Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification of Marine Bacteria BO53 and GA327
2.2. Antibacterial Activity of VOCs Produced by Bacillus sp. BO53 and Pseudoalteromonas sp. GA327
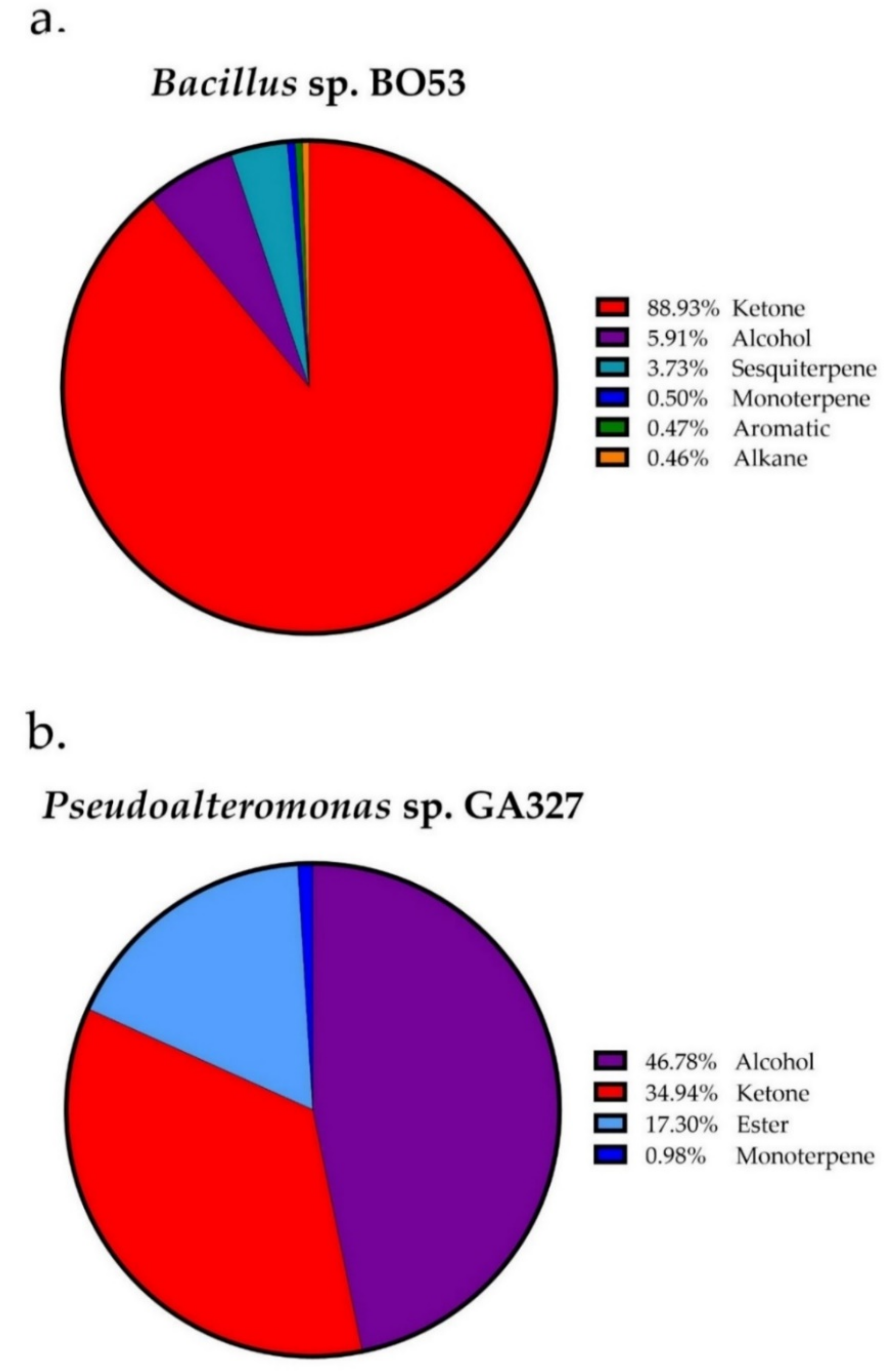
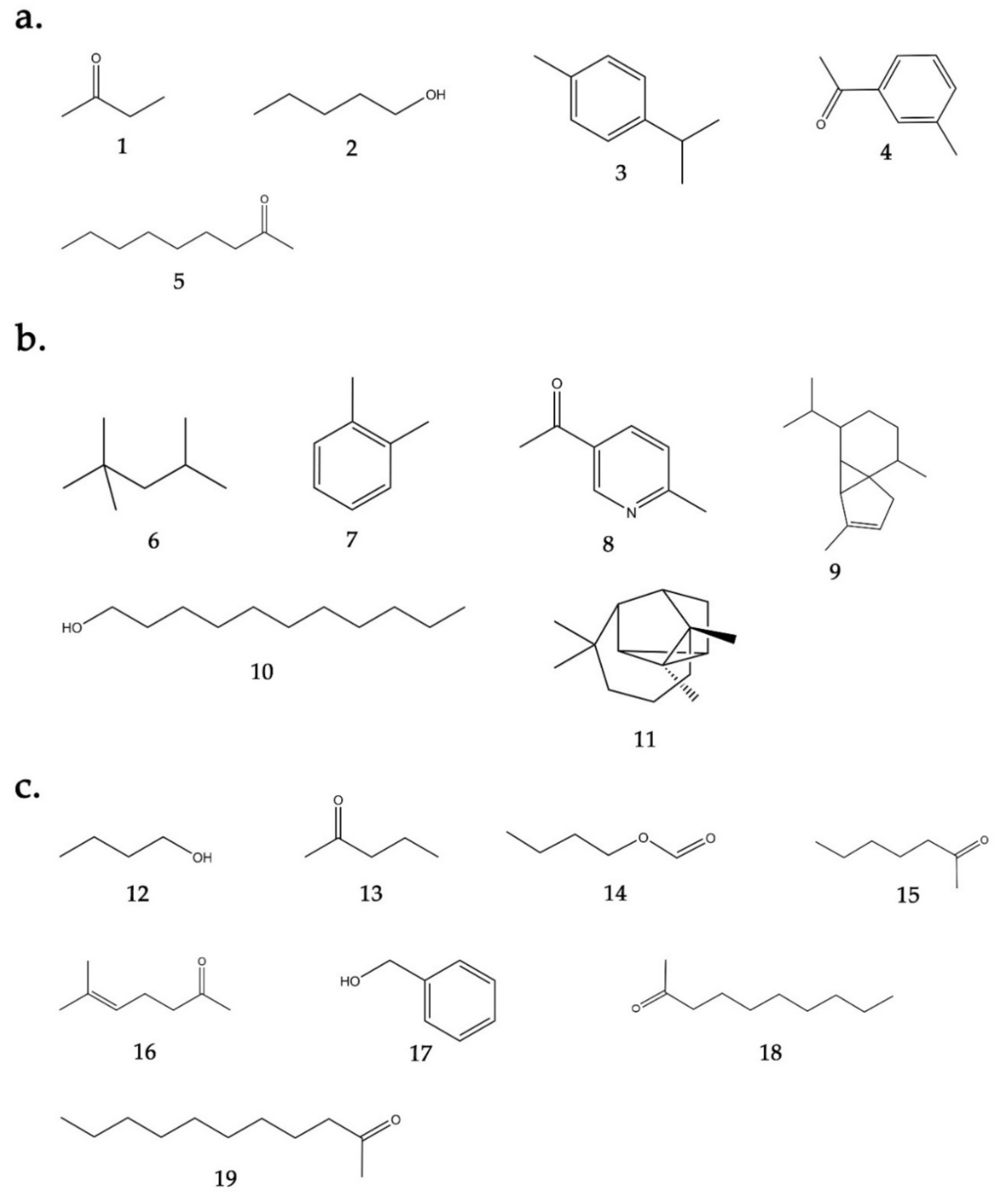
2.3. Identification of VOCs from Bacillus sp. BO53 and Pseudoalteromonas sp. GA327
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolation from Octocorals
4.2. Molecular Identification of Octocoral-Associated Bacteria Species
4.3. Pathogenic Bacterial Strains
4.4. Antibacterial Activity of Marine bVOCs
4.5. Marine Bacteria Volatolome Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Patini, R.; Mangino, G.; Martellacci, L.; Quaranta, G.; Masucci, L.; Gallenzi, P. The Effect of Different Antibiotic Regimens on Bacterial Resistance: A Systematic Review. Antibiotics 2020, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Priority List of Antibiotic-Resistance Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 16 May 2020).
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus virulence. Microbiol. Spectr. 2018, 6, 669–686. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39–68. [Google Scholar] [CrossRef] [Green Version]
- Asif, M.; Alvi, I.A. Insight into Acinetobacter baumannii: Pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infect. Drug Resist. 2018, 11, 1249–1260. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.B. Confronting the challenges of discovery of novel antibacterial agents. Bioorg. Med. Chem. Lett. 2014, 24, 3683–3689. [Google Scholar] [CrossRef] [Green Version]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Sneed, J.M.; Ritson-Williams, R.; Young, R. Marine chemical ecology in benthic environments. Nat. Prod. Rep. 2019, 36, 410–429. [Google Scholar] [CrossRef]
- Sang, V.T.; Dat, T.T.H.; Vinh, L.B.; Cuong, L.C.V.; Oanh, P.T.T.; Ha, H.; Kim, Y.H.; Anh, H.L.T.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific source of Potential Bioactive Natural Products. Mar. Drugs. 2019, 17, 468. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the Core Microbiome in Corals’ Microbial Soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Atencio, L.A.; Dal Grande, F.; Young, G.O.; Gavilán, R.; Guzmán, H.M.; Schmitt, I.; Mejía, L.C.; Gutiérrez, M. Antimicrobial-producing Pseudoalteromonas from the marine environment of Panama shows a high phylogenetic diversity and clonal structure. J. Basic Microbiol. 2018, 58, 747–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive secondary metabolites from octocoral-Associated microbes—New chances for blue growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, S.; Dickschat, J.S.; Kunze, B.; Wagner-Dobler, I.; Diestel, R.; Sasse, F. Biological activity of volatiles from marine and terrestrial bacteria. Mar. Drugs 2010, 8, 2976–2987. [Google Scholar] [CrossRef] [PubMed]
- Papaleo, M.C.; Fondi, M.; Maida, I.; Perrin, E.; Lo Giudice, A.; Michaud, L.; Mangano, S.; Bartolucci, G.; Romoli, R.; Fani, R. Sponge-associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012, 30, 272–293. [Google Scholar] [CrossRef]
- Papaleo, M.C.; Romoli, R.; Bartolucci, G.; Maida, I.; Perrin, E.; Fondi, M.; Orlandini, V.; Mengoni, A.; Emiliani, G.; Tutino, M.L.; et al. Bioactive volatile organic compounds from Antarctic (sponges) bacteria. New Biotechnol. 2013, 30, 824–838. [Google Scholar] [CrossRef]
- Romoli, R.; Papaleo, M.C.; De Pascale, D.; Tutino, M.L.; Michaud, L.; LoGiudice, A.; Fani, R.; Bartolucci, G. GC-MS volatolomic approach to study the antimicrobial activity of the antarctic bacterium Pseudoalteromonas sp. TB41. Metabolomics 2014, 10, 42–51. [Google Scholar] [CrossRef]
- Orlandini, V.; Maida, I.; Fondi, M.; Perrin, E.; Papaleo, M.C.; Bosi, E.; de Pascale, D.; Tutino, M.L.; Michaud, L.; Lo Giudice, A.; et al. Genomic analysis of three sponge-associated Arthrobacter Antarctic speciess, inhibiting the growth of Burkholderia cepacia complex bacteria by synthesizing volatile organic compounds. Microbiol. Res. 2014, 169, 593–601. [Google Scholar] [CrossRef]
- Sannino, F.; Parrilli, E.; Apuzzo, G.A.; de Pascale, D.; Tedesco, P.; Maida, I.; Perrin, E.; Fondi, M.; Fani, R.; Marino, G.; et al. Pseudoalteromonas haloplanktis produces methylamine, a volatile compound active against Burkholderia cepacia complex strains. New Biotechnol. 2017, 35, 13–18. [Google Scholar] [CrossRef]
- Risticevic, S.; Lord, H.; Górecki, T.; Arthur, C.L.; Pawliszyn, J. Protocol for solid-phase microextraction method development. Nat. Protoc. 2010, 5, 122–139. [Google Scholar] [CrossRef]
- Durant, A.A.; Rodríguez, C.; Herrera, L.; Almanza, A.; Santana, A.I.; Spadafora, C.; Gupta, M.P. Anti-malarial activity and HS-SPME-GC-MS chemical profiling of Plinia cerrocampanensis leaf essential oil. Malar. J. 2014, 13, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laukaleja, I.; Kruma, Z. Evaluation of a headspace solid-phase microextraction with different fibres for volatile compoun d determination in specialtycoffee brews. Res. Rural Dev. 2019, 1, 215–221. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; De Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef] [PubMed]
- Altaf, A.A.; Shahzad, A.; Gul, Z.; Rasool, N.; Badshah, A.; Lal, B.; Khan, E. A Review on the Medicinal Importance of Pyridine Derivatives. J. Drug Des. Med. Chem. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Strobel, G.A.; Moore, E.; Robison, R.; Sears, J. Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 2010, 156, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Arámbula, C.I.; Diaz, C.E.; Garcia, M.I. Performance, chemical composition and antibacterial activity of the essential oil of Ruta chalepensis and Origanum vulgare. J. Phys. Conf. Ser. 2019, 1386. [Google Scholar] [CrossRef]
- Togashi, N.; Shiraishi, A.; Nishizaka, M.; Matsuoka, K.; Endo, K.; Hamashima, H.; Inoue, Y. Antibacterial activity of long-chain fatty alcohols against Staphylococcus aureus. Molecules 2007, 12, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Yano, T.; Miyahara, Y.; Morii, N.; Okano, T.; Kubota, H. Pentanol and benzyl alcohol attack bacterial surface structures differently. Appl. Environ. Microbiol. 2016, 82, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Hanbali, E.L.F.; Mellouki, F.; Akssira, M.; Boira, H.; Blázquez, M.A. Composition and antimicrobial activity of essential oil of anthemis tenuisecta ball. J. Essent. Oil Bear. Plants 2007, 10, 499–503. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Korsten, L. Effect of volatile compounds produced by Bacillus strains on postharvest decay in citrus. Biol. Control 2010, 53, 122–128. [Google Scholar] [CrossRef]
- Popova, A.A.; Koksharova, O.A.; Lipasova, V.A.; Zaitseva, J.V.; Katkova-Zhukotskaya, O.A.; Eremina, S.I.; Mironov, A.S.; Chernin, L.S.; Khmel, I.A. Inhibitory and toxic effects of volatiles emitted by strains of Pseudomonas and Serratia on growth and survival of selected microorganisms, Caenorhabditis elegans, and Drosophila melanogaster. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ould Bellahcen, T.; Cherki, M.; Sánchez, J.A.C.; Cherif, A.; EL Amrani, A. Chemical Composition and Antibacterial Activity of the Essential Oil of Spirulina platensis from Morocco. J. Essent. Oil Bear. Plants 2019, 22, 1265–1276. [Google Scholar] [CrossRef]
- Benali, T.; Habbadi, K.; Khabbach, A.; Marmouzi, I. GC–MS Analysis, Antioxidant and Antimicrobial Activities of Achillea Odorata Subsp. Pectinata and Ruta Montana Essential Oils and Their Potential Use as Food Preservatives. Foods 2020, 9, 668. [Google Scholar] [CrossRef]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Venturini, M.E. Antifungal activity of the volatile organic compounds produced by Bacillus velezensis strains against postharvest fungal pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Romoli, R.; Papaleo, M.C.; De Pascale, D.; Tutino, M.L.; Michaud, L.; Logiudice, A.; Fani, R.; Bartolucci, G. Characterization of the volatile profile of Antarctic bacteria by using solid-phase microextraction-gas chromatography-mass spectrometry. J. Mass Spectrom. 2011, 46, 1051–1059. [Google Scholar] [CrossRef]
- Correa, R.; Coronado, L.M.; Garrido, A.C.; Durant-Archibold, A.A.; Spadafora, C. Volatile organic compounds associated with Plasmodium falciparum infection in vitro. Parasites Vectors 2017, 10, 215. [Google Scholar] [CrossRef] [Green Version]
- Garrido, A.; Ledezma, J.G.; Durant-Archibold, A.A.; Allen, N.S.; Villarreal A, J.C.; Gupta, M.P. Chemical Profiling of Volatile Components of the Gametophyte and Sporophyte Stages of the Hornwort Leiosporoceros dussii (Leiosporocerotaceae) From Panama by HS-SPME-GC-MS. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | Strain | Bacillus sp. BO53 | Pseudoalteromonas sp. GA327 | ||
|---|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | ||
| A. baumanni | ATCC 19606 | - | ± | + | + |
| S. aureus | ATCC 43300 | - | ± | - | - |
| P. aeruginosa | ATCC 10145 | - | + | ± | + |
| % Detected Compound on Each Species | ||||
|---|---|---|---|---|
| Compound | TRI | ERI | Bacillus sp. BO53 | Pseudoalteromonas sp. GA327 |
| 2-Butanone | 602 | 609 | 17.03 | 20.14 |
| 1-Butanol | 671 | 676 | - | 4.55 |
| 2,2,4-Trimethylpentane | 680 | 687 | 0.46 | - |
| 2-Pentanone | 687 | 693 | - | 1.29 |
| 1-Pentanol | 775 | 780 | 1.33 | 38.91 |
| Butyl formate | 787 | 793 | - | 17.30 |
| o-Xylene | 884 | 891 | 0.47 | - |
| 2-Heptanone | 889 | 895 | - | 7.99 |
| 6-Methyl-5-heptene-2-one | 988 | 995 | - | 0.37 |
| p-Cymene | 1021 | 1027 | 0.50 | 0.98 |
| Benzyl Alcohol | 1033 | 1040 | - | 3.32 |
| 2-Nonanone | 1096 | 1102 | 7.00 | 1.38 |
| 3-Methylacetophenone | 1176 | 1184 | 0.26 | 1.18 |
| 5-Acetyl-2-methylpyridine | 1189 | 1193 | 64.63 | - |
| 2-Decanone | 1190 | 1198 | - | 1.01 |
| 2-Undecanone | 1291 | 1300 | - | 1.58 |
| α-Cubebene | 1354 | 1355 | 0.43 | - |
| 1-Undecanol | 1370 | 1374 | 4.58 | - |
| α-Longicyclene | 1374 | 1380 | 3.30 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, A.; Atencio, L.A.; Bethancourt, R.; Bethancourt, A.; Guzmán, H.; Gutiérrez, M.; Durant-Archibold, A.A. Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327. Antibiotics 2020, 9, 923. https://doi.org/10.3390/antibiotics9120923
Garrido A, Atencio LA, Bethancourt R, Bethancourt A, Guzmán H, Gutiérrez M, Durant-Archibold AA. Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327. Antibiotics. 2020; 9(12):923. https://doi.org/10.3390/antibiotics9120923
Chicago/Turabian StyleGarrido, Anette, Librada A. Atencio, Rita Bethancourt, Ariadna Bethancourt, Héctor Guzmán, Marcelino Gutiérrez, and Armando A. Durant-Archibold. 2020. "Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327" Antibiotics 9, no. 12: 923. https://doi.org/10.3390/antibiotics9120923
APA StyleGarrido, A., Atencio, L. A., Bethancourt, R., Bethancourt, A., Guzmán, H., Gutiérrez, M., & Durant-Archibold, A. A. (2020). Antibacterial Activity of Volatile Organic Compounds Produced by the Octocoral-Associated Bacteria Bacillus sp. BO53 and Pseudoalteromonas sp. GA327. Antibiotics, 9(12), 923. https://doi.org/10.3390/antibiotics9120923