, 1838, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, A.; Wenzel, M.; Strahl, H.; Grein, F.; Saaki, T.N.V.; Kohl, B.; Siersma, T.; Bandow, J.E.; Sahl, H.-G.; Schneider, T.; et al. Daptomycin inhibits bacterial cell envelope synthesis by interfering with fluid membrane microdomains. Proc. Natl. Acad. Sci. USA 2016, 113, E7077–E7086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Scoten, K.; Straus, S.K. Daptomycin Leakage Is Selective. ACS Infect. Dis. 2016, 2, 682–687. [Google Scholar] [CrossRef]
- Silverman, J.A.; Perlmutter, N.G.; Shapiro, H.M. Correlation of daptomycin bactericidal activity and membrane depolarization in Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 2538–2544. [Google Scholar] [CrossRef] [Green Version]
- Boudjemaa, R.; Cabriel, C.; Dubois-Brissonnet, F.; Bourg, N.; Dupuis, G.; Gruss, A.; Lévêque-Fort, S.; Briandet, R.; Fontaine-Aupart, M.-P.; Steenkeste, K. Impact of bacterial membrane fatty acid composition on the failure of daptomycin to kill Staphylococcus aureus. Antimicrob. Agents Chemother. 2018, 62, e00023-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Wang, W.; Dai, S.; Liu, T.; Tan, J.; Qu, G.; Li, Y.; Ling, Y.; Liu, G.; Fu, X.; et al. Daptomycin exerts rapid bactericidal activity against Bacillus anthracis without disrupting membrane integrity. Acta Pharmacol. Sin. 2014, 35, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-T.; Yang, P.-Y.; Charron, N.E.; Hsieh, M.-H.; Chang, Y.-Y.; Huang, H.W. Comparison of the Effects of Daptomycin on Bacterial and Model Membranes. Biochemistry 2018, 57, 5629–5639. [Google Scholar] [CrossRef]
- Taylor, R.; Butt, K.; Scott, B.; Zhang, T.; Muraih, J.K.; Mintzer, E.; Taylor, S.; Palmer, M. Two successive calcium-dependent transitions mediate membrane binding and oligomerization of daptomycin and the related antibiotic A54145. Biochim. Biophys. Acta 2016, 1858, 1999–2005. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Scott, W.R.P.; Gabel, F.; Wu, M.; Desmond, R.; Bae, J.; Zaccai, G.; Algar, W.R.; Straus, S.K. On the quest for the elusive mechanism of action of daptomycin: Binding, fusion, and oligomerization. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Beriashvili, D.; Taylor, S.; Palmer, M. Daptomycin Pore Formation Is Restricted by Lipid Acyl Chain Composition. ACS Infect. Dis. 2017, 3, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.M.; Pollett, S.; Sakoulas, G. A current perspective on daptomycin for the clinical microbiologist. Clin. Microbiol. Rev. 2013, 26, 759–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogliano, J.; Pogliano, N.; Silverman, J.A. Daptomycin-mediated reorganization of membrane architecture causes mislocalization of essential cell division proteins. J. Bacteriol. 2012, 194, 4494–4504. [Google Scholar] [CrossRef]
- Munch, D.; Muller, A.; Schneider, T.; Kohl, B.; Wenzel, M.; Bandow, J.E.; Maffioli, S.; Sosio, M.; Donadio, S.; Wimmer, R.; et al. The lantibiotic NAI-107 binds to bactoprenol-bound cell wall precursors and impairs membrane functions. J. Biol. Chem. 2014, 289, 12063–12076. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Kohl, B.; Münch, D.; Raatschen, N.; Albada, H.B.; Hamoen, L.; Metzler-Nolte, N.; Sahl, H.G.; Bandow, J.E. Proteomic response of Bacillus subtilis to lantibiotics reflects differences in interaction with the cytoplasmic membrane. Antimicrob. Agents Chemother. 2012, 56, 5749–5757. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Rautenbach, M.; Vosloo, J.; Siersma, T.; Aisenbrey, C.; Zaitseva, E.; Laubscher, W.; van Rensburg, W.; Behrends, J.; Bechinger, B.; et al. The multifaceted antibacterial mechanisms of the pioneering peptide antibiotics tyrocidine and gramicidin S. MBio 2018, 9, e00802-18. [Google Scholar] [CrossRef] [Green Version]
- Rangarajan, N.; Bakshi, S.; Weisshaar, J.C. Localized permeabilization of E. coli membranes by the antimicrobial peptide Cecropin, A. Biochemistry 2013, 52, 6584–6594. [Google Scholar] [CrossRef] [Green Version]
- Barns, K.J.; Weisshaar, J.C. Real-time attack of LL-37 on single Bacillus subtilis cells. Biochim. Biophys. Acta 2013, 1828, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Barns, K.J.; Weisshaar, J.C. Single-cell, time-resolved study of the effects of the antimicrobial peptide alamethicin on Bacillus subtilis. Biochim. Biophys. Acta 2016, 1858, 725–732. [Google Scholar] [CrossRef]
- Cotroneo, N.; Harris, R.; Perlmutter, N.; Beveridge, T.; Silverman, J.A. Daptomycin exerts bactericidal activity without lysis of Staphylococcus aureus. Antimicrob. Agents Chemother. 2008, 52, 2223–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, J.K.; Miller, K.; O’Neill, A.J.; Chopra, I. Consequences of daptomycin-mediated membrane damage in Staphylococcus aureus. J. Antimicrob. Chemother. 2008, 62, 1003–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mensa, B.; Howell, G.L.; Scott, R.; DeGrado, W.F. Comparative mechanistic studies of brilacidin, daptomycin, and the antimicrobial peptide LL16. Antimicrob. Agents Chemother. 2014, 58, 5136–5145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hover, B.M.; Kim, S.-H.; Katz, M.; Charlop-Powers, Z.; Owen, J.G.; Ternei, M.A.; Maniko, J.; Estrela, A.B.; Molina, H.; Park, S.; et al. Culture-independent discovery of the malacidins as calcium-dependent antibiotics with activity against multidrug-resistant Gram-positive pathogens. Nat. Microbiol. 2018, 3, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Rubinchik, E.; Schneider, T.; Elliott, M.; Scott, W.R.P.; Pan, J.; Anklin, C.; Yang, H.; Dugourd, D.; Muller, A.; Gries, K.; et al. Mechanism of action and limited cross-resistance of new lipopeptide MX-2401. Antimicrob. Agents Chemother. 2011, 55, 2743–2754. [Google Scholar] [CrossRef] [Green Version]
- Cauz, A.C.G.; Carretero, G.P.B.; Saraiva, G.K.V.; Park, P.; Mortara, L.; Cuccovia, I.M.; Brocchi, M.; Gueiros-Filho, F.J. Violacein Targets the Cytoplasmic Membrane of Bacteria. ACS Infect. Dis. 2019, 5, 539–549. [Google Scholar] [CrossRef]
- Seydlová, G.; Sokol, A.; Lišková, P.; Konopásek, I.; Fišer, R. Daptomycin Pore Formation and Stoichiometry Depend on Membrane Potential of Target Membrane. Antimicrob. Agents Chemother. 2019, 63, e01589-18. [Google Scholar] [CrossRef] [Green Version]
- Canepari, P.; Boaretti, M.; Del Mar Lleo, M.; Satta, G.; Lleo, M.M.; Satta, G. Lipoteichoic acid as a new target for activity of antibiotics: Mode of action of daptomycin (LY146032). Antimicrob. Agents Chemother. 1990, 34, 1220–1226. [Google Scholar] [CrossRef] [Green Version]
- Boaretti, M.; Canepari, P. Identification of daptomycin-binding proteins in the membrane of Enterococcus hirae. Antimicrob. Agents Chemother. 1995, 39, 2068–2072. [Google Scholar] [CrossRef] [Green Version]
- Laganas, V.; Alder, J.; Silverman, J.A. In vitro bactericidal activities of daptomycin against Staphylococcus aureus and Enterococcus faecalis are not mediated by inhibition of lipoteichoic acid biosynthesis. Antimicrob. Agents Chemother. 2003, 47, 2682–2684. [Google Scholar] [CrossRef] [Green Version]
- Boaretti, M.; Canepari, P.; Lleo, M.M.; Satta, G. The activity of daptomycin on Enterococcus faecium protoplasts: Indirect evidence supporting a novel mode of action on lipoteichoic acid synthesis. J. Antimicrob. Chemother. 1993, 31, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Mengin-Lecreulx, D.; Allen, N.E.; Hobbs, J.N.; van Heijenoort, J. Inhibition of peptidoglycan biosynthesis in Bacillus megaterium by daptomycin. FEMS Microbiol. Lett. 1990, 57, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Wale, L.J.; Shelton, A.P.; Greenwood, D. Scanning electronmicroscopy of Staphylococcus aureus and Enterococcus faecalis exposed to daptomycin. J. Med. Microbiol. 1989, 30, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Hashizume, H.; Sawa, R.; Harada, S.; Igarashi, M.; Adachi, H.; Nishimura, Y.; Nomoto, A. Tripropeptin C blocks the lipid cycle of cell wall biosynthesis by complex formation with undecaprenyl pyrophosphate. Antimicrob. Agents Chemother. 2011, 55, 3821–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, T.; Gries, K.; Josten, M.; Wiedemann, I.; Pelzer, S.; Labischinski, H.; Sahl, H.G. The lipopeptide antibiotic friulimicin B inhibits cell wall biosynthesis through complex formation with bactoprenol phosphate. Antimicrob. Agents Chemother. 2009, 53, 1610–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantibhedhyangkul, W.; Wongsawat, E.; Matamnan, S.; Inthasin, N.; Sueasuay, J.; Suputtamongkol, Y. Anti-Mycoplasma Activity of Daptomycin and Its Use for Mycoplasma Elimination in Cell Cultures of Rickettsiae. Antibiotics 2019, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, D.; Dominguez-Cuevas, P.; Daniel, R.A.; Mascher, T. Cell envelope stress response in cell wall-deficient L-forms of Bacillus subtilis. Antimicrob. Agents Chemother. 2012, 56, 5907–5915. [Google Scholar] [CrossRef] [Green Version]
- Mascio, C.T.M.; Alder, J.D.; Silverman, J.A. Bactericidal action of daptomycin against stationary-phase and nondividing Staphylococcus aureus cells. Antimicrob. Agents Chemother. 2007, 51, 4255–4260. [Google Scholar] [CrossRef] [Green Version]
- Renzoni, A.; Kelley, W.L.; Rosato, R.R.; Martinez, M.P.; Roch, M.; Fatouraei, M.; Haeusser, D.P.; Margolin, W.; Fenn, S.; Turner, R.D.; et al. Molecular Bases Determining Daptomycin Resistance-Mediated Resensitization to beta-Lactams (Seesaw Effect) in Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2017, 61, e01634-16. [Google Scholar] [CrossRef] [Green Version]
- Rand, K.H.; Houck, H. Daptomycin synergy with rifampicin and ampicillin against vancomycin-resistant enterococci. J. Antimicrob. Chemother. 2004, 53, 530–532. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Xia, Z.; Zhang, D.; Sheng, Z.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Multifunctional Pharmaceutical Effects of the Antibiotic Daptomycin. Biomed. Res. Int. 2019, 2019, 8609218. [Google Scholar] [CrossRef] [PubMed]
- Wecke, T.; Zühlke, D.; Mäder, U.; Jordan, S.; Voigt, B.; Pelzer, S.; Labischinski, H.; Homuth, G.; Hecker, M.; Mascher, T. Daptomycin versus friulimicin B: In-depth profiling of the Bacillus subtilis cell envelope stress responses. Antimicrob. Agents Chemother. 2009, 53, 1619–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Zhang, D.; Li, G.; Liu, J.; He, G.; Zhang, P.; Yang, L.; Zhu, H.; Xu, N.; Liang, S. Antibacterial mechanism of daptomycin antibiotic against Staphylococcus aureus based on a quantitative bacterial proteome analysis. J. Proteomics 2017, 150, 242–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthaiyan, A.; Silverman, J.A.; Jayaswal, R.K.; Wilkinson, B.J. Transcriptional profiling reveals that daptomycin induces the Staphylococcus aureus cell wall stress stimulon and genes responsive to membrane depolarization. Antimicrob. Agents Chemother. 2008, 52, 980–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dengler, V.; Meier, P.S.; Heusser, R.; Berger-Bachi, B.; McCallum, N. Induction kinetics of the Staphylococcus aureus cell wall stress stimulon in response to different cell wall active antibiotics. BMC Microbiol. 2011, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Strahl, H.; Burmann, F.; Hamoen, L.W. The actin homologue MreB organizes the bacterial cell membrane. Nat. Commun. 2014, 5, 3442. [Google Scholar] [CrossRef] [Green Version]
- Oswald, F.; Varadarajan, A.; Lill, H.; Peterman, E.J.G.; Bollen, Y.J.M. MreB-Dependent Organization of the E. coli Cytoplasmic Membrane Controls Membrane Protein Diffusion. Biophys. J. 2016, 110, 1139–1149. [Google Scholar] [CrossRef] [Green Version]
- Saeloh, D.; Tipmanee, V.; Jim, K.K.; Dekker, M.P.; Bitter, W.; Voravuthikunchai, S.P.; Wenzel, M.; Hamoen, L.W.L.W. The novel antibiotic rhodomyrtone traps membrane proteins in vesicles with increased fluidity. PLoS Pathog. 2018, 14, e1006876. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Dekker, M.P.; Wang, B.; Burggraaf, M.J.; Bitter, W.; van Weering, J.R.T.; Hamoen, L.W. New flat embedding method for transmission electron microscopy reveals an unknown mechanism of tetracycline. BioRxiv 2019, 820191. [Google Scholar] [CrossRef] [Green Version]
- Strahl, H.; Hamoen, L.W. Membrane potential is important for bacterial cell division. Proc. Natl. Acad. Sci. USA 2010, 107, 12281–12286. [Google Scholar] [CrossRef] [Green Version]
- Falk, S.P.; Noah, J.W.; Weisblum, B. Screen for inducers of autolysis in Bacillus subtilis. Antimicrob. Agents Chemother. 2010, 54, 3723–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafiso, V.; Bertuccio, T.; Spina, D.; Purrello, S.; Campanile, F.; Di Pietro, C.; Purrello, M.; Stefani, S. Modulating activity of vancomycin and daptomycin on the expression of autolysis cell-wall turnover and membrane charge genes in hVISA and VISA strains. PLoS ONE 2012, 7, e29573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barak, I.; Muchova, K.; Wilkinson, A.J.; O’Toole, P.J.; Pavlendova, N. Lipid spirals in Bacillus subtilis and their role in cell division. Mol. Microbiol. 2008, 68, 1315–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchová, K.; Wilkinson, A.J.; Barák, I. Changes of lipid domains in Bacillus subtilis cells with disrupted cell wall peptidoglycan. FEMS Microbiol. Lett. 2011, 325, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Escobar, J.; Chastanet, A.; Crevenna, A.H.; Fromion, V.; Wedlich-Soldner, R.; Carballido-Lopez, R. Processive movement of MreB-associated cell wall biosynthetic complexes in bacteria. Science 2011, 333, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Garner, E.C.; Bernard, R.; Wang, W.; Zhuang, X.; Rudner, D.Z.; Mitchison, T. Coupled, circumferential motions of the cell wall synthesis machinery and MreB filaments in B. subtilis. Science 2011, 333, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Wivagg, C.N.; Szwedziak, P.; Wong, F.; Schaefer, K.; Izoré, T.; Renner, L.D.; Holmes, M.J.; Sun, Y.; Bisson-Filho, A.W.; et al. MreB filaments align along greatest principal membrane curvature to orient cell wall synthesis. Elife 2018, 7, e32471. [Google Scholar] [CrossRef]
- Schirner, K.; Eun, Y.-J.; Dion, M.; Luo, Y.; Helmann, J.D.; Garner, E.C.; Walker, S. Lipid-linked cell wall precursors regulate membrane association of bacterial actin MreB. Nat. Chem. Biol. 2015, 11, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Koch, D.C.; Schmidt, T.H.; Sahl, H.-G.; Kubitscheck, U.; Kandt, C. Structural dynamics of the cell wall precursor lipid II in the presence and absence of the lantibiotic nisin. Biochim. Biophys. Acta 2014, 1838, 3061–3068. [Google Scholar] [CrossRef] [Green Version]
- Scherer, K.M.; Spille, J.-H.; Sahl, H.-G.; Grein, F.; Kubitscheck, U. The lantibiotic nisin induces lipid II aggregation, causing membrane instability and vesicle budding. Biophys. J. 2015, 108, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Schneider, T.; Kruse, T.; Wimmer, R.; Wiedemann, I.; Sass, V.; Pag, U.; Jansen, A.; Nielsen, A.K.; Mygind, P.H.; Raventós, D.S.; et al. Plectasin, a fungal defensin, targets the bacterial cell wall precursor Lipid II. Science 2010, 328, 1168–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sass, V.; Schneider, T.; Wilmes, M.; Körner, C.; Tossi, A.; Novikova, N.; Shamova, O.; Sahl, H.G. Human $β$-defensin 3 inhibits cell wall biosynthesis in staphylococci. Infect. Immun. 2010, 78, 2793–2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, C.M.; Peschel, A. MprF-mediated daptomycin resistance. Int. J. Med. Microbiol. 2019, 309, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Van der Es, D.; Hogendorf, W.F.J.; Overkleeft, H.S.; van der Marel, G.A.; Codee, J.D.C. Teichoic acids: Synthesis and applications. Chem. Soc. Rev. 2017, 46, 1464–1482. [Google Scholar] [CrossRef] [Green Version]
- Formstone, A.; Carballido-Lopez, R.; Noirot, P.; Errington, J.; Scheffers, D.-J. Localization and interactions of teichoic acid synthetic enzymes in Bacillus subtilis. J. Bacteriol. 2008, 190, 1812–1821. [Google Scholar] [CrossRef] [Green Version]
- Muller, A.; Grein, F.; Otto, A.; Gries, K.; Orlov, D.; Zarubaev, V.; Girard, M.; Sher, X.; Shamova, O.; Roemer, T.; et al. Differential daptomycin resistance development in Staphylococcus aureus strains with active and mutated gra regulatory systems. Int. J. Med. Microbiol. 2018, 308, 335–348. [Google Scholar] [CrossRef]
- Wilmes, M.; Stockem, M.; Bierbaum, G.; Schlag, M.; Gotz, F.; Tran, D.Q.; Schaal, J.B.; Ouellette, A.J.; Selsted, M.E.; Sahl, H.-G. Killing of staphylococci by theta-defensins involves membrane impairment and activation of autolytic enzymes. Antibiotics 2014, 3, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Schlag, M.; Biswas, R.; Krismer, B.; Kohler, T.; Zoll, S.; Yu, W.; Schwarz, H.; Peschel, A.; Gotz, F. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol. Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef]
- Zoll, S.; Schlag, M.; Shkumatov, A.V.; Rautenberg, M.; Svergun, D.I.; Gotz, F.; Stehle, T. Ligand-binding properties and conformational dynamics of autolysin repeat domains in staphylococcal cell wall recognition. J. Bacteriol. 2012, 194, 3789–3802. [Google Scholar] [CrossRef] [Green Version]
- Canepari, P.; Boaretti, M. Lipoteichoic acid as a target for antimicrobial action. Microb. Drug Resist. 1996, 2, 85–89. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batrakov, S.G.; Bergelson, L.D. Lipids of the streptomycetes structural investigation and biological interrelation: A review. Chem. Phys. Lipids 1978, 21, 1–29. [Google Scholar] [CrossRef]
- Da Cunha Camargo, I.L.; Neoh, H.-M.; Cui, L.; Hiramatsu, K. Serial daptomycin selection generates daptomycin-nonsusceptible Staphylococcus aureus strains with a heterogeneous vancomycin-intermediate phenotype. Antimicrob. Agents Chemother. 2008, 52, 4289–4299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, S.A.; White, D.C. Biosynthesis of cardiolipin from phosphatidylglycerol in Staphylococcus aureus. J. Bacteriol. 1972, 109, 820–826. [Google Scholar]
- Ohniwa, R.L.; Kitabayashi, K.; Morikawa, K. Alternative cardiolipin synthase Cls1 compensates for stalled Cls2 function in Staphylococcus aureus under conditions of acute acid stress. FEMS Microbiol. Lett. 2013, 338, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Koprivnjak, T.; Zhang, D.; Ernst, C.M.; Peschel, A.; Nauseef, W.M.; Weiss, J.P. Characterization of Staphylococcus aureus cardiolipin synthases 1 and 2 and their contribution to accumulation of cardiolipin in stationary phase and within phagocytes. J. Bacteriol. 2011, 193, 4134–4142. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.A.; Panesso, D.; McGrath, D.M.; Qin, X.; Mojica, M.F.; Miller, C.; Diaz, L.; Tran, T.T.; Rincon, S.; Barbu, E.M.; et al. Genetic basis for in vivo daptomycin resistance in enterococci. N. Engl. J. Med. 2011, 365, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.; Panesso, D.; Mishra, N.N.; Mileykovskaya, E.; Guan, Z.; Munita, J.M.; Reyes, J.; Diaz, L.; Weinstock, G.M.; Murray, B.E.; et al. Daptomycin-resistant Enterococcus faecalis diverts the antibiotic molecule from the division septum and remodels cell membrane phospholipids. MBio 2013, 4, e00281-13. [Google Scholar] [CrossRef] [Green Version]
- Palmer, K.L.; Daniel, A.; Hardy, C.; Silverman, J.; Gilmore, M.S. Genetic basis for daptomycin resistance in enterococci. Antimicrob. Agents Chemother. 2011, 55, 3345–3356. [Google Scholar] [CrossRef] [Green Version]
- Diaz, L.; Tran, T.T.; Munita, J.M.; Miller, W.R.; Rincon, S.; Carvajal, L.P.; Wollam, A.; Reyes, J.; Panesso, D.; Rojas, N.L.; et al. Whole-genome analyses of Enterococcus faecium isolates with diverse daptomycin MICs. Antimicrob. Agents Chemother. 2014, 58, 4527–4534. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Kelesidis, T.; Tewhey, R.; Rose, W.E.; Schork, N.; Nizet, V.; Sakoulas, G. Genotypic and phenotypic evaluation of the evolution of high-level daptomycin nonsusceptibility in vancomycin-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2012, 56, 6051–6053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelesidis, T.; Tewhey, R.; Humphries, R.M. Evolution of high-level daptomycin resistance in Enterococcus faecium during daptomycin therapy is associated with limited mutations in the bacterial genome. J. Antimicrob. Chemother. 2013, 68, 1926–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.; Panesso, D.; Gao, H.; Roh, J.H.; Munita, J.M.; Reyes, J.; Diaz, L.; Lobos, E.A.; Shamoo, Y.; Mishra, N.N.; et al. Whole-genome analysis of a daptomycin-susceptible enterococcus faecium strain and its daptomycin-resistant variant arising during therapy. Antimicrob. Agents Chemother. 2013, 57, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.; Munita, J.M.; Arias, C.A. Mechanisms of Drug Resistance: Daptomycin Resistance; Blackwell Publishing Inc.: New York, NY, USA, 2015; Volume 1354, pp. 32–53. [Google Scholar]
- Lewis, R.N.; McElhaney, R.N. The physicochemical properties of cardiolipin bilayers and cardiolipin-containing lipid membranes. Biochim. Biophys. Acta 2009, 1788, 2069–2079. [Google Scholar] [CrossRef] [Green Version]
- Pader, V.; Hakim, S.; Painter, K.L.; Wigneshweraraj, S.; Clarke, T.B.; Edwards, A.M. Staphylococcus aureus inactivates daptomycin by releasing membrane phospholipids. Nat. Microbiol. 2016, 2, 16194. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.; Kalamorz, F.; Wecke, T.; Juszczak, A.; Mäder, U.; Homuth, G.; Jordan, S.; Kirstein, J.; Hoppert, M.; Voigt, B.; et al. In-depth profiling of the LiaR response of bacillus subtilis. J. Bacteriol. 2010, 192, 4680–4693. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.N.; McKinnell, J.; Yeaman, M.R.; Rubio, A.; Nast, C.C.; Chen, L.; Kreiswirth, B.N.; Bayer, A.S. In vitro cross-resistance to daptomycin and host defense cationic antimicrobial peptides in clinical methicillin-resistant Staphylococcus aureus isolates. Antimicrob. Agents Chemother. 2011, 55, 4012–4018. [Google Scholar] [CrossRef] [Green Version]
- Howden, B.P.; McEvoy, C.R.E.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.F.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; et al. Evolution of multidrug resistance during Staphylococcus aureus infection involves mutation of the essential two component regulator WalKR. PLoS Pathog. 2011, 7, e1002359. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, H.; Li, S.; Gao, H.; Sun, S.; Li, H.; Wang, R.; Jin, L.; Liu, Y.; Wang, H. Daptomycin resistance in methicillin-resistant Staphylococcus aureus is conferred by IS256 insertion in the promoter of mprF along with mutations in mprF and walK. Int. J. Antimicrob. Agents 2019, 54, 673–680. [Google Scholar] [CrossRef]
- Mishra, N.N.; Bayer, A.S.; Weidenmaier, C.; Grau, T.; Wanner, S.; Stefani, S.; Cafiso, V.; Bertuccio, T.; Yeaman, M.R.; Nast, C.C.; et al. Phenotypic and genotypic characterization of daptomycin-resistant methicillin-resistant Staphylococcus aureus strains: Relative roles of mprF and dlt operons. PLoS ONE 2014, 9, e107426. [Google Scholar] [CrossRef] [Green Version]
- Bertsche, U.; Yang, S.J.; Kuehner, D.; Wanner, S.; Mishra, N.N.; Roth, T.; Nega, M.; Schneider, A.; Mayer, C.; Grau, T.; et al. Increased Cell Wall Teichoic Acid Production and D-alanylation Are Common Phenotypes among Daptomycin-Resistant Methicillin-Resistant Staphylococcus aureus (MRSA) Clinical Isolates. PLoS ONE 2013, 8, e67398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafiso, V.; Bertuccio, T.; Purrello, S.; Campanile, F.; Mammina, C.; Sartor, A.; Raglio, A.; Stefani, S. DltA overexpression: A strain-independent keystone of daptomycin resistance in methicillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2014, 43, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Yang, S.-J.; Bayer, A.S.; Vaezzadeh, A.R.; Herzig, S.; Stenz, L.; Girard, M.; Sakoulas, G.; Scherl, A.; Yeaman, M.R.; et al. Daptomycin resistance mechanisms in clinically derived Staphylococcus aureus strains assessed by a combined transcriptomics and proteomics approach. J. Antimicrob. Chemother. 2011, 66, 1696–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertsche, U.; Weidenmaier, C.; Kuehner, D.; Yang, S.J.; Baur, S.; Wanner, S.; Francois, P.; Schrenzel, J.; Yeaman, M.R.; Bayer, A.S. Correlation of daptomycin resistance in a clinical Staphylococcus aureus strain with increased cell wall teichoic acid production and D-alanylation. Antimicrob. Agents Chemother. 2011, 55, 3922–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-J.; Kreiswirth, B.N.; Sakoulas, G.; Yeaman, M.R.; Xiong, Y.Q.; Sawa, A.; Bayer, A.S. Enhanced expression of dltABCD is associated with the development of daptomycin nonsusceptibility in a clinical endocarditis isolate of Staphylococcus aureus. J. Infect. Dis. 2009, 200, 1916–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keinhorster, D.; George, S.E.; Weidenmaier, C.; Wolz, C. Function and regulation of Staphylococcus aureus wall teichoic acids and capsular polysaccharides. Int. J. Med. Microbiol. 2019, 309, 151333. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Cuirolo, A.X.; Plata, K.B.; Riosa, S.; Silverman, J.A.; Rubio, A.; Rosato, R.R.; Rosato, A.E. VraSR two-component regulatory system contributes to mprF-mediated decreased susceptibility to daptomycin in in vivo-selected clinical strains of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Barros, E.M.; Martin, M.J.; Selleck, E.M.; Lebreton, F.; Sampaio, J.L.M.; Gilmore, M.S. Daptomycin Resistance and Tolerance Due to Loss of Function in Staphylococcus aureus dsp1 and asp23. Antimicrob. Agents Chemother. 2019, 63, e01542-18. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.; Panesso, D.; Tran, T.T.; Mishra, N.N.; Cruz, M.R.; Munita, J.M.; Singh, K.V.; Yeaman, M.R.; Murray, B.E.; Shamoo, Y.; et al. A liaR deletion restores susceptibility to daptomycin and antimicrobial peptides in multidrug-resistant Enterococcus faecalis. J. Infect. Dis. 2015, 211, 1317–1325. [Google Scholar] [CrossRef]
- Munita, J.M.; Panesso, D.; Diaz, L.; Tran, T.T.; Reyes, J.; Wanger, A.; Murray, B.E.; Arias, C.A. Correlation between mutations in liaFSR of Enterococcus faecium and MIC of daptomycin: Revisiting daptomycin breakpoints. Antimicrob. Agents Chemother. 2012, 56, 4354–4359. [Google Scholar] [CrossRef] [Green Version]
- Munita, J.M.; Mishra, N.N.; Alvarez, D.; Tran, T.T.; Diaz, L.; Panesso, D.; Reyes, J.; Murray, B.E.; Adachi, J.A.; Bayer, A.S.; et al. Failure of high-dose daptomycin for bacteremia caused by daptomycin-susceptible Enterococcus faecium harboring LiaSR substitutions. Clin. Infect. Dis. 2014, 59, 1277–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werth, B.J.; Steed, M.E.; Ireland, C.E.; Tran, T.T.; Nonejuie, P.; Murray, B.E.; Rose, W.E.; Sakoulas, G.; Pogliano, J.; Arias, C.A.; et al. Defining daptomycin resistance prevention exposures in vancomycin-resistant Enterococcus faecium and E. faecalis. Antimicrob. Agents Chemother. 2014, 58, 5253–5261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrac, S.; Bisicchia, P.; Devine, K.M.; Msadek, T. A matter of life and death: Cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway. Mol. Microbiol. 2008, 70, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Takada, H.; Yoshikawa, H. Essentiality and function of WalK/WalR two-component system: The past, present, and future of research. Biosci. Biotechnol. Biochem. 2018, 82, 741–751. [Google Scholar] [CrossRef]
- Salzberg, L.I.; Powell, L.; Hokamp, K.; Botella, E.; Noone, D.; Devine, K.M. The WalRK (YycFG) and sigma(I) RsgI regulators cooperate to control CwlO and LytE expression in exponentially growing and stressed Bacillus subtilis cells. Mol. Microbiol. 2013, 87, 180–195. [Google Scholar] [CrossRef]
- Mohedano, M.L.; Amblar, M.; de la Fuente, A.; Wells, J.M.; Lopez, P. The Response Regulator YycF Inhibits Expression of the Fatty Acid Biosynthesis Repressor FabT in Streptococcus pneumoniae. Front. Microbiol. 2016, 7, 1326. [Google Scholar] [CrossRef] [Green Version]
- Beranová, J.; Jemioła-Rzemińska, M.; Elhottová, D.; Strzałka, K.; Konopásek, I.; Beranova, J.; Jemiola-Rzeminska, M.; Elhottova, D.; Strzalka, K.; Konopasek, I. Metabolic control of the membrane fluidity in Bacillus subtilis during cold adaptation. Biochim. Biophys. Acta-Biomembr. 2008, 1778, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Bacterial sensing of antimicrobial peptides. Contrib. Microbiol. 2009, 16, 136–149. [Google Scholar]
- Jordan, S.; Junker, A.; Helmann, J.D.; Mascher, T. Regulation of LiaRS-dependent gene expression in Bacillus subtilis: Identification of inhibitor proteins, regulator binding sites, and target genes of a conserved cell envelope stress-sensing two-component system. J. Bacteriol. 2006, 188, 5153–5166. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Escobar, J.; Wolf, D.; Fritz, G.; Hofler, C.; Wedlich-Soldner, R.; Mascher, T. Subcellular localization, interactions and dynamics of the phage-shock protein-like Lia response in Bacillus subtilis. Mol. Microbiol. 2014, 92, 716–732. [Google Scholar] [CrossRef]
- Kingston, A.W.; Liao, X.; Helmann, J.D. Contributions of the sigma(W), sigma(M) and sigma(X) regulons to the lantibiotic resistome of Bacillus subtilis. Mol. Microbiol. 2013, 90, 502–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brötz-Oesterhelt, H.; Brunner, N.A. How many modes of action should an antibiotic have? Curr. Opin. Pharmacol. 2008, 8, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Robbel, L.; Marahiel, M.A. Daptomycin, a bacterial lipopeptide synthesized by a nonribosomal machinery. J. Biol. Chem. 2010, 285, 27501–27508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Sheng, J.; Huang, G.; Ma, R.; Yin, F.; Song, D.; Zhao, C.; Ma, S. Design, synthesis and antibacterial activity of cinnamaldehyde derivatives as inhibitors of the bacterial cell division protein FtsZ. Eur. J. Med. Chem. 2015, 97, 32–41. [Google Scholar] [CrossRef]
- Daley, P.; Louie, T.; Lutz, J.E.; Khanna, S.; Stoutenburgh, U.; Jin, M.; Adedoyin, A.; Chesnel, L.; Guris, D.; Larson, K.B.; et al. Surotomycin versus vancomycin in adults with Clostridium difficile infection: Primary clinical outcomes from the second pivotal, randomized, double-blind, Phase 3 trial. J. Antimicrob. Chemother. 2017, 72, 3462–3470. [Google Scholar] [CrossRef]
- Boix, V.; Fedorak, R.N.; Mullane, K.M.; Pesant, Y.; Stoutenburgh, U.; Jin, M.; Adedoyin, A.; Chesnel, L.; Guris, D.; Larson, K.B.; et al. Primary Outcomes From a Phase 3, Randomized, Double-Blind, Active-Controlled Trial of Surotomycin in Subjects With Clostridium difficile Infection. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2017; Volume 4. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, D.A.; Wenzel, M. More Than a Pore: A Current Perspective on the In Vivo Mode of Action of the Lipopeptide Antibiotic Daptomycin. Antibiotics 2020, 9, 17. https://doi.org/10.3390/antibiotics9010017
Gray DA, Wenzel M. More Than a Pore: A Current Perspective on the In Vivo Mode of Action of the Lipopeptide Antibiotic Daptomycin. Antibiotics. 2020; 9(1):17. https://doi.org/10.3390/antibiotics9010017
Chicago/Turabian StyleGray, Declan Alan, and Michaela Wenzel. 2020. "More Than a Pore: A Current Perspective on the In Vivo Mode of Action of the Lipopeptide Antibiotic Daptomycin" Antibiotics 9, no. 1: 17. https://doi.org/10.3390/antibiotics9010017
APA StyleGray, D. A., & Wenzel, M. (2020). More Than a Pore: A Current Perspective on the In Vivo Mode of Action of the Lipopeptide Antibiotic Daptomycin. Antibiotics, 9(1), 17. https://doi.org/10.3390/antibiotics9010017