Fragments of a Wheat Hevein-Like Antimicrobial Peptide Augment the Inhibitory Effect of a Triazole Fungicide on Spore Germination of Fusarium oxysporum and Alternaria solani

 , , and
, , and
Abstract
:1. Introduction
2. Results
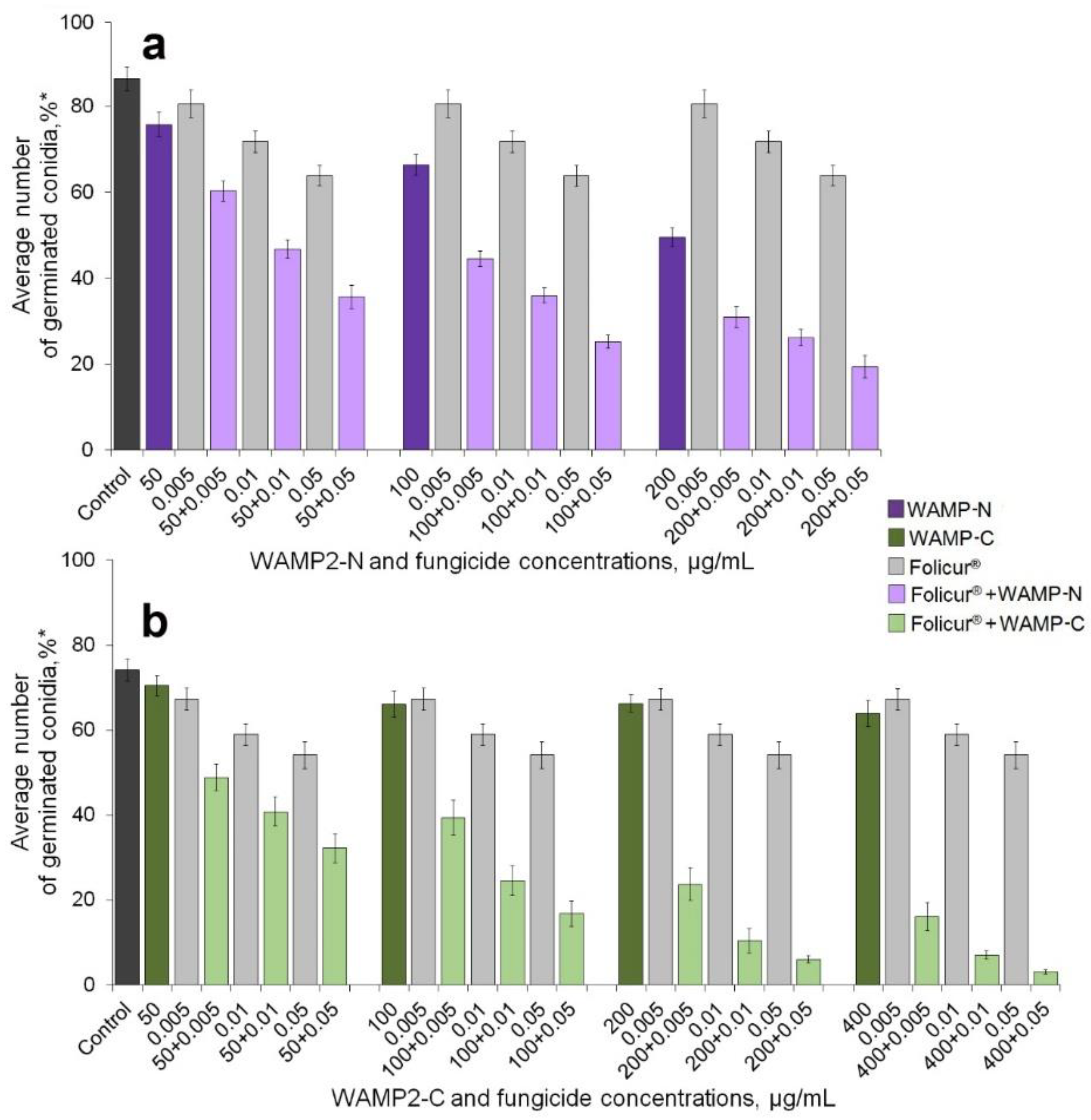
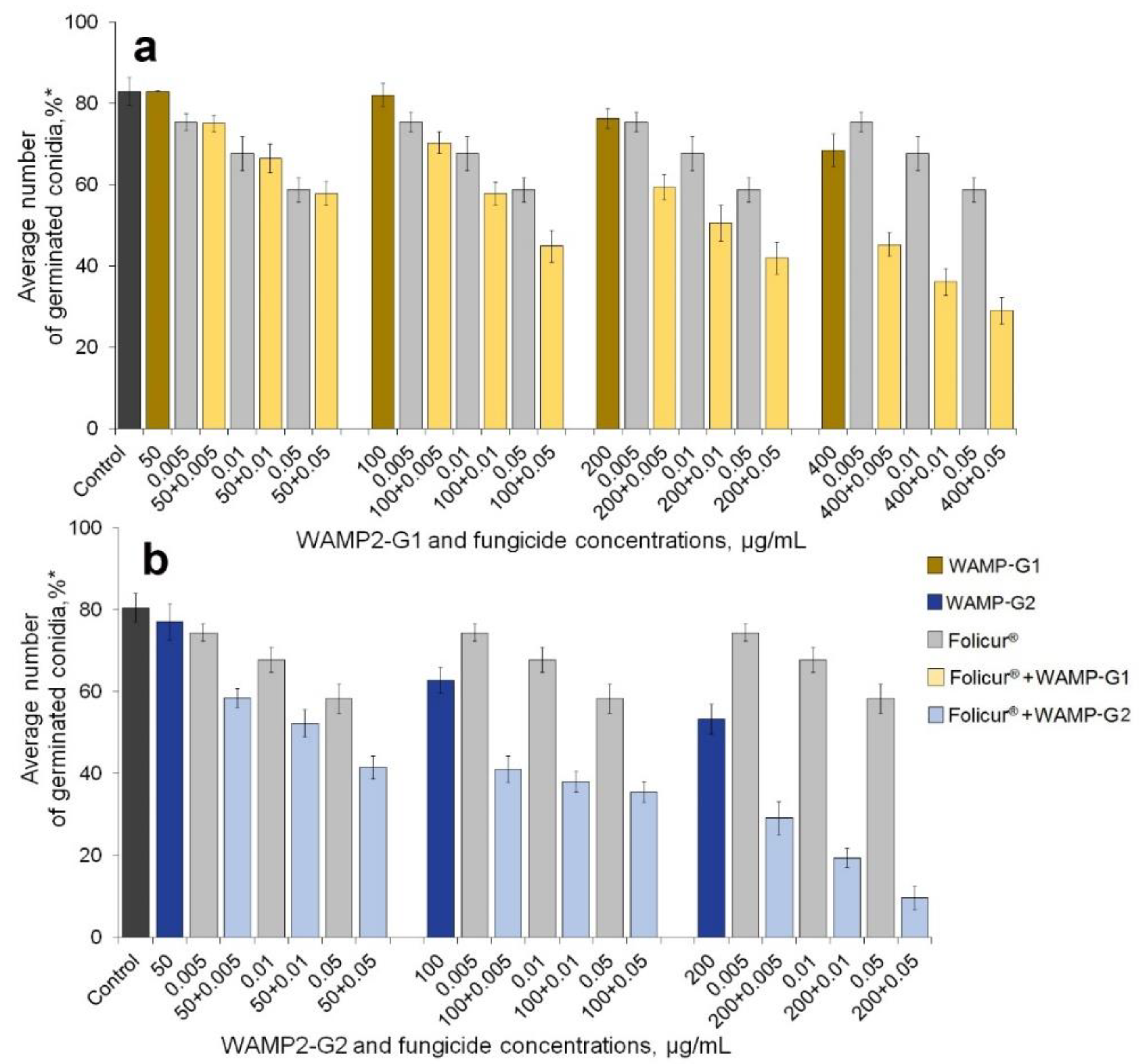
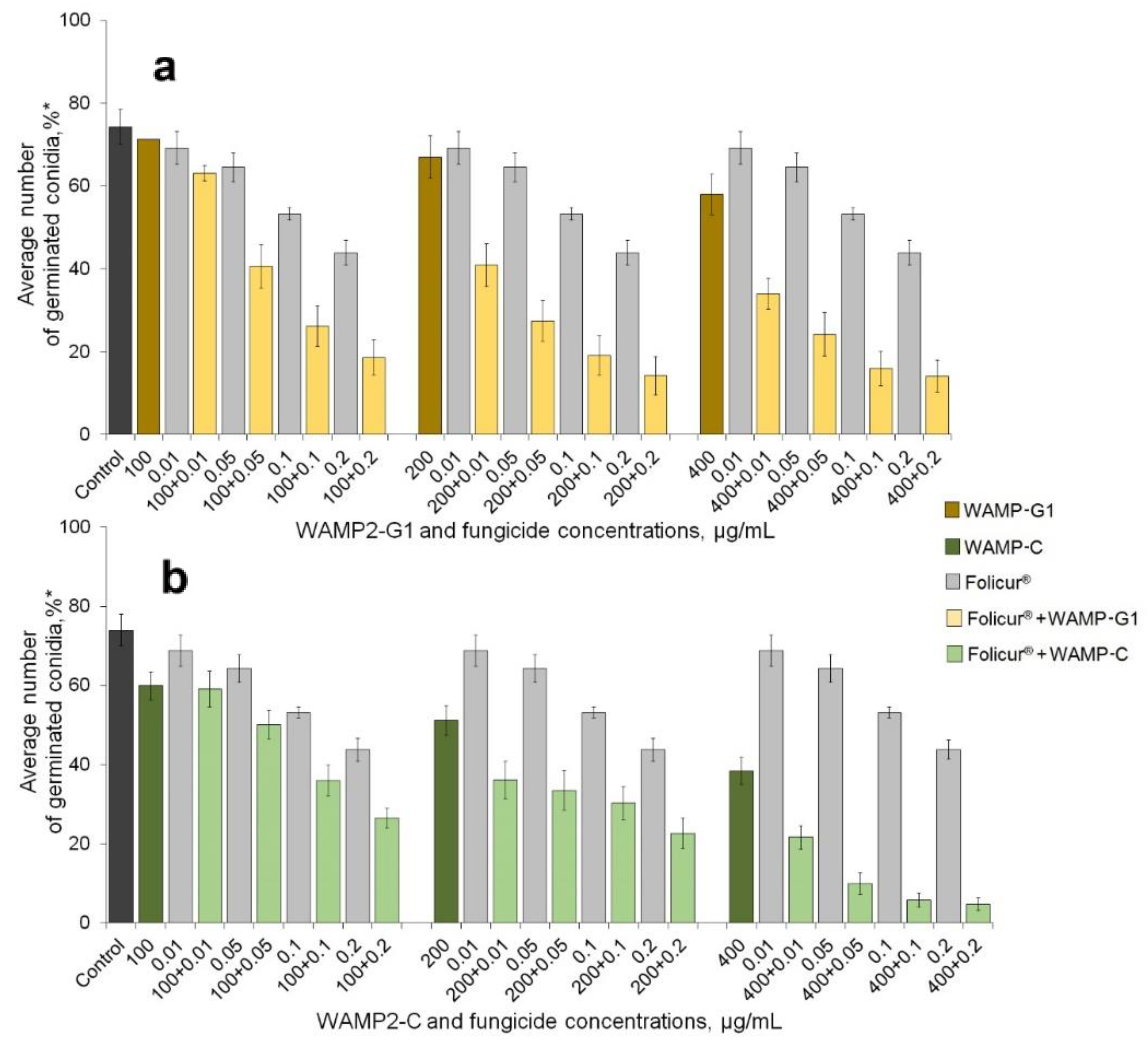
2.1. Enhancement of the Fungicidal Effect by Folucur® EC 250 by Co-Application with WAMP-2 Fragments In Vitro
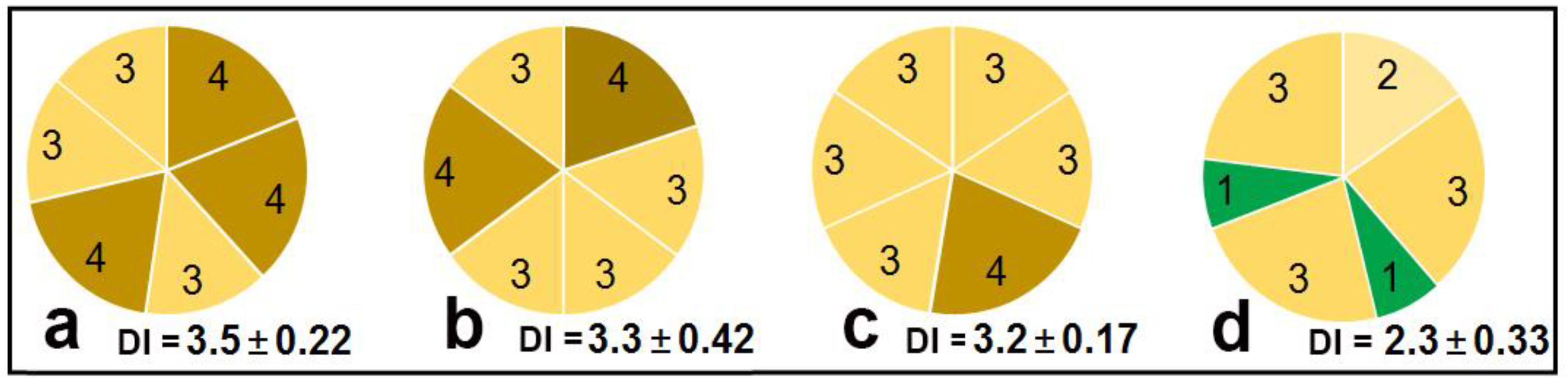
2.2. Study of the Sensitizing Activity of WAMP-2 Fragments Using a Wilting Test on Detached Seedlings and a Leaf Disk Assay
3. Discussion
4. Materials and Methods
4.1. Tomato Pathogens
4.2. Production of WAMP-2-Derived-Peptides and WAMP-2
4.3. Fungicide
4.4. In Vitro Assay of Inhibitory and Sensitizing Activities
4.5. Detached Seedling Wilting Test
4.6. Tomato Leaf Disk Assay
4.7. Data Analysis and Statistical Treatment
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Commodity Production. 2012. Available online: http://faostat.fao.org/site/339/default.aspx (accessed on 11 May 2013).
- Foolad, M.R.; Sharma, A.; Ashrafi, H.; Lin, G. Genetics and breeding of early blight resistance in tomato. Hort. Science 2005, 40, 1114C-1114. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.B.; Zitter, T.A.; Momol, T.M.; Miller, S.A. (Eds.) Compendium of Tomato Diseases and Pests, 2nd ed.; APS Press: St. Paul, MN, USA, 2014; pp. 1–168. ISBN 978-0-89054-434-1. [Google Scholar]
- Fravel, D.; Olivain, C.; Alabouvette, C. Fusarium oxysporum and its biocontrol. New Phytol. 2003, 157, 493–502. [Google Scholar] [CrossRef]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, A.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici, causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef]
- De La Noval, B.; Pérez, E.; Martínez, B.; León, O.; Martínez-Gallardo, N.; Délano-Frier, J. Exogenous systemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrhiza 2007, 17, 449–460. [Google Scholar] [CrossRef]
- Amini, J.; Sidovich, D. The effects of fungicides on Fusarium oxysporum f. sp. lycopersici associated with fusarium wilt of tomato. J. Plant Prot. Res. 2010, 50, 172–178. [Google Scholar] [CrossRef]
- Gondal, A.S.; Riaz, K.; Khan, A.; Ijaz, M. Effect of different doses of fungicide (Mancozeb) against alternaria leaf blight of tomato in tunnel. J. Plant Pathol. Microbiol. 2012, 3, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.C.; Kumar, A.; Jamwal, S.; Jamval, R.; Jamwal, A. Integrated management of tomato wilt caused by Fusarium oxysporum f. sp. lycopersici. J. Entomol. Zool. Stud. 2018, 6, 1338–1341. [Google Scholar] [CrossRef]
- Marek, J.; De Azevedo, D.; Ono, E.O.; Rodrigues, J.D.; Faria, C.M.D.R. Photoynthetic and productive increase in tomato plants treated with strobilurins and carboxamides for the control of Alternaria solani. Sci. Hortic. 2018, 242, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Farooq, S.; Jat, R.R.; Gupta, A.; Singh, R.; Majeed, M.; Un Nabi, S.; Bashir, N.; Shah, O.S. Evaluation of different fungicides against Alternaria solani (Ellis & Martin) Sorauer cause of early blight of tomato under laboratory conditions. Pharma Innov. 2019, 8, 140–142. [Google Scholar]
- FRAC Code List© * 2020. Available online: https://www.frac.info/docs/default-source/publications/frac-code-list/frac-code-list-2020-finalb16c2b2c512362eb9a1eff00004acf5d.pdf?sfvrsn=54f499a_2 (accessed on 11 October 2020).
- Foolad, M.; Sharma, A.; Ashrafi, H.; Lin, G. Genetics of early blight resistance in tomato. Acta Hortic. 2005, 695, 397–406. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Panthee, D.R.; Gardner, R.G. Genetic improvement of fresh market tomatoes for yield and fruit quality over 35 years in North Carolina: A review. Int. J. Veg. Sci. 2011, 17, 259–273. [Google Scholar] [CrossRef]
- Catanzariti, A.-M.; Lim, G.T.T.; Jones, D.A. The tomato I-3 gene: A novel gene for resistance to Fusarium wilt disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, P.; Singh, P.; Sinha, B.; Singh, M.; Kumar, R.; Pandey, K.K.; Mathura, R. Sources of resistance against early blight (Alternaria solani) in tomato (Solanum lycopersicum). Indian J. Agr. Sci. 2009, 79, 752–753. [Google Scholar]
- Hassanein, N.; Zeid, M.; Youssef, K.; Mahmoud, D. Control of tomato early blight and wilt using aqueous extract of neem leaves. Phytopathol. Mediterr. 2010, 49, 143–151. Available online: https://www.jstor.org/stable/26458587 (accessed on 3 December 2020).
- Camlica, E.; Tozlu, E. Biological control of Alternaria solani in tomato. Fresenius Environ. Bull. 2019, 28, 7092–7100. [Google Scholar]
- Mulugeta, T.; Muhinyuza, J.-B.; Gouws-Meyer, R.; Matsaunyane, L.; Andreasson, E.; Alexandersson, E. Botanicals and plant strengtheners for potato and tomato cultivation in Africa. J. Integr. Agric. 2020, 19, 406–427. [Google Scholar] [CrossRef]
- Shcherbakova, L.; Odintsova, T.; Stakheev, A.A.; Fravel, D.R.; Zavriev, S.K. Identification of a novel small cysteine-rich protein in the fraction from the biocontrol Fusarium oxysporum strain CS-20 that mitigates fusarium wilt symptoms and triggers defense responses in tomato. Front. Plant Sci. 2016, 6, 1207. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Pandey, S.; Bhattacharya, A.; Mishra, A.; Nautiyal, C. Protective role of biosynthesized silver nanoparticles against early blight disease in Solanum lycopersicum. Plant Physiol. Biochem. 2017, 121, 216–225. [Google Scholar] [CrossRef]
- Campbell, B.; Chan, K.L.; Kim, J.H. Chemosensitization as a means to augment commercial antifungal agents. Front. Microbiol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzhavakhiya, V.; Shcherbakova, L.; Semina, Y.; Zhemchuzhina, N.; Campbell, B. Chemosensitization of plant pathogenic fungi to agricultural fungicides. Front. Microbiol. 2012, 3, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, R.P. A reassessment of the risk of rust fungi developing resistance to fungicides. Pest Manag. Sci. 2014, 70, 1641–1645. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The evolution of fungicide resistance. Adv. Virus Res. 2015, 90, 29–92. [Google Scholar] [CrossRef]
- Kim, J.H.; Chan, K.L. Augmenting the antifungal activity of an oxidizing agent with kojic acid: Control of Penicillium strains infecting crops. Molecules 2014, 19, 18448–18464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Lee, Y.; Ha, A.; Kim, J.-C.; Park, A.R.; Yu, N.H.; Son, H.; Choi, G.J.; Park, H.W.; Lee, C.W.; et al. Chemosensitization of Fusarium graminearum to chemical fungicides using cyclic lipopeptides produced by Bacillus amyloliquefaciens strain JCK-12. Front. Plant Sci. 2017, 8, 2010. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, L.A.; Syomina, Y.V.; Arslanova, L.R.; Nazarova, T.A.; Dzhavakhiya, V. Metabolites secreted by a nonpathogenic Fusarium sambucinum inhabiting wheat rhizosphere enhance fungicidal effect of some triazoles against Parastagonospora nodorum. AIP Conf. Proc. 2019, 2063, 030018. [Google Scholar] [CrossRef]
- Kartashov, M.I.; Shcherbakova, L.A.; Dzhavakhiya, V.G. In vitro enhancement of the sensitivity to tebuconazole in Bipolaris sorokiniana, a causative agent of cereal root rots, by a microbial metabolite 6-demethylmevinolin. In Proceedings of the 19th International Reinhardsbrunn Symposium, Braunschweig, Friedrichoda, Germany, 7–11 April 2019; p. 48. [Google Scholar]
- Alan, A.R.; Earle, E.D. Sensitivity of bacterial and fungal plant pathogens to the lytic peptides, MSI-99, Magainin II, and Cecropin B. Mol. Plant-Microbe Interact. 2002, 15, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.P.; Wang, G.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Das, K.; Datta, K.; Karmakar, S.; Datta, S.K. Antimicrobial peptides–Small but mighty weapons for plants to fight phytopathogens. Protein Pept. Lett. 2019, 26, 720–742. [Google Scholar] [CrossRef]
- Beintema, J.J. Structural features of plant chitinases and chitin-binding proteins. FEBS Lett. 1994, 350, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, S.; Iida, Y.; Kawasaki, Y.; Minami, Y.; Watanabe, K.; Yagi, F. The chitin-binding capability of Cy-AMP1 from cycad is essential to antifungal activity. J. Pept. Sci. 2009, 15, 492–497. [Google Scholar] [CrossRef]
- Chen, C.-S.; Chen, C.-Y.; Ravinath, D.M.; Bungahot, A.; Cheng, C.-P.; You, R.-I. Functional characterization of chitin-binding lectin from Solanum integrifolium containing anti-fungal and insecticidal activities. BMC Plant Biol. 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yen, T.-B.; Chang, S.-T. Synergistic effects of cinnamaldehyde in combination with eugenol against wood decay fungi. Bioresour. Technol. 2008, 99, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Canhoto, J.; Vale-Silva, L.; Silva, M.J.; Pinto, E.; Salgueiro, L. Chemical composition and antifungal activity of the essential oils of Lavandula viridis L’Hér. J. Med Microbiol. 2011, 60, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Campbell, B.; Mahoney, N.; Chan, K.; Molyneux, R.; May, G. Chemosensitization prevents tolerance of Aspergillus fumigatus to antimycotic drugs. Biochem. Biophys. Res. Commun. 2008, 372, 266–271. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Vassilevski, A.A.; Slavokhotova, A.A.; Musolyamov, A.K.; Finkina, E.I.; Khadeeva, N.V.; Rogozhin, E.A.; Korostyleva, T.V.; Pukhalsky, V.A.; Grishin, E.V.; et al. A novel antifungal hevein-type peptide from Triticum kiharae seeds with a unique 10-cysteine motif. FEBS J. 2009, 276, 4266–4275. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Rogozhin, E.A.; Utkina, L.; Vassilevski, A.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. Genes encoding hevein-like defense peptides in wheat: Distribution, evolution, and role in stress response. Biochimie 2012, 94, 1009–1016. [Google Scholar] [CrossRef]
- Istomina, E.A.; Slavokhotova, A.A.; Korostyleva, T.V.; Semina, Y.; Shcherbakova, L.A.; Pukhalskij, V.A.; Odintsova, T.I. Genes encoding hevein-like antimicrobial peptides WAMPs in the species of the genus Aegilops L. Russ. J. Genet. 2017, 53, 1320–1327. [Google Scholar] [CrossRef]
- Odintsova, T.; Shcherbakova, L.; Slezina, M.; Pasechnik, T.; Kartabaeva, B.; Istomina, E.; Dzhavakhiya, V. Hevein-like antimicrobial peptides WAMPs: Structure-function relationship in antifungal activity and sensitization of plant pathogenic fungi to tebuconazole by WAMP-2-derived peptides. Int. J. Mol. Sci. 2020, 21, 7912. [Google Scholar] [CrossRef]
- Efremov, R.G.; Vassilevski, A.A.; Slavokhotova, A.A.; Odintsova, T.I.; Grishin, E.V.; Egorov, T.A.; Arseniev, A.S. Solution structure of a defense peptide from wheat with a 10-cysteine motif. Biochem. Biophys. Res. Commun. 2011, 411, 14–18. [Google Scholar] [CrossRef]
- Slezina, M.P.; Korostyleva, T.V.; Slavokhotova, A.A.; Istomina, E.A.; Shcherbakova, L.A.; Pukhalskij, V.A.; Odintsova, T. Genes encoding hevein-like antimicrobial peptides from Elytrigia repens (L.) Desv. ex Nevski. Russ. J. Genet. 2018, 54, 1152–1159. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Naumann, T.A.; Price, N.P.J.; Rogozhin, E.A.; Andreev, Y.A.; Vassilevski, A.A.; Odintsova, T.I. Novel mode of action of plant defense peptides-hevein-like antimicrobial peptides from wheat inhibit fungal metalloproteases. FEBS J. 2014, 281, 4754–4764. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Cools, T.L.; Harvey, P.J.; Craik, D.J.; Braem, A.; Vleugels, J.; De Coninck, B.; Cammue, B.; Thevissen, K. The radish defensins RsAFP1 and RsAFP2 act synergistically with caspofungin against Candida albicans biofilms. Pept. 2016, 75, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Dzhavakhiya, V.G.; Voinova, T.M.; Shumilina, D.V. Search for the active center of peptidyl-prolyl cys/trans isomerase from Pseudomonas fluorescens responsible for the induction of tobacco (Nicotiana tabacum L.) plant resistance to tobacco mosaic virus. Agr. Bio. (Sel’skokhozyaistvennaya Biologiya) 2016, 51, 392–400. [Google Scholar] [CrossRef]
- Ma, Z.; Michailides, T.J. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop. Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Fourtouni, A.; Manetas, Y.; Christias, C. Effects of UV-B radiation on growth, pigmentation, and spore production in the phytopathogenic fungus Alternaria solani. Can. J. Bot. 1998, 76, 2093–2099. [Google Scholar] [CrossRef]
- Utkina, L.L.; Zhabon, E.O.; Slavokhotova, A.A.; Rogozhin, E.A.; Shiyan, A.N.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I.; Pukhal’Skiy, V.A. Heterologous expression of a synthetic gene encoding a novel hevein-type antimicrobial peptide of Leymus arenarius in Escherichia coli cells. Russ. J. Genet. 2010, 46, 1449–1454. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: New York, NY, USA, 2005; pp. 571–607. [Google Scholar]
- Aver’yanov, A.A.; Lapikova, V.P.; Pasechnik, T.D.; Abramova, O.S.; Gaivoronskaya, L.M.; Kuznetsov, V.V.; Baker, C.J. Pre-illumination of rice blast conidia induces tolerance to subsequent oxidative stress. Fungal Biol. 2014, 118, 743–753. [Google Scholar] [CrossRef]
- Orhan, G.; Bayram, A.; Zer, Y.; Balci, I. Synergy tests by E test and checkerboard methods of antimicrobial combinations against Brucella melitensis. J. Clin. Microbiol. 2005, 43, 140–143. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, L.A.; Nazarova, T.A.; Mikityuk, O.D.; Fravel, D.R. Fusarium sambucinum isolate FS-94 induces resistance against fusarium wilt of tomato via activation and priming of a salicylic acid-dependent signaling system. Russ. J. Plant. Physiol. 2011, 58, 808–818. [Google Scholar] [CrossRef]
- Sparling, D.W. Modeling in Ecotoxicology. In Ecotoxicology Essentials; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 361–390. [Google Scholar]
- Richer, D.L. Synergism–A patent view. Pestic. Sci. 1987, 19, 309–315. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides, mg/mL | F. oxysporum | A. solani | ||||
|---|---|---|---|---|---|---|
| ED50 | ED99 (MFC) * | R2 | ED50 | ED99 (MFC) * | R2 | |
| WAMP-N | 0.24 a | 22.7 | 0.997 | n/d | n/d | n/d |
| WAMP-G1 | >1000 | >10,000 | 0.936 | 40.0 c | 3980.0 f | 0.872 |
| WAMP-G2 | 0.35 a | 35.1 b | 0.961 | n/d | n/d | n/d |
| WAMP-C | 35.2 b | 106.9 d | 0.975 | 0.55 e | 53.4 g | 0.998 |
| WAMP-2 | 45.0 c | 79.0 d | 0.885 | n/d | n/d | n/d |
| Peptide Fractional Concentrations, µg/mL | Fractional Concentrations of Folicur®, µg/mL | ||||||
|---|---|---|---|---|---|---|---|
| 0.0 | 0.005 | 0.01 | 0.05 | ||||
| Inhibitory Effect, % | |||||||
| Er | Er | Ee | Er | Ee | Er | Ee | |
| WAMP-N | |||||||
| 0 | − | 6.7 aa | − | 16.9 ab | − | 26.4 ac | − |
| 50 | 12.3 a | 30.2 b,A | 18.2 | 45.9 b,A | 27.1 | 58.4 b | 35.5 |
| 100 | 23.1 b | 48.4 c,B | 28.3 | 58.3 c | 36.1 | 70.8 c,D | 43.4 |
| 200 | 42.7 c | 64.1 d,C | 48.9 | 69.7 d,C | 52.4 | 75.0 c,D | 57.8 |
| WAMP-C | |||||||
| 0 | − | 6.5 aa | − | 17.8 ab | − | 25.9 ac | − |
| 50 | 4.3 a | 33.1 b,A | 10.5 | 44.2 b,A | 21.0 | 51.9 b | 29.1 |
| 100 | 6.2 b | 46.2 c,B | 12.3 | 66.1 c,C | 23.8 | 76.8 c,D | 30.5 |
| 200 | 8.6 c | 67.3 d,C | 14.5 | 83.7 d | 24.9 | 91.7 d | 32.3 |
| 400 | 11.6 c | 77.8 e | 17.3 | 87.8 d | 27.3 | 95.9 e | 34.5 |
| WAMP-G1 | |||||||
| 0 | − | 9.0 aa | − | 18.4 ab | − | 29.2 ac | − |
| 100 | 1.1 a | 15.2 b | 10.1 | 30.2 b | 19.3 | 45.3 b | 30.0 |
| 200 | 8.0 b | 28.4 c | 16.3 | 39.0 c | 24.9 | 49.8 b | 34.8 |
| 400 | 17.5 c | 45.4 d | 24.9 | 56.3 d | 32.7 | 64.9 c | 41.7 |
| WAMP-G2 | |||||||
| 0 | − | 8.1 aa | − | 19.7 ab | − | 28.0 ac | − |
| 50 | 7.0 a | 27.8 b | 14.5 | 36.4 b | 24.8 | 47.4 b | 33.0 |
| 100 | 22.4 b | 49.3 c | 28.8 | 53.1 c | 37.7 | 56.8 c | 44.1 |
| 200 | 34.1 c | 62.6 d | 41.8 | 74.8 d | 47.1 | 87.1 d | 52.7 |
| Peptide Fractional Concentrations, µg/mL | Fractional Concentrations of Folicur®, µg/mL * | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.0 | 0.01 | 0.05 | 0.1 | 0.2 | |||||
| Inhibitory Effect, % | |||||||||
| Er | Er | Ee | Er | Ee | Er | Ee | Er | Ee | |
| WAMP-G1 | |||||||||
| 0 | − | 6.9 aa | 13.1 ab | 28.2 ac | 40.9 ad | − | |||
| 100 | 2.0 a | 15.1 b,A | 8.8 A | 45.4 b | 14.8 | 64.7 b | 29.8 | 75.0 b | 42.1 |
| 200 | 9.8 b | 44.9 c | 16.7 | 63.1 c | 21.6 | 74.3 c | 35.2 | 78.4 b | 46.7 |
| 400 | 21.9 c | 54.2 c | 27.3 | 67.4 c | 32.1 | 78.4 c | 43.9 | 81.0 b | 53.8 |
| WAMP-C | |||||||||
| 0 | − | 6.7 aa | − | 12.9 ab | − | 28.1 ac | − | 40.5 ad | − |
| 100 | 19.0 a | 26.6 b,B | 24.5 B | 32.4 b,C | 29.4 C | 51.4 b,D | 41.8 D | 64.3 b | 51.8 |
| 200 | 31.0 b | 51.3 c | 35.6 | 54.9 c | 39.9 | 59.1 b,E | 50.5 E | 69.4 b,F | 59.7 F |
| 400 | 48.2 c | 70.8 d | 51.7 | 87.0 d | 54.9 | 92.3 c | 62.8 | 93.8 c | 69.3 |
| Peptide Name | Amino Acid Sequence | Length, aa Residues | Molecular Weight, Da * |
|---|---|---|---|
| WAMP-N | AQRCGDQARGAKC | 13 (1–13) ** | 1363.53 |
| WAMP-G1 | LCCGKYGFCGSG | 12 (17–28) ** | 1194.40 |
| WAMP-G2 | CCGKYGFCGSGDAYC | 15 (18–32) ** | 1533.73 |
| WAMP-C | GKGSCQSQCRGCR | 13 (33–45) ** | 1369.55 |
| WAMP-2 | AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGKGSCQSQCRGCR | 45 | 4658.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shcherbakova, L.; Odintsova, T.; Pasechnik, T.; Arslanova, L.; Smetanina, T.; Kartashov, M.; Slezina, M.; Dzhavakhiya, V. Fragments of a Wheat Hevein-Like Antimicrobial Peptide Augment the Inhibitory Effect of a Triazole Fungicide on Spore Germination of Fusarium oxysporum and Alternaria solani. Antibiotics 2020, 9, 870. https://doi.org/10.3390/antibiotics9120870
Shcherbakova L, Odintsova T, Pasechnik T, Arslanova L, Smetanina T, Kartashov M, Slezina M, Dzhavakhiya V. Fragments of a Wheat Hevein-Like Antimicrobial Peptide Augment the Inhibitory Effect of a Triazole Fungicide on Spore Germination of Fusarium oxysporum and Alternaria solani. Antibiotics. 2020; 9(12):870. https://doi.org/10.3390/antibiotics9120870
Chicago/Turabian StyleShcherbakova, Larisa, Tatyana Odintsova, Tatyana Pasechnik, Lenara Arslanova, Tatyana Smetanina, Maxim Kartashov, Marina Slezina, and Vitaly Dzhavakhiya. 2020. "Fragments of a Wheat Hevein-Like Antimicrobial Peptide Augment the Inhibitory Effect of a Triazole Fungicide on Spore Germination of Fusarium oxysporum and Alternaria solani" Antibiotics 9, no. 12: 870. https://doi.org/10.3390/antibiotics9120870
APA StyleShcherbakova, L., Odintsova, T., Pasechnik, T., Arslanova, L., Smetanina, T., Kartashov, M., Slezina, M., & Dzhavakhiya, V. (2020). Fragments of a Wheat Hevein-Like Antimicrobial Peptide Augment the Inhibitory Effect of a Triazole Fungicide on Spore Germination of Fusarium oxysporum and Alternaria solani. Antibiotics, 9(12), 870. https://doi.org/10.3390/antibiotics9120870