Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus

 ,
, 
 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Characterization of MRSA Biofilm Producers
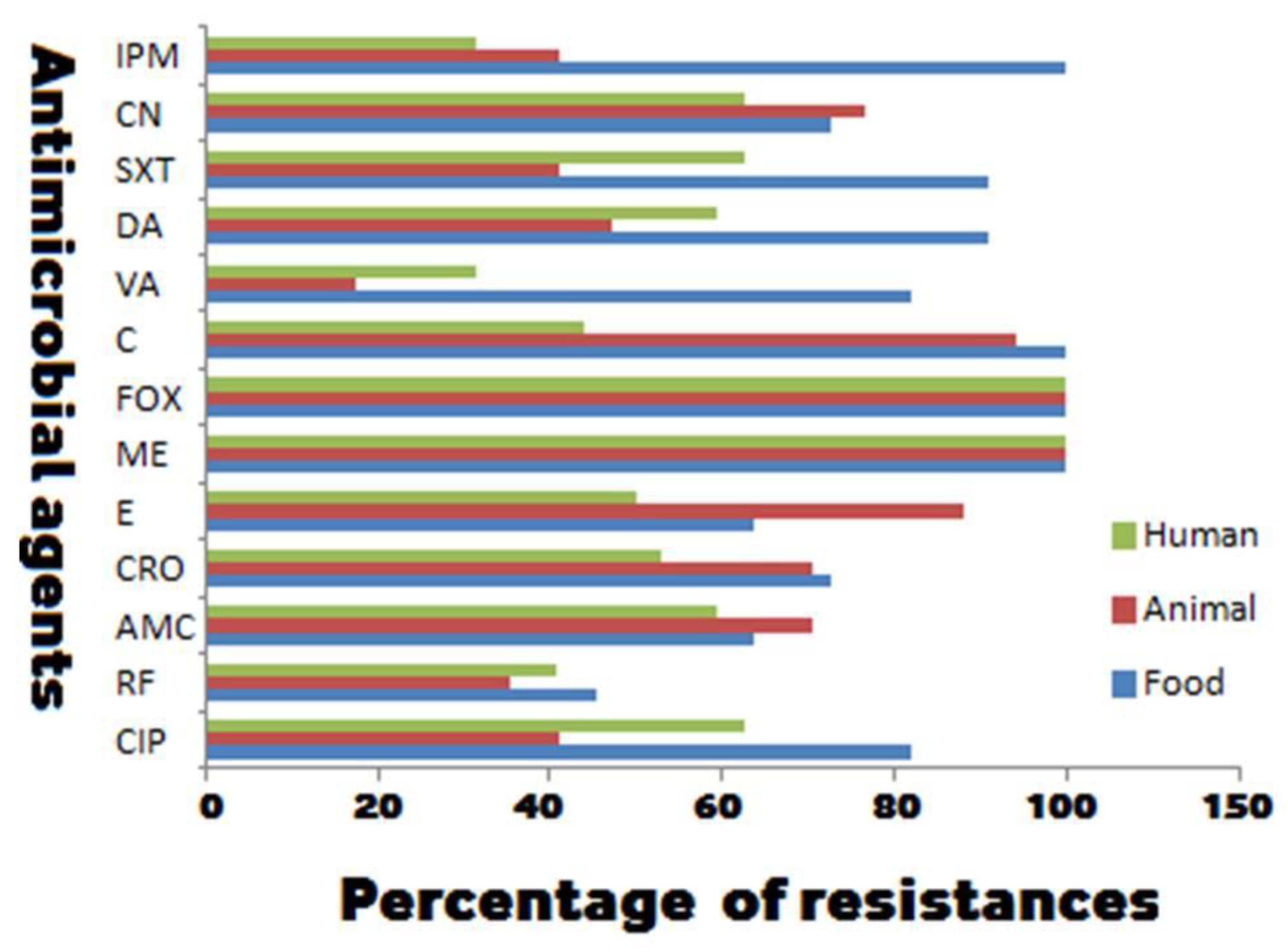
2.2. Antibiogram of Biofilm Producing and Non-Producing MRSA
2.3. Genetic Characterization of MRSA Isolates
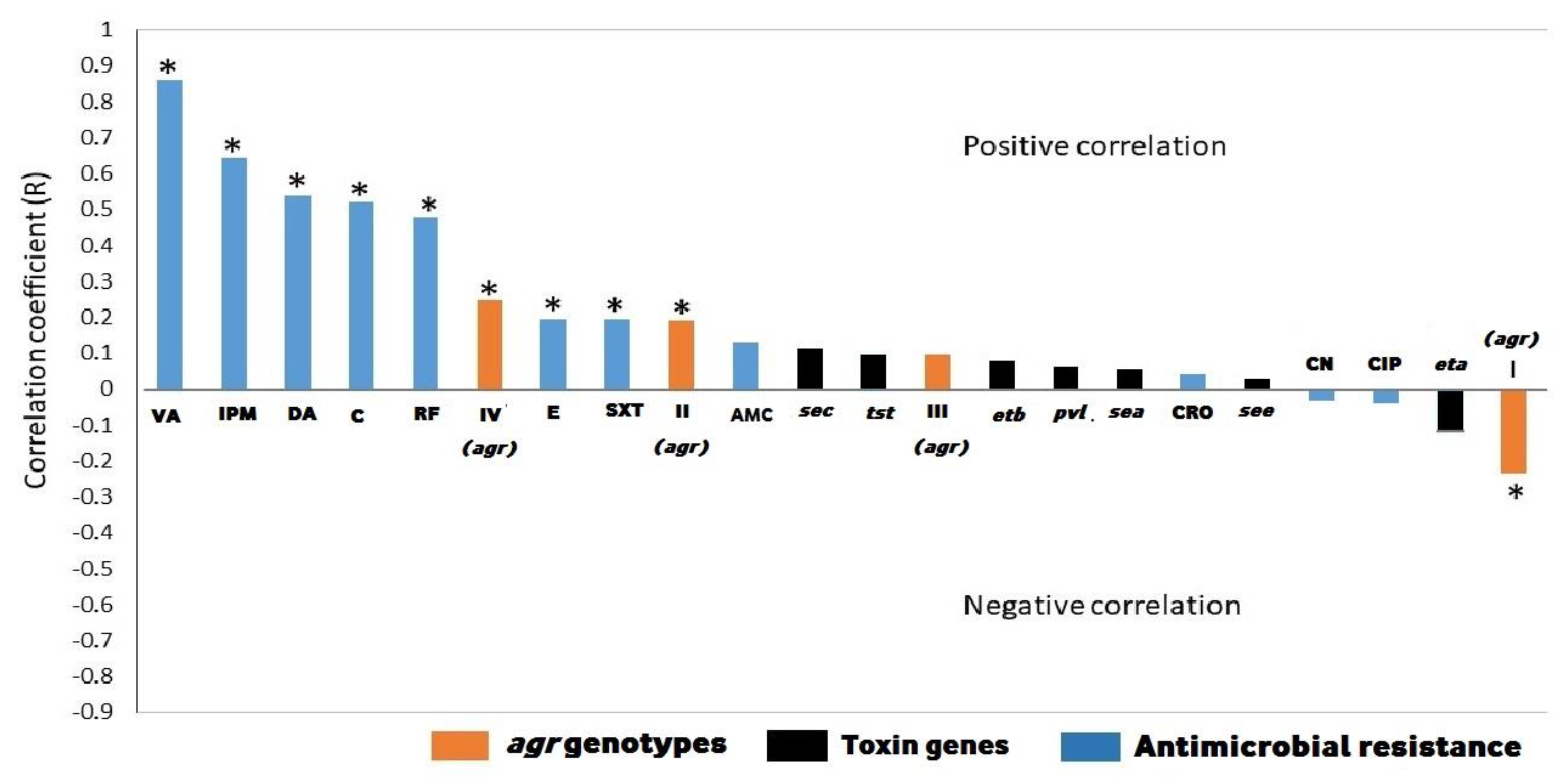
2.4. Correlation Between Biofilm Production and Antimicrobial Resistance, Toxin Genes, and agr Genotypes
2.5. Phenotypic Analysis of Antibiofilm Activities of Zinc Oxide Nanoparticles (ZnO NPs), Proteinase K, and Hamamelitannin (HAM)
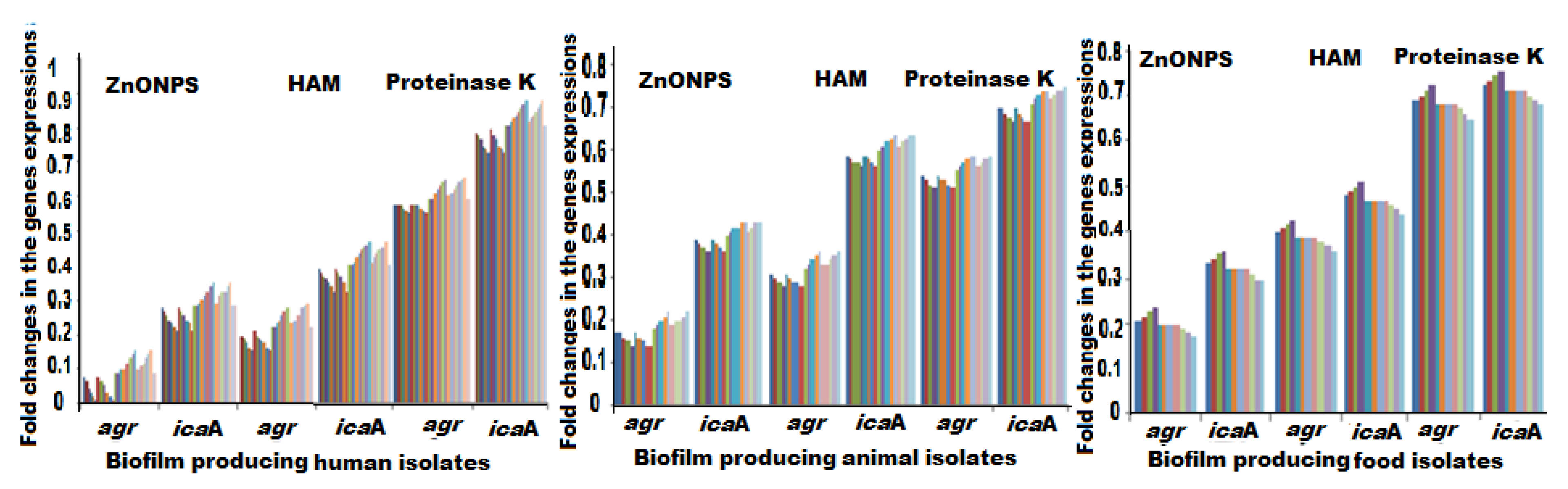
2.6. Biofilm Associated Genes Expressions Post Exposure to the Antibiofilm Agents
2.7. Antimicrobial Activities of ZnO NPs, Proteinase K, and HAM
3. Discussion
4. Materials and Methods
4.1. Ethical Statements
4.2. Isolation and Identification of MRSA
4.3. Detection of Biofilm Producing MRSA
4.4. Antimicrobial Susceptibility Testing of MRSA
4.5. Toxin Genes Profiles and agr Alleles of MRSA Isolates
4.6. Assessment of Antibiofilm and Antimicrobial Activities of Promising Agents
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Hamid, M.I.; Bendary, M.M. Comparative phenotypic and genotypic discrimination of methicillin resistant and susceptible Staphylococcus aureus in Egypt. Cell. Mol. Biol. 2015, 61, 106–117. [Google Scholar]
- Abd El-Hamid, M.I.; Bendary, M.M.; Merwad, A.M.; Elsohaby, I.; Ghaith, D.; Alshareef, W.A. What is behind phylogenetic analysis of hospital-, community- and livestock-associated methicillin-resistant Staphylococcus aureus? Transbound. Emerg. Dis. 2019, 66, 1506–1517. [Google Scholar] [CrossRef]
- Laverty, G.; Gorman, S.P.; Gilmore, B.F. Biomolecular mechanisms of staphylococcal biofilm formation. Future Microbiol. 2013, 8, 509–524. [Google Scholar] [CrossRef] [Green Version]
- Van-Acker, H.; Van-Dijck, P.; Coenye, T. Molecular mechanisms of antimicrobial tolerance and resistance in bacterial and fungal biofilms. Trends Microbiol. 2014, 22, 326–333. [Google Scholar] [CrossRef]
- McCarthy, H.; Rudkin, J.K.; Black, N.S.; Gallagher, L.; O’Neill, E.; O’Gara, J.P. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2015, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Römling, U.; Balsalobre, C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern. Med. 2012, 272, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Multidrug tolerance of biofilms and persister cells. Curr. Top. Microbiol. Immunol. 2008, 322, 107–131. [Google Scholar] [PubMed]
- Otto, M. Physical stress and bacterial colonization. FEMS Microbiol. Rev. 2014, 38, 1250–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell. Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Manso, D.; del-Prado, G.; Ortiz-Pérez, A.; Manrubia-Cobo, M.; Gómez-Barrena, E.; Cordero-Ampuero, J.; Esteban, J. In vitro susceptibility to antibiotics of staphylococci in biofilms isolated from orthopaedic infections. Int. J. Antimicrob. Agents 2013, 41, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Trentin, D.S.; Giordani, R.B.; Macedo, A.J. Biofilmes bacterianos patogênicos: Aspectos gerais, importância clínica e estratégias de combate. Rev. Liberato 2013, 14, 113–238. [Google Scholar]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Baig, M.H. Eugenol inhibits quorum sensing and biofilm of toxigenic MRSA strains isolated from food handlers employed in Saudi Arabia. Biotechnol. Biotechnol. Equip. 2017, 31, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [Green Version]
- Kiran, M.D.; Adikesavan, N.V.; Cirioni, O.; Giacometti, A.; Silvestri, C.; Scalise, G.; Ghiselli, R.; Saba, V.; Orlando, F.; Shoham, M.; et al. Discovery of a quorum-sensing inhibitor of drug-resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008, 73, 1578–1586. [Google Scholar] [CrossRef] [Green Version]
- Loughran, A.J.; Atwood, D.N.; Anthony, A.C.; Harik, N.S.; Spencer, H.J.; Beenken, K.E.; Smeltzer, M.S. Impact of individual extracellular proteases on Staphylococcus aureus biofilm formation in diverse clinical isolates and their isogenic sarA mutants. Microbiol. Open 2014, 3, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Smijs, T.G.; Pavel, S. Titanium dioxide and zinc oxide nanoparticles in sunscreens: Focus on their safety and effectiveness. Nanotechnol. Sci. Appl. 2011, 4, 95–112. [Google Scholar] [CrossRef] [Green Version]
- Ashajyothi, C.; Harish, K.H.; Dubey, N.; Chandrakanth, R.K. Antibiofilm activity of biogenic copper and zinc oxide nanoparticles-antimicrobials collegiate against multiple drug resistant bacteria: A nanoscale approach. J. Nanostruct. Chem. 2016, 6, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Ghaly, M.F.; Shaheen, A.; Bouhy, A.; Bendary, M.M. Alternative therapy to manage otitis media caused by multidrug-resistant fungi. Arch. Microbiol. 2020, 202, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Neopane, P.; Nepal, H.P.; Shrestha, R.; Uehara, O.; Abiko, Y. In vitro biofilm formation by Staphylococcus aureus isolated from wounds of hospital-admitted patients and their association with antimicrobial resistance. Int. J. Gen. Med. 2018, 11, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Algburi, A.; Comito, N.; Kashtanov, D.; Dicks, L.M.; Chikindasc, M.L. Control of biofilm formation: Antibiotics and beyond. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diane, C.B.; Marlène, Z.L.; Pierre-Louis, T.; Frédérique, W.; Farid, E.G.; Alain, B.; Aude, A.F. Differential activity of the combination of vancomycin and amikacin on planktonic vs. biofilm-growing Staphylococcus aureus bacteria in a hollow fiber infection model. Front. Microbiol. 2018, 9, 572. [Google Scholar]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Role of persisters and small-colony variants in antibiotic resistance of planktonic and biofilm associated Staphylococcus aureus: An in vitro study. J. Med. Microbiol. 2009, 58, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebeaux, D.; Leflon-Guibout, V.; Ghigo, J.M.; Beloin, C. In vitro activity of gentamicin, vancomycin or amikacin combined with EDTA or l-arginine as lock therapy against a wide spectrum of biofilm-forming clinical strains isolated from catheter-related infections. J. Antimicrob. Chemother. 2015, 70, 1704–1712. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Jiang, B.; Jia, C.; Wu, H.; Shen, J.; Hu, X.; Xie, Z. Investigation of bioflm production and its association with genetic and phenotypic characteristics of OM (osteomyelitis) and non-OM orthopedic Staphylococcus aureus. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 10. [Google Scholar] [CrossRef] [Green Version]
- Paharik, A.E.; Horswill, A.R. The staphylococcal biofilm: Adhesins, regulation, and host response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Cheung, G.Y.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Manago, K.; Nishi, J.; Wakimoto, N.; Miyanohara, H.; Sarantuya, J.; Tokuda, K.; Iwashita, M. Biofilm formation by accessory gene regulator typing of methicillin resistant S. aureus strains recovered from patients with nosocomial infections. Infect. Control Hosp. Epidemiol. 2006, 27, 188–190. [Google Scholar] [CrossRef]
- Yoon, H.J.; Choi, J.Y.; Lee, K.; Yong, D.; Kim, J.M.; Song, Y.G. Accessory gene regulator group polymorphisms in MRSA: An association with clinical significance. Yonsei Med. J. 2007, 4, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Bardiau, M.; Detilleux, J.; Farnir, F.; Mainil, J.G.; Ote, I. Associations between properties linked with persistence in a collection of Staphylococcus aureus isolates from bovine mastitis. Vet. Microbiol. 2014, 169, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoramrooz, S.S.; Mansouri, F.; Marashifard, M.; Hosseini, S.A.; Akbarian, C.F.; Ganavehei, B.; Gharibpour, F.; Shahbazi, A.; Mirzaii, M.; Darban, S.D. Detection of biofilm related genes, classical enterotoxin genes and agr typing among Staphylococcus aureus isolated from bovine with subclinical mastitis in southwest of Iran. Microb. Pathog. 2016, 97, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.; Ramon, E.; Hoyo, J.; Tzanov, T. Innovative approaches for controlling clinically relevant biofilms: Current trends and future prospects. Curr. Top. Med. Chem. 2017, 17, 1889–1914. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Malaikozhundan, B.; Shanthi, S.; Vaseeharan, B.; Thajuddin, N. Control of biofilm forming clinically important bacteria by green synthesized ZnO nanoparticles and its ecotoxicity on Ceriodaphnia cornuta. Microb. Pathog. 2015, 107, 88–97. [Google Scholar] [CrossRef]
- Mahamuni, P.P.; Patil, P.M.; Dhanavade, M.J.; Badiger, M.V.; Shadija, P.G.; Lokhande, A.C.; Bohara, R.A. Synthesis and characterization of zinc oxide nanoparticles by using polyol chemistry for their antimicrobial and antibiofilm activity. Biochem. Biophys. Rep. 2019, 17, 71–80. [Google Scholar] [CrossRef]
- Shukla, K.S.; Rao, S.T. Staphylococcus aureus biofilm removal by targeting biofilm-associated extracellular protein. J. Antibiot. 2017, 66, 55–60. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.G.; Cho, M.H.; Lee, J. ZnO nanoparticles inhibit Peudomonas aeruginosa biofilm formation and virulence factor production. Microbiol. Res. 2014, 169, 888–896. [Google Scholar] [CrossRef]
- Shakerimoghaddam, A.; Razavi, D.; Rahvar, F.; Khurshid, M.; Mogharabi Ostadkelayeh, S.; Esmaeili, S.A.; Khaledi, A.; Eshraghi, M. Evaluate the effect of zinc oxide and silver nanoparticles on biofilm and icaA gene expression in methicillin-resistant Staphylococcus aureus isolated from burn wound infection. J. Burn Care Res. 2020, iraa085. [Google Scholar] [CrossRef]
- Roxanne, M. Effect of hamamelitannin on Staphylococcus aureus biofilm susceptibility. Vakgroep FW02. Master’s Thesis, Ghent University, Ghent, Belgium, 2015. Available online: https://lib.ugent.be/catalog/rug01:002217472 (accessed on 1 October 2020).
- Boles, B.R.; Horswill, A.R. agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Breyne, K.; Rycke, R.; Vermote, A.; Van Nieuwerburgh, F.; Meyer, E.; Van Calenbergh, S.; Coenye, T. The quorum sensing inhibitor hamamelitannin increases antibiotic susceptibility of S. aureus biofilms by affecting peptidoglycan biosynthesis and DNA release. Sci. Rep. 2016, 6, 20321. [Google Scholar] [CrossRef] [PubMed]
- Namasivayam, S.; Prasanna, M.; Subathra, S. Synergistic antibacterial activity of zinc oxide nanoparticles with antibiotics against human pathogenic bacteria. J. Chem. Pharm. Res. 2015, 7, 133–138. [Google Scholar]
- Becker, K.; Skov, R.L.; von Eiff, C. Staphylococcus, Micrococcus, and other catalase-positive cocci. In Manual of Clinical Microbiology; Jorgensen, J.H., Pfaller, M.A., Carroll, K.C., Funke, G., Landry, M.L., Richter, S.S., Warnock, D.W., Eds.; American Society for Microbiology: Washington, DC, USA, 2015; pp. 354–382. [Google Scholar]
- Mason, W.J.; Blevins, J.S.; Beenken, K.; Wibowo, N.; Ojha, N.; Smeltzer, M.S. Multiplex PCR protocol for the diagnosis of staphylococcal infection. J. Clin. Microbiol. 2001, 39, 3332–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase-negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savie, B.; Svabic-Vlahovic, M. A modified microtiter plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Ciftci, A.; Findik, A.; Onuk, A.; Savasan, S. Detection of methicillin resistance and slime factor prouction of Staphylococus aureus in bovine mastitis. Braz. J. Microbiol. 2009, 40, 254–261. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Approved Guideline (M45-A); Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Bendary, M.M.; Solyman, S.M.; Azab, M.M.; Mahmoud, N.F.; Hanora, A.M. Genetic diversity of multidrug resistant Staphylococcus aureus isolated from clinical and non clinical samples in Egypt. Cell. Mol. Biol. 2016, 62, 55. [Google Scholar]
- Franklin, R.C.; Patricia, A.B.; George, M.E.; Janet, A.H.; Stephen, G.J.; James, S.L.; Brandi, L. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement, M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins exfoliative toxins toxic shock syndrome toxin 1 methicillin resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [CrossRef] [Green Version]
- Melake, N.A.; Zakaria, A.S.; Ibrahim, N.H.; Salma, M.A.; Mahmoud, A.Z. Prevalence of agr specificity groups among in vitro biofilm forming methiciilin resistance Staphylococcus aureus strains isolated from nasal carriers. Int. J. Microbiol. Res. 2014, 5, 76–84. [Google Scholar]
- Marquenie, R. Effect of HAM on S. aureus Biofilm Susceptibility. Master’s Thesis, Ghent University, Ghent, Belgium, 2015. [Google Scholar]
- Mirani, Z.A.; Jamil, N. Effect of sub-lethal doses of vancomycin and oxacillin on biofilm formation by vancomycin intermediate resistant Staphylococcus aureus. J. Basic Microbiol. 2011, 51, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.A.; Dos Santos, T.F.; de Almeida, M.E.; Junior, L.A.; Andrade, E.F.; Rezende, R.P.; Marques, L.M.; Romano, C.C. Inhibition of Staphylococcus aureus biofilm by Lactobacillus isolated from fine cocoa. BMC Microbiol. 2016, 16, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, A.A. Antibacterial activity of zinc oxide nanoparticles against toxigenic Bacillus cereus and S. aureus isolated from some Egyptian food. Int. J. Microbiol. Res. 2015, 6, 145–154. [Google Scholar]
- Friendly, M. Corrgrams. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]
- Frank, E.H., Jr. Package ‘Hmisc’. 2020. Available online: https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (accessed on 11 August 2020).




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Resistance/agr and Toxin Genes | No. (%) of MRSA Stains from Various Sources with Different Biofilm Phenotypes | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Non-Biofilm Producers | Biofilm Producers | ||||||||
| Human (84) * | Animal (49) * | Food (29) * | Total (162) * | Human (32) * | Animal (17) * | Food (11) * | Total (60) * | ||
| Antimicrobial resistance ** | CIP | 46 (54.8) | 20 (41) | 13 (44.8) | 79 (48.8) | 20 (62.5) | 7 (41.2) | 9 (81.8) | 36 (60) |
| RF | 15 (17.9) | 9 (18.4) | 7 (24.1) | 31 (19.1) | 13 (40.6) | 6 (35.3) | 5 (45.5) | 24 (40) | |
| AMC | 38 (45.2) | 22 (44.9) | 17 (58.6) | 77 (47.5) | 19 (59.4) | 12 (70.6) | 7 (63.6) | 38 (63.3) | |
| CRO | 38 (45.2) | 22 (44.9) | 17 (58.6) | 77 (47.5) | 17 (53.1) | 12 (70.6) | 8 (72.7) | 37 (61.7) | |
| E | 33 (39.3) | 19 (38.8) | 16 (55.2) | 68 (42) | 16 (50) | 15 (88.2) | 7 (63.6) | 38 (63.3) | |
| C | 9 (10.7) | 5 (10.2) | 3 (10.3) | 17 (10.5) | 14 (43.8) | 16 (94.1) | 11 (100) | 41 (68.3) | |
| VA | 9 (10.7) | 2 (4.1) | 2 (6.9) | 13 (8) | 10 (31.3) | 3 (17.6) | 9 (81.8) | 22 (36.7) | |
| DA | 33 (39.3) | 23 (46.9) | 9 (31) | 65 (40.1) | 19 (59.4) | 8 (47.1) | 10 (90.9) | 37 (61.7) | |
| SXT | 39 (46.4) | 18 (36.7) | 11 (37.9) | 68 (42) | 20 (62.5) | 7 (41.2) | 10 (90.9) | 37 (61.7) | |
| CN | 40 (47.6) | 19 (38.8) | 10 (34.5) | 69 (42.6) | 20 (62.5) | 13 (76.5) | 8 (72.7) | 41 (68.3) | |
| IPM | 5 (6) | 1 (2) | 0 (0) | 6 (3.7) | 10 (31.3) | 7 (41.2) | 11 (100) | 28 (46.7) | |
| agr alleles | agr I | 74 (88.1) | 34 (69.4) | 18 (62.1) | 126 (77.8) | 22 (68.8) | 8 (47.1) | 9 (81.8) | 39 (65) |
| agr II | 0 (0) | 4 (8.2) | 2 (6.9) | 6 (3.7) | 0 (0) | 2 (11.8) | 1 (9.1) | 3 (5) | |
| agr III | 10 (11.9) | 6 (12.2) | 6 (20.7) | 22 (13.6) | 7 (21.9) | 5 (29.4) | 1 (9.1) | 13 (21.7) | |
| agr IV | 0 (0) | 5 (10.2) | 3 (10.3) | 8 (4.9) | 0 (0) | 2 (11.8) | 3 (27.3) | 5 (8.3) | |
| Toxin genes | pvl | 36 (42.9) | 25 (51) | 16 (55.2) | 77 (47.5) | 16 (50) | 12 (70.6) | 4 (36.4) | 32 (53.3) |
| sea | 28 (33.3) | 25 (51) | 12 (41.4) | 65 (40.1) | 16 (50) | 9 (52.9) | 3 (27.3) | 28 (46.7) | |
| see | 11 (13.1) | 5 (10.2) | 3 (10.3) | 19 (11.7) | 4 (12.5) | 3 (17.6) | 1 (9.1) | 8 (13.3) | |
| sec | 10 (11.9) | 1 (2) | 1 (3.4) | 12 (7.4) | 4 (12.5) | 0 (0) | 4 (36.4) | 8 (13.3) | |
| tst | 7 (8.3) | 5 (10.2) | 3 (10.3) | 15 (9.3) | 6 (18.8) | 3 (17.6) | 1 (9.1) | 10 (16.7) | |
| eta | 10 (11.9) | 9 (18.4) | 7 (24.1) | 26 (16) | 2 (6.3) | 2 (11.8) | 0 (0) | 4 (6.7) | |
| etb | 5 (6) | 3 (6.1) | 5 (17.2) | 13 (8) | 4 (12.5) | 3 (17.6) | 1 (9.1) | 8 (13.3) | |
| Antibiofilm Activities | Promising | MRSA Isolates Sources (No.) | |||
|---|---|---|---|---|---|
| Antibiofilm Agents | Human (32) | Animal (17) | Food (11) | ||
| Reduction percentage of biofilm capacity * (Phenotypically, MTP method) | ZnO NPs | 99.79 ± 0.089 | 99.81 ± 0.088 | 99.58 ± 0.016 | |
| HAM | 99.58 ± 0.072 | 99.81 ± 0.007 | 99.58 ± 0.016 | ||
| Proteinase K | 99.51 ± 0.065 | 99.28 ± 0.013 | 99.44 ± 0.004 | ||
| Fold changes of biofilm and QS genes expressions | icaA | ZnO NPs | 0.28 ± 0.042 | 0.18 ± 0.027 | 0.15 ± 0.020 |
| HAM | 0.40 ± 0.042 | 0.32 ± 0.027 | 0.45 ± 0.020 | ||
| Proteinase K | 0.81 ± 0.045 | 0.55 ± 0.027 | 0.69 ± 0.020 | ||
| agr | ZnO NPs | 0.08 ± 0.042 | 0.40 ± 0.025 | 0.48 ± 0.020 | |
| HAM | 0.22 ± 0.042 | 0.60 ± 0.027 | 0.59 ± 0.020 | ||
| Proteinase K | 0.59 ± 0.032 | 0.71 ± 0.027 | 0.82 ± 0.020 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Hamid, M.I.; Y. El-Naenaeey, E.-s.; M kandeel, T.; Hegazy, W.A.H.; Mosbah, R.A.; Nassar, M.S.; Bakhrebah, M.A.; Abdulaal, W.H.; Alhakamy, N.A.; Bendary, M.M. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 667. https://doi.org/10.3390/antibiotics9100667
Abd El-Hamid MI, Y. El-Naenaeey E-s, M kandeel T, Hegazy WAH, Mosbah RA, Nassar MS, Bakhrebah MA, Abdulaal WH, Alhakamy NA, Bendary MM. Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2020; 9(10):667. https://doi.org/10.3390/antibiotics9100667
Chicago/Turabian StyleAbd El-Hamid, Marwa I., El-sayed Y. El-Naenaeey, Toka M kandeel, Wael A. H. Hegazy, Rasha A. Mosbah, Majed S. Nassar, Muhammed A. Bakhrebah, Wesam H. Abdulaal, Nabil A. Alhakamy, and Mahmoud M. Bendary. 2020. "Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus" Antibiotics 9, no. 10: 667. https://doi.org/10.3390/antibiotics9100667
APA StyleAbd El-Hamid, M. I., Y. El-Naenaeey, E. -s., M kandeel, T., Hegazy, W. A. H., Mosbah, R. A., Nassar, M. S., Bakhrebah, M. A., Abdulaal, W. H., Alhakamy, N. A., & Bendary, M. M. (2020). Promising Antibiofilm Agents: Recent Breakthrough against Biofilm Producing Methicillin-Resistant Staphylococcus aureus. Antibiotics, 9(10), 667. https://doi.org/10.3390/antibiotics9100667