A Review of Fungal Protoilludane Sesquiterpenoid Natural Products



{kind=link}
{kind=link}
{kind=link}
{kind=link}
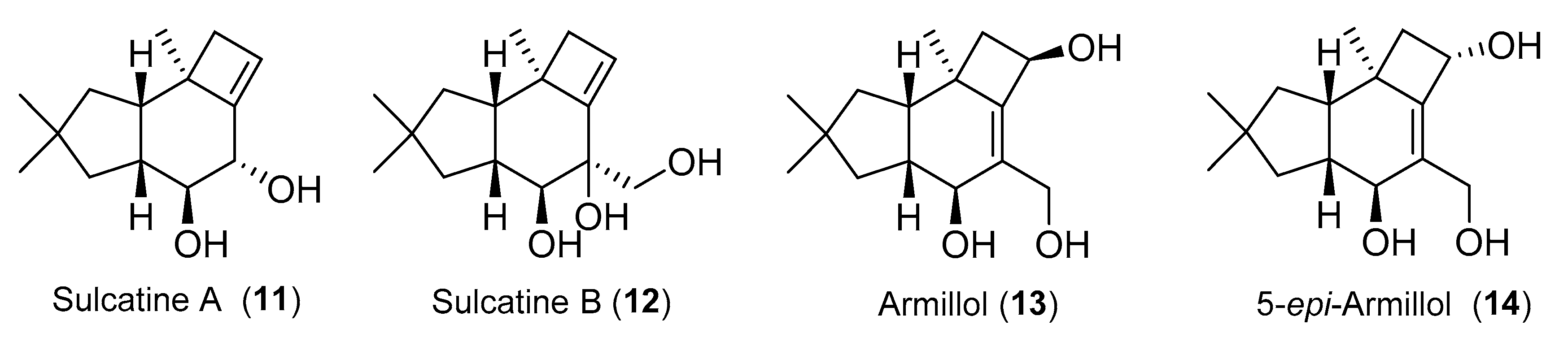{kind=link}
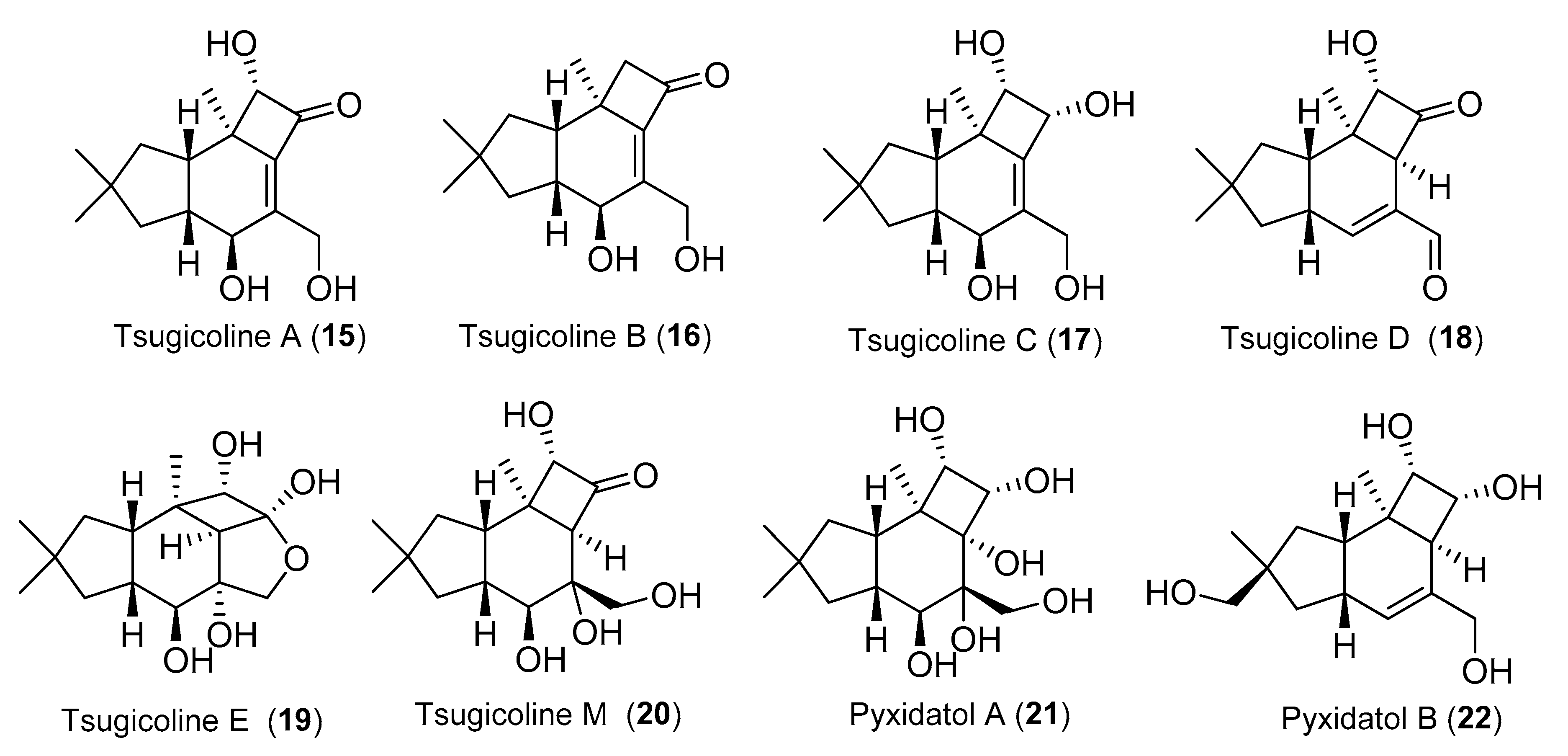{kind=link}
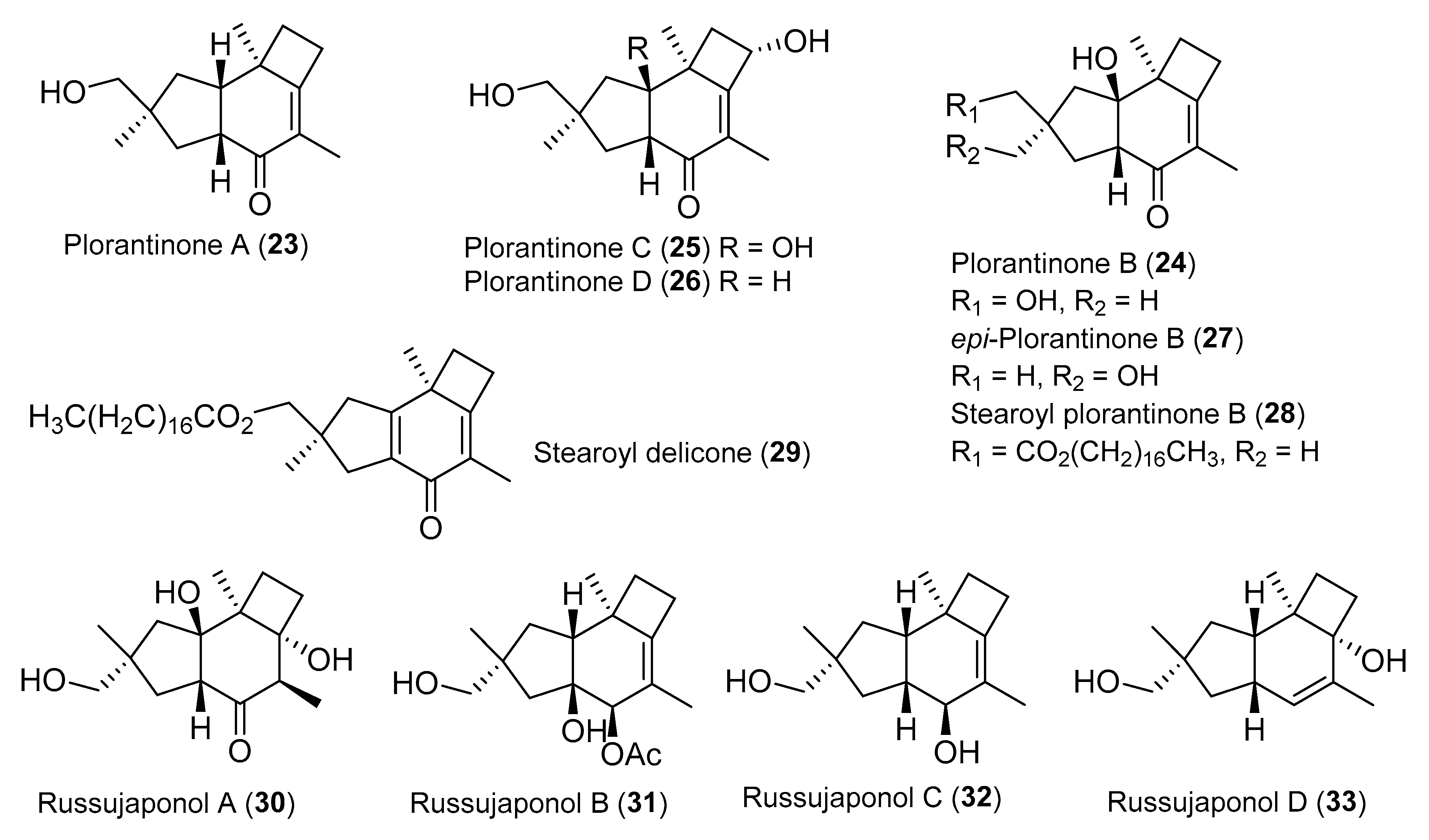{kind=link}
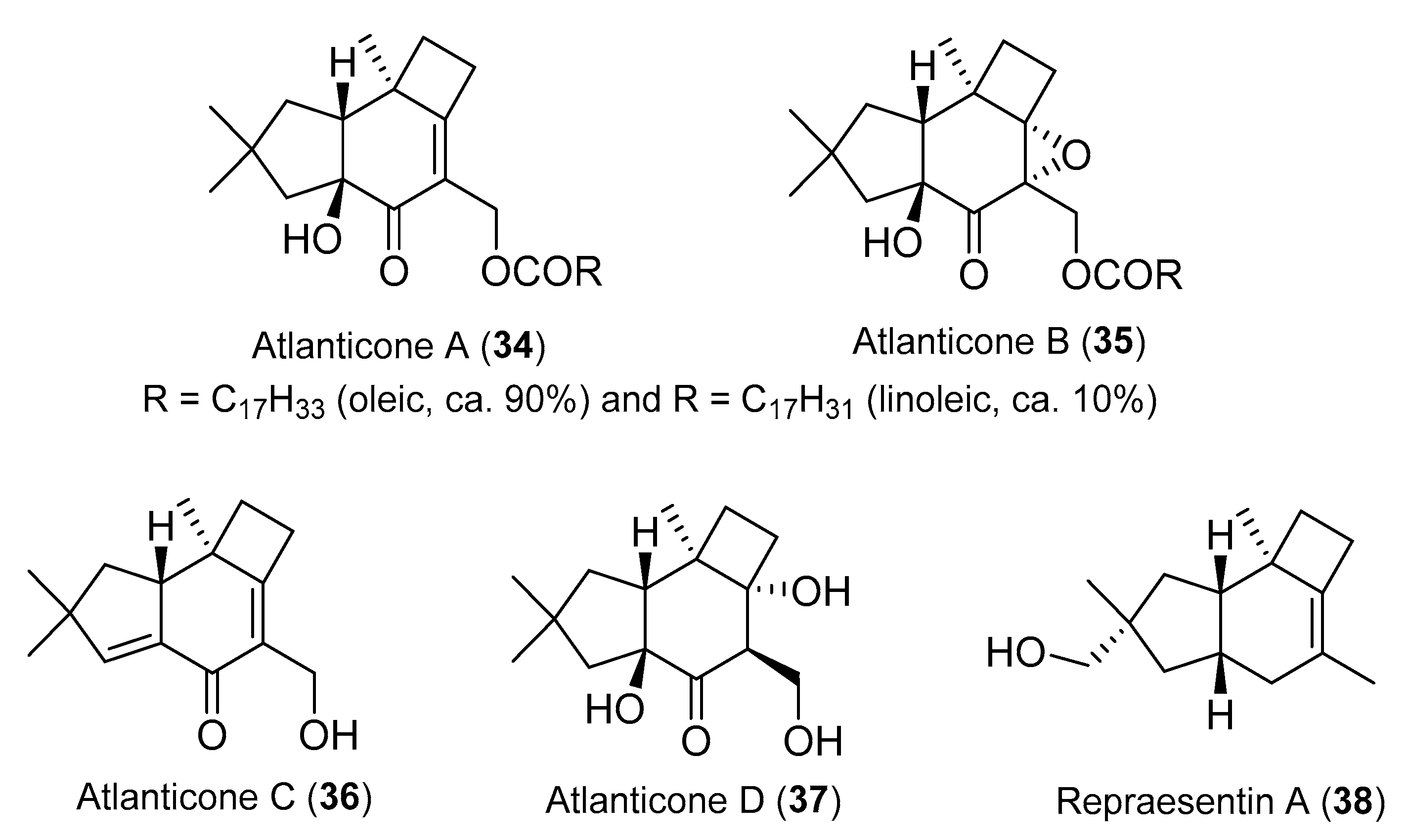{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
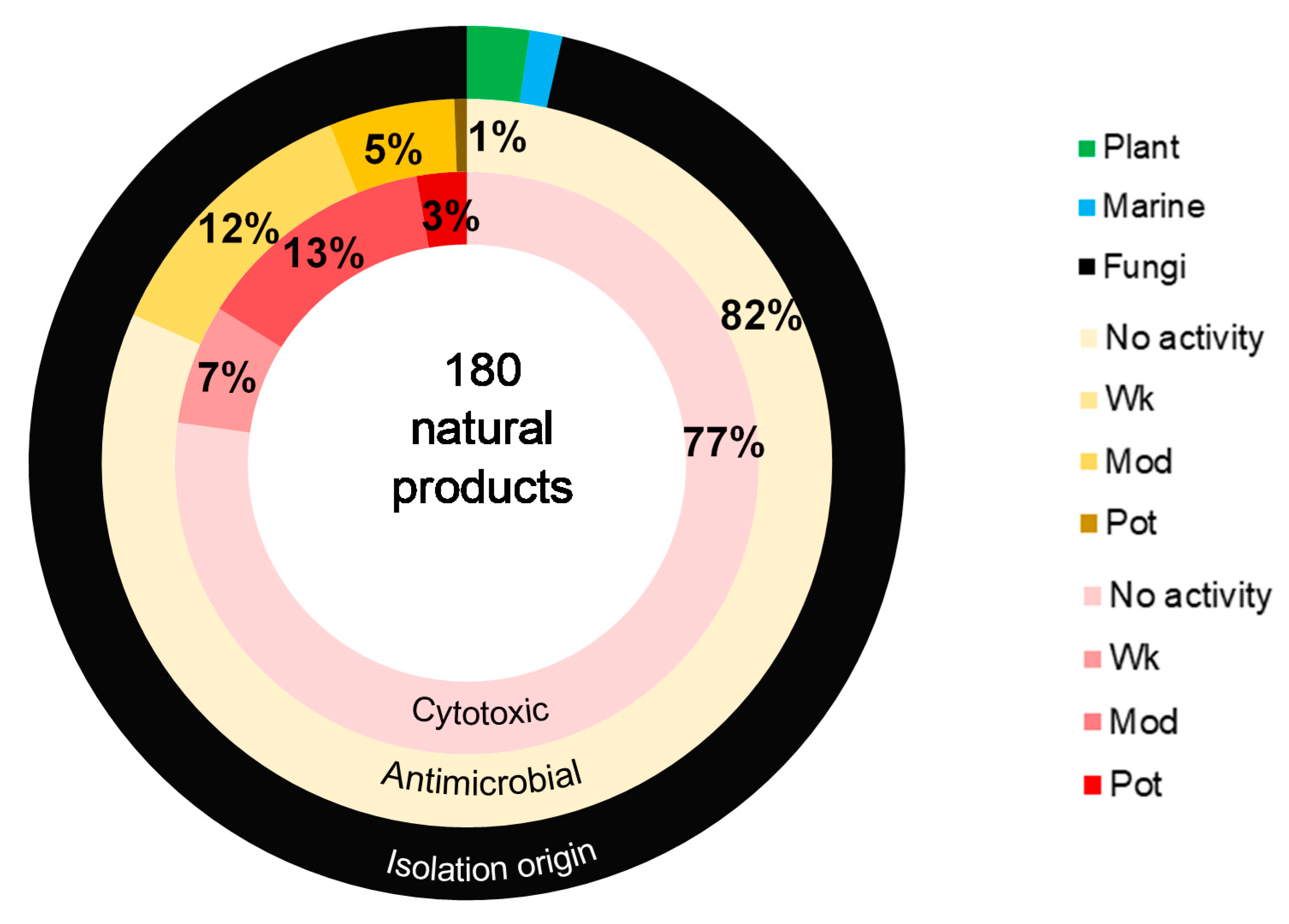
Abstract
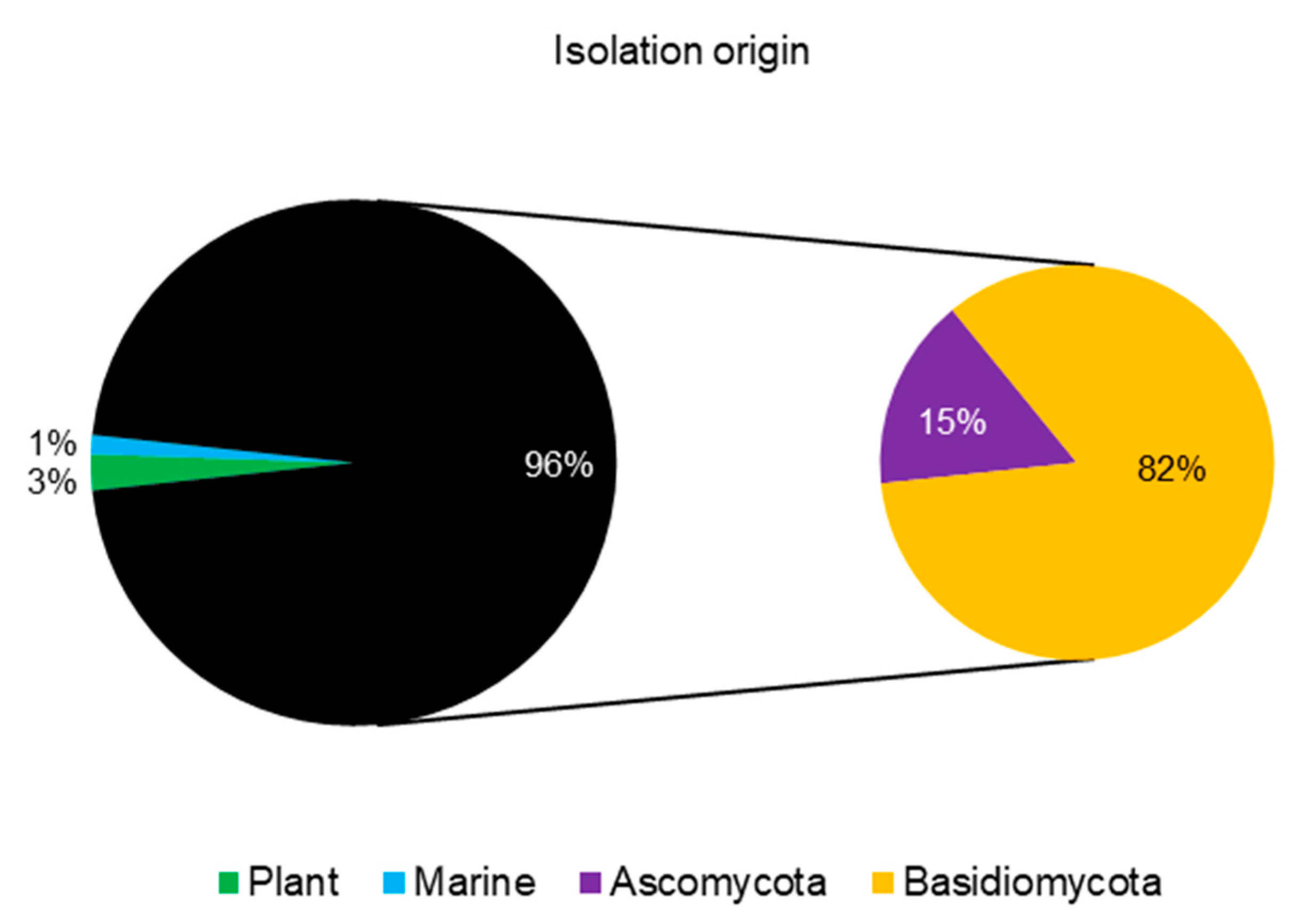
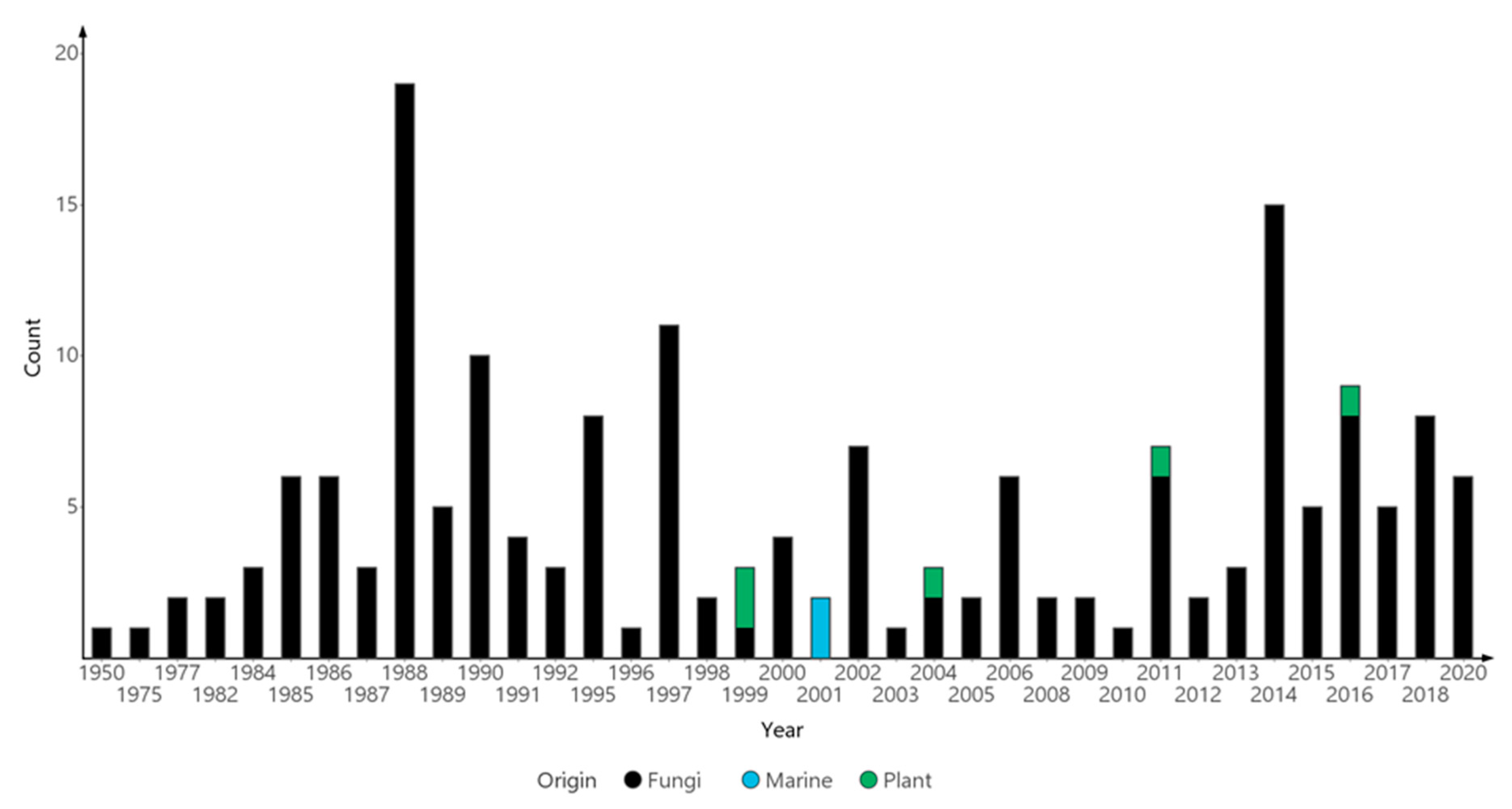
:1. Introduction
2. Basidiomycota
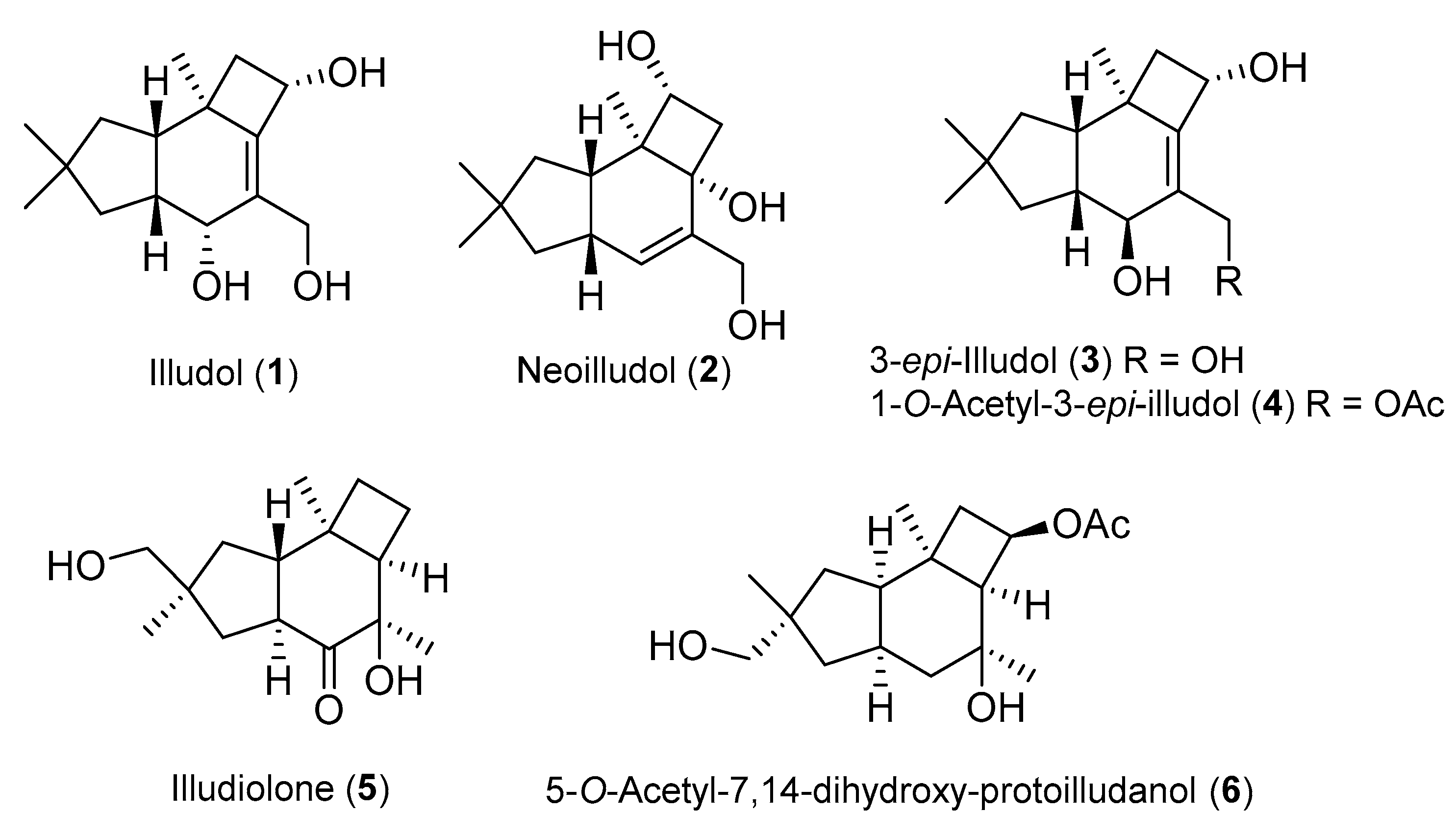
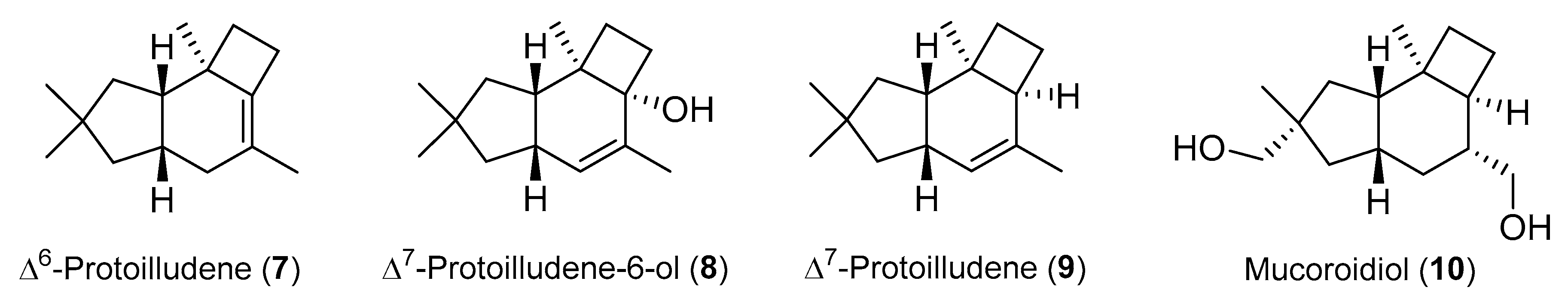
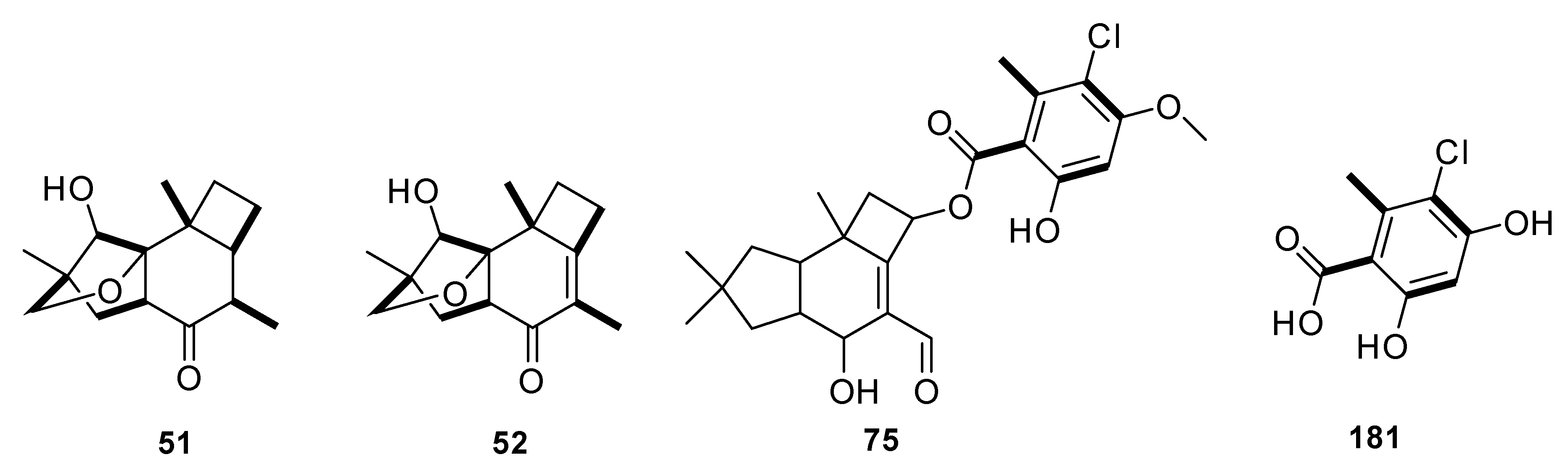
2.1. Protoilludane Sesquiterpenes
2.2. Protoilludane Sesquiterpene Aryl Esters
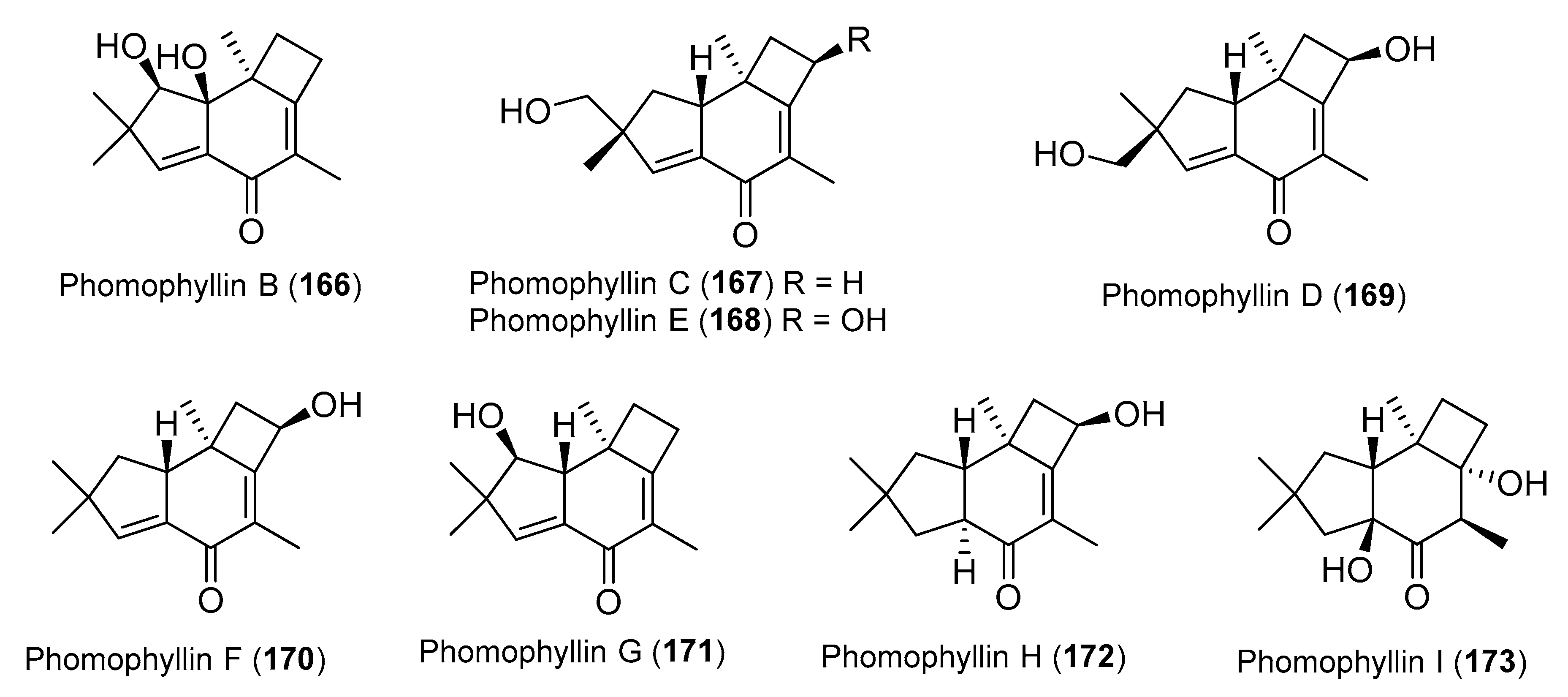
3. Ascomycota
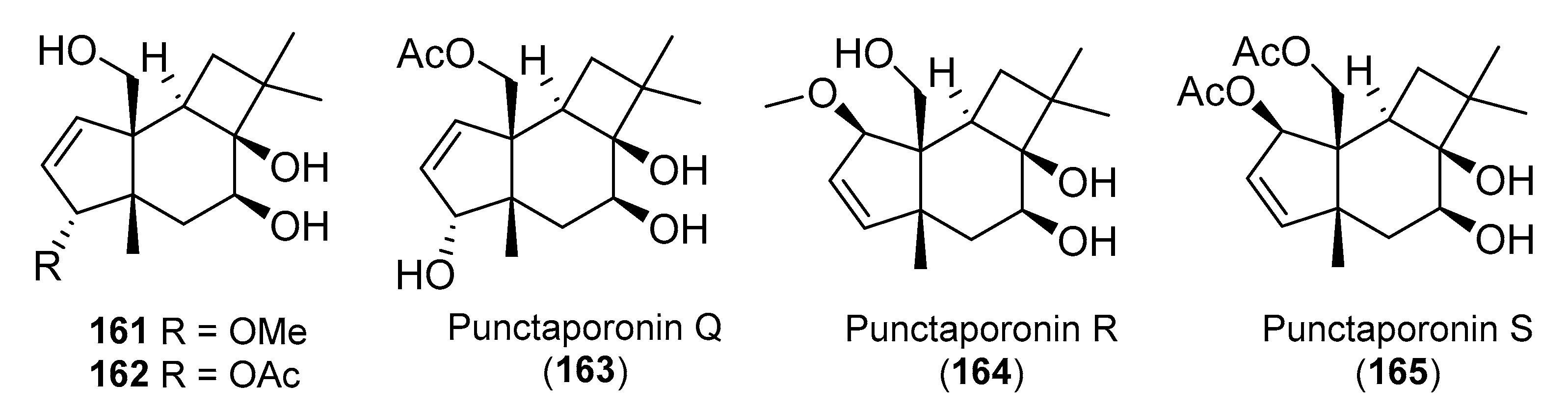
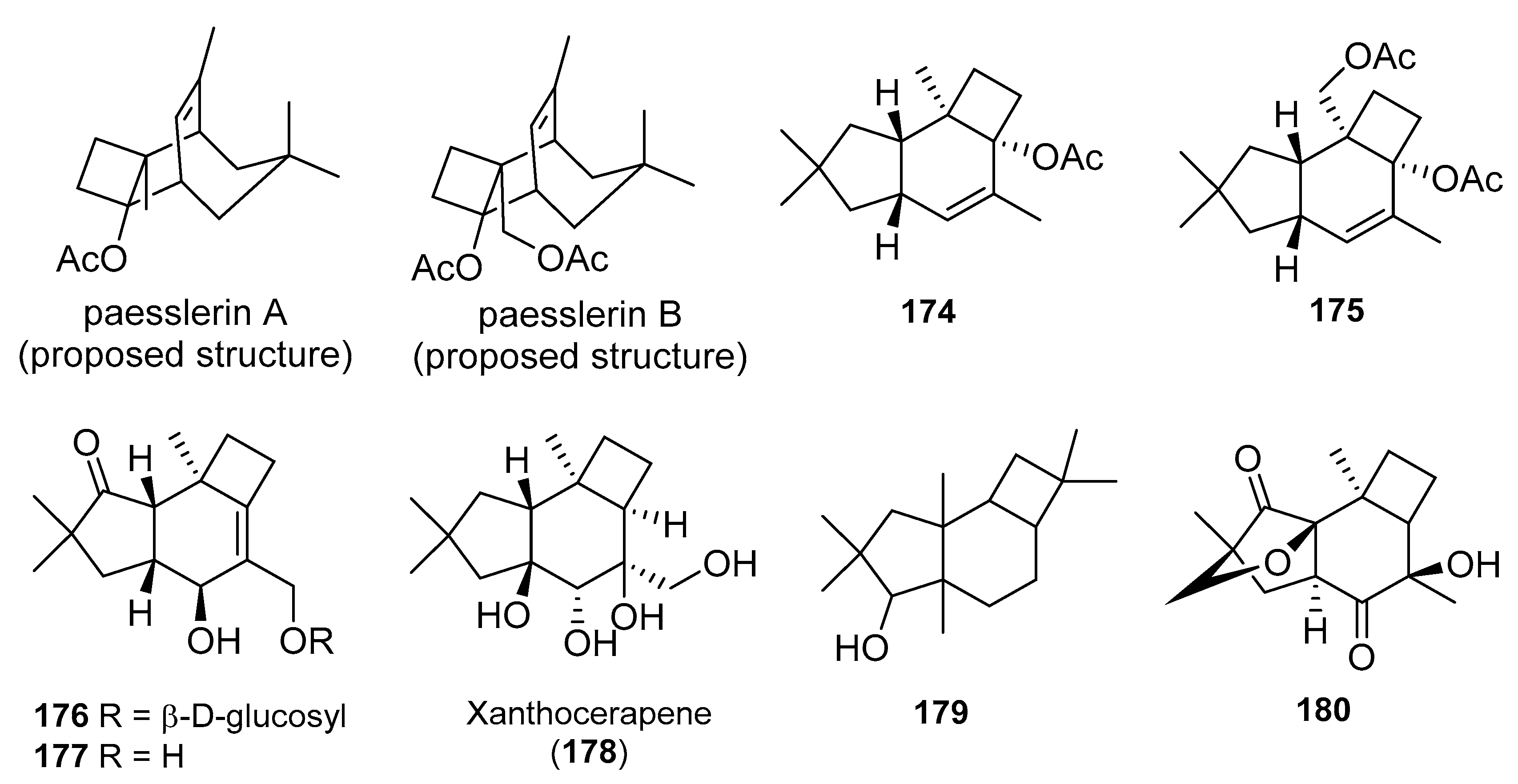
4. Protoilludanes from Other Sources
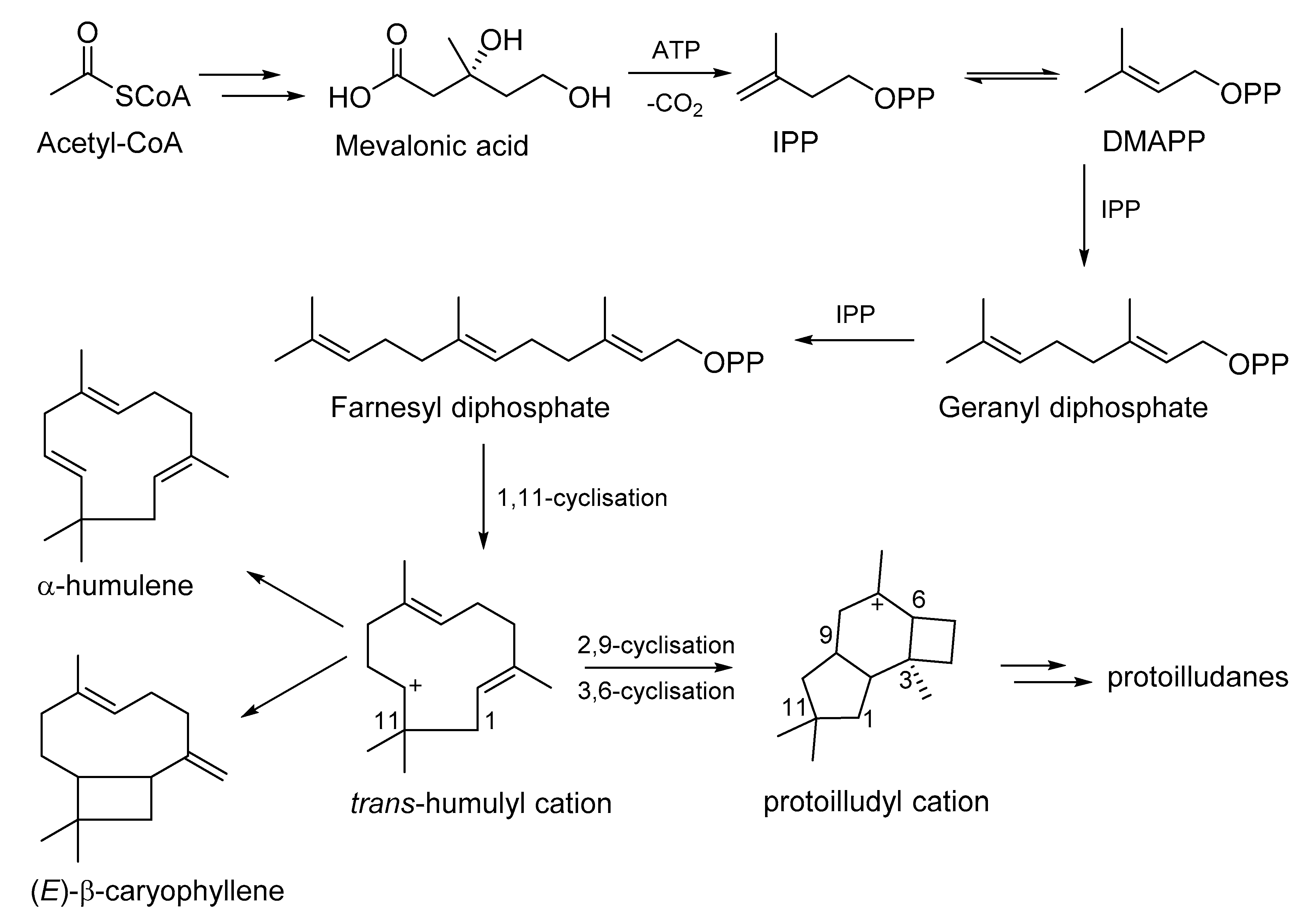
5. Biosynthetic Studies
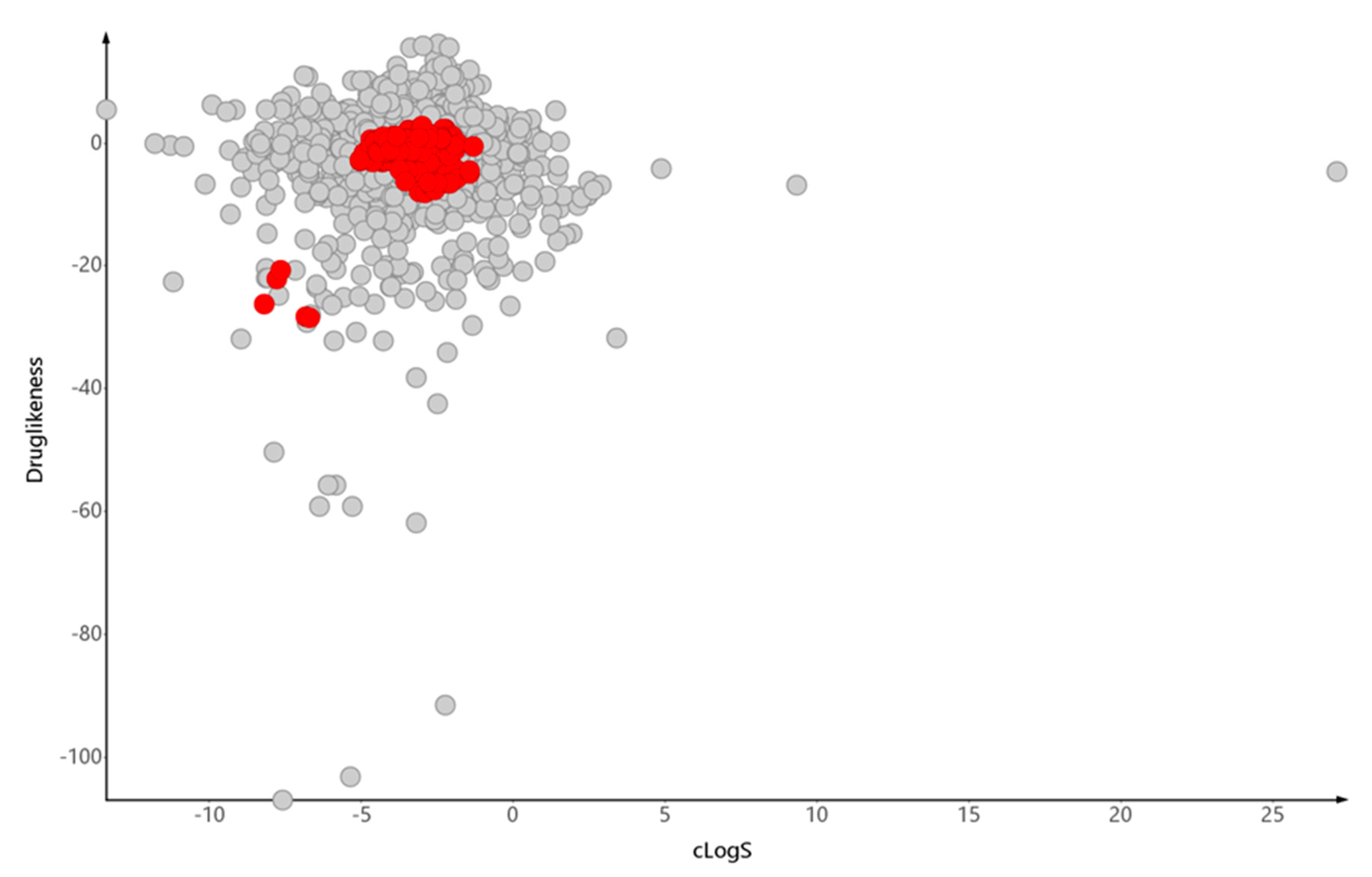
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.-J.; Bai, F.-W.; Zhang, W. (Eds.) Biotechnology in China I; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-88414-9. [Google Scholar]
- Schrader, J.; Bohlmann, J. (Eds.) Biotechnology of Isoprenoids; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Germany, 2015; Volume 148, ISBN 978-3-319-20106-1. [Google Scholar]
- Alves, M.J.; Ferreira, I.C.F.R.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A Review on Antimicrobial Activity of Mushroom (Basidiomycetes) Extracts and Isolated Compounds. Planta Med. 2012, 78, 1707–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Bass, D.; Richards, T.A. Three reasons to re-evaluate fungal diversity ‘on Earth and in the ocean’. Fungal Biol. Rev. 2011, 25, 159–164. [Google Scholar] [CrossRef]
- Hans, J.R. Natural Products: The Secondary Metabolites; Royal Society of Chemistry: Cambridge, UK, 2003; Volume 17, ISBN 0-85404-490-6. [Google Scholar]
- Mann, J. Chemical Aspects of Biosynthesis; Oxford Chemistry Primers, 1st ed.; Oxford University Press: Oxford, UK, 1994; ISBN 0-19-855676-4. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; John Wiley & Sons, Ltd.: England, UK, 2002; ISBN 0-471-49641-3. [Google Scholar]
- Dewick, P.M. The biosynthesis of C5–C25 terpenoid compounds. Nat. Prod. Rep. 2002, 19, 181–222. [Google Scholar] [CrossRef] [PubMed]
- Tibrewal, N.; Tang, Y. Biocatalysts for Natural Product Biosynthesis. Annu. Rev. Chem. Biomol. Eng. 2014, 5, 347–366. [Google Scholar] [CrossRef]
- Quin, M.B.; Flynn, C.M.; Schmidt-Dannert, C. Traversing the fungal terpenome. Nat. Prod. Rep. 2014, 31, 1449–1473. [Google Scholar] [CrossRef]
- Nord, C.L.; Menkis, A.; Broberg, A. Cytotoxic Illudane Sesquiterpenes from the Fungus Granulobasidium vellereum (Ellis and Cragin) Jülich. J. Nat. Prod. 2015, 78, 2559–2564. [Google Scholar] [CrossRef]
- Siengalewicz, P.; Mulzer, J.; Rinner, U. Synthesis of Protoilludanes and Related Sesquiterpenes. Eur. J. Org. Chem. 2011, 2011, 7041–7055. [Google Scholar] [CrossRef]
- Burgess, M.L.; Barrow, K.D. Biosynthesis of illudosin, a fomannosane-type sesquiterpene, by the Basidiomycete Omphalotus nidiformis. J. Chem. Soc. Perkin Trans. 1 1999, 1, 2461–2466. [Google Scholar] [CrossRef]
- Anchel, M.; Hervey, A.; Robbins, W.J. Antibiotic Substances from Basidiomycetes: VII. Clitocybe Illudens. Proc. Natl. Acad. Sci. USA 1950, 36, 300–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMorris, T.C.; Nair, M.S.R.; Anchel, M. Structure of illudol, a sesquiterpenoid triol from Clitocybe illudens. J. Am. Chem. Soc. 1967, 89, 4562–4563. [Google Scholar] [CrossRef]
- McMorris, T.C.; Nair, M.S.R.; Singh, P.; Anchel, M. The Structure of Illudol. Phytochemistry 1971, 10, 1611. [Google Scholar] [CrossRef]
- Matsumoto, T.; Miyano, K.; Kagawa, S.; Yi, S.; Ogawa, J.; Ichihara, A. Total Synthesis of dl-illudol. Tet. Lett. 1971, 38, 3521–3524. [Google Scholar] [CrossRef]
- Nair, M.S.R.; Anchel, M. Metabolic Products of Clitocybe Illudens XI. The Structure of Neoilludol. Tet. Lett. 1975, 14, 1267–1268. [Google Scholar] [CrossRef]
- Arnone, A.; Cardillo, R.; Modugno, V.D.; Nasini, G. Secondary Mould Metabolites. Part 29.’ Isolation and Structure Elucidation of Candicansol, 3-epi-Illudol and 1-O-Acetyl-3-epi-illudol, Novel Sesquiterpenoids from Clitocybe candicans, and Absolute Configuration of 3-epi-llludol. J. Chem. Soc. Perkin Trans. 1 1989, 11, 1995–2000. [Google Scholar] [CrossRef]
- Suzuki, S.; Murayama, T.; Shiono, Y. Echinolactones C and D: Two Illudalane Sesquiterpenoids Isolated from the Cultured Mycelia of the Fungus Echinodontium japonicum. Z. Naturforsch. 2006, 61b, 1295–1298. [Google Scholar] [CrossRef]
- Suzuki, S.; Murayama, T.; Shiono, Y. Illudalane sesquiterpenoids, echinolactones A and B, from a mycelial culture of Echinodontium japonicum. Phytochemistry 2005, 66, 2329–2333. [Google Scholar] [CrossRef]
- Elliott, M.R.; Dhimane, A.-L.; Malacria, M. Biomimetic Diastereoselective Total Synthesis of epi-Illudol via a Transannular Radical Cyclizations Strategy. J. Am. Chem. Soc. 1997, 119, 3427–3428. [Google Scholar] [CrossRef]
- McMorris, T.C.; Kashinatham, A.; Lira, R.; Rundgren, H.; Gantzel, P.K.; Kelner, M.J.; Dawe, R. Sesquiterpenes from Omphalotus illudens. Phytochemistry 2002, 61, 395–398. [Google Scholar] [CrossRef]
- Yang, X.-Y.; Li, Z.-H.; Dong, Z.-J.; Feng, T.; Liu, J.-K. Three new sesquiterpenoids from cultures of the basidiomycete Conocybe siliginea. J. Asian Nat. Prod. Res. 2014, 17, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nozoe, S.; Kobayashi, H.; Urano, S.; Furukawa, J. Isolation of Δ6-protoilludene and the related alcohols. Tetrahedron Lett. 1977, 18, 1381–1384. [Google Scholar] [CrossRef]
- Furukawa, J.; Morisaki, N.; Kobayashi, H.; Iwasaki, S.; Nozoe, S.; Okuda, S. Synthesis of dl-6-protoilludene. Chem. Pharm. Bull. 1985, 33, 440–443. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, H.; Iwabuchi, H.; Kouno, I.; Iino, M.; Nomura, D. Photochemical construction of the protoilludane skeleton: A total synthesis of protoillud-7-ene and some oxygenated derivatives. Chem. Lett. 1979, 8, 649–652. [Google Scholar] [CrossRef]
- Rabe, P.; Rinkel, J.; Nubbemeyer, B.; Köllner, T.G.; Chen, F.; Dickschat, J.S. Terpene Cyclases from Social Amoebae. Angew. Chem. Int. Ed. 2016, 55, 15420–15423. [Google Scholar] [CrossRef]
- Sasaki, H.; Kubohara, Y.; Ishigaki, H.; Takahashi, K.; Eguchi, H.; Sugawara, A.; Oshima, Y.; Kikuchi, H. Two New Terpenes Isolated from Dictyostelium Cellular Slime Molds. Molecules 2020, 25, 2895. [Google Scholar] [CrossRef]
- Arnone, A.; Nasini, G.; Assante, G.; Roeijmans, H.J.; Van Euk, G.W. Sulcatine, a norsesquiterpene from the fungus Laurilia sulcata. Phytochemistry 1987, 26, 1739–1742. [Google Scholar] [CrossRef]
- Arnone, A.; Nasini, G.; Assante, G.; Eijk, G.W. Three sesquiterpenes produced by the fungus Laurilia sulcata. Phytochemistry 1992, 31, 2047–2050. [Google Scholar] [CrossRef]
- Assante, G.; Dallavalle, S.; Martino, P.A. Protoilludane sesquiterpenoids as scaffold structures for new antimicrobials against Mannheimia haemolytica. J. Antibiot. 2012, 66, 43–45. [Google Scholar] [CrossRef]
- Arnone, A.; Brambilla, U.; Nasini, G.; De Pava, O.V. Isolation and structure elucidation of tsugicolines A-D, novel protoilludane sesquiterpenes from Laurilia tsugicola. Tetrahedron 1995, 51, 13357–13364. [Google Scholar] [CrossRef]
- Arnone, A.; De Gregorio, C.; Meille, S.V.; Nasini, G.; Sidoti, G. Tsugicoline E, a New Polyoxygenated Protoilludane Sesquiterpene from the Fungus Laurilia tsugicola. J. Nat. Prod. 1999, 62, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Shen, Y. Clavicorolides A and B, Sesquiterpenoids from the Fermentation Products of Edible Fungus Clavicorona pyxidata. Org. Lett. 2009, 11, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-B.; Lu, C.-H.; Zheng, Z.-H.; Lin, X.-J.; Su, W.-J.; Shen, Y.-M. New Sesquiterpenes from Edible Fungus Clavicorona pyxidata. Helv. Chim. Acta 2008, 91, 2174–2180. [Google Scholar] [CrossRef]
- Clericuzio, M.; Fu, J.; Pan, F.; Pang, Z.; Sterner, O. Structure and absolute configuration of protoilludane sesquiterpenes from Russula delica. Tetrahedron 1997, 53, 9735–9740. [Google Scholar] [CrossRef]
- Clericuzio, M.; Pan, F.; Han, F.; Pang, Z.; Sterner, O. Stearoyldelicone, an unstable protoilludane sesquiterpenoid from intact fruit bodies of Russula delica. Tetrahedron Lett. 1997, 38, 8237–8240. [Google Scholar] [CrossRef]
- Clericuzio, M.; Han, F.; Pan, F.; Pang, Z.; Sterner, O. The Sesquiterpenoid Contents of Fruit Bodies of Russula delica. Acta Chem. Scand. 1998, 52, 1333–1337. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, K.; Kaneko, A.; Matsumoto, Y.; Hama, H.; Arihara, S. Russujaponols A−F, Illudoid Sesquiterpenes from the Fruiting Body of Russula japonica. J. Nat. Prod. 2006, 69, 1267–1270. [Google Scholar] [CrossRef]
- Clericuzio, M.; Mella, M.; Toma, L.; Finzi, P.V.; Vidari, G. Atlanticones, New Protoilludane Sesquiterpenes from the Mushroom Lactarius atlanticus (Basidiomycetes). Eur. J. Org. Chem. 2002, 6, 988–994. [Google Scholar] [CrossRef]
- Zech, M.S.A.; Jandl, C.; Bach, T. Concise Access to the Skeleton of Protoilludane Sesquiterpenes through a Photochemical Reaction Cascade: Total Synthesis of Atlanticone C. Angew. Chem. Int. Ed. 2019, 58, 14629–14632. [Google Scholar] [CrossRef]
- Bach, T.; Proessdorf, J.; Zech, A.; Jandl, C. Concise Total Synthesis of (+)-Atlanticone C. Synlett 2020, 31, 1598–1602. [Google Scholar] [CrossRef]
- Hirota, M.; Shimizu, Y.; Kamo, T.; Makabe, H.; Shibata, H. New Plant Growth Promoters, Repraesentins A, B and C, from Lactarius repraesentaneus. Biosci. Biotechnol. Biochem. 2003, 67, 1597–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Iinuma, H.; Momose, I.; Matsui, S. Antibiotic Pasteurestin A and B and their Manufacture with Agrocybe cylindracea. JP Patent 2002212137, 31 July 2002. [Google Scholar]
- Surup, F.; Hennicke, F.; Sella, N.; Stroot, M.; Bernecker, S.; Pfütze, S.; Stadler, M.; Rühl, M. New terpenoids from the fermentation broth of the edible mushroom Cyclocybe aegerita. Beilstein J. Org. Chem. 2019, 15, 1000–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kögl, M.; Brecker, L.; Warrass, R.; Mulzer, J. Total Synthesis and Configurational Assignment of Pasteurestin A and B. Angew. Chem. Int. Ed. 2007, 46, 9320–9322. [Google Scholar] [CrossRef] [PubMed]
- Kögl, M.; Brecker, L.; Warrass, R.; Mulzer, J. Novel Protoilludane Lead Structure for Veterinary Antibiotics: Total Synthesis of Pasteurestins A and B and Assignment of Their Configurations. Eur. J. Org. Chem. 2008, 2008, 2714–2730. [Google Scholar] [CrossRef]
- Li, H.-T.; Tang, L.-H.; Liu, T.; Yang, R.-N.; Yang, Y.-B.; Zhou, H.; Ding, Z.-T. Protoilludane-type sesquiterpenoids from Armillaria sp. by co-culture with the endophytic fungus Epicoccum sp. associated with Gastrodia elata. Bioorganic Chem. 2020, 95, 103503. [Google Scholar] [CrossRef]
- Stärk, A.; Anke, T.; Mocek, U.; Steglich, W.; Kirfel, A.; Will, G. Lentinellic Acid, a Biologically Active Protoilludane Derivative from Lentinellus Species (Basidiomycetes). Z. Naturforsch. C 1988, 43, 177–183. [Google Scholar] [CrossRef]
- Wunder, A.; Anke, T.; Klostermeyer, D.; Steglich, W. Lactarane Type Sesquiterpenoids as Inhibitors of Leukotriene Biosynthesis and Other, New Metabolites from Submerged Cultures of Lentinellus cochleatus (Pers. ex Fr.) Karst. Z. Naturforsch. C 1996, 51, 493–499. [Google Scholar] [CrossRef]
- Fabian, K.; Lorenzen, K.; Anke, T.; Johansson, M.; Sterner, O. Five new bioactive sesquiterpenes from the fungus Radulomyces confluens (Fr.) Christ. Z. Naturforsch. C 1999, 53, 939–945. [Google Scholar] [CrossRef]
- Chang, E.L.; Bolte, B.; Lan, P.; Willis, A.C.; Banwell, M.G. Chemoenzymatic Total Syntheses of the Enantiomers of the Protoilludanes 8-Deoxydihydrotsugicoline and Radudiol. J. Org. Chem. 2016, 81, 2078–2086. [Google Scholar] [CrossRef]
- Starratt, A.N.; Stothers, J.B.; Ward, E.W.B. Coprinolone, an oxygen-bridged protoilludane from the fungus Coprinus psychromorbidus: Structure determination by chemical and n.m.r. studies aided by biosynthetic incorporation of [1,2-13C2]acetate. J. Chem. Soc. Chem. Commun. 1988, 590–591. [Google Scholar] [CrossRef]
- Starratt, A.N.; Ward, E.W.B.; Stothers, J.B. Coprinolone and Δ6-coprinolone: New sesquiterpenes from Coprinus psychromorbidus. Can. J. Chem. 1989, 67, 417–427. [Google Scholar] [CrossRef]
- Nord, C.L.; Menkis, A.; Vasaitis, R.; Broberg, A. Protoilludane sesquiterpenes from the wood decomposing fungus Granulobasidium vellereum (Ellis & Cragin) Jülich. Phytochemistry 2013, 90, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Nord, C.L.; Menkis, A.; Lendel, C.; Vasaitis, R.; Broberg, A. Sesquiterpenes from the saprotrophic fungus Granulobasidium vellereum (Ellis & Cragin) Jülich. Phytochemistry 2014, 102, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Shiono, Y.; Seto, T.; Kamata, M.; Takita, C.; Suzuki, S.; Murayama, T.; Ikeda, M. Protoilludane-Type Sesquiterpenes, Echinocidins A and B, from a Mycelial Culture of Echinodontium tsugicola. Z. Naturforsch. B 2004, 59, 925–929. [Google Scholar] [CrossRef]
- Shiono, Y.; Suzuki, S.; Murayama, T.; Ikeda, M.; Abe, Y. Protoilludane Sequiterpenoids, Echinocidins C and D Produced by a Decay Causing Fungal Strain Echinodontium tsugicola. Z. Naturforsch. B 2005, 60, 449–452. [Google Scholar] [CrossRef]
- Shimoda, K.; Yamaoka, Y.; Yoo, D.; Yamada, K.-I.; Takikawa, H.; Takasu, K. Total Syntheses of Allelopathic 4-Oxyprotoilludanes, Melleolides, and Echinocidins. J. Org. Chem. 2019, 84, 11014–11024. [Google Scholar] [CrossRef]
- Donnelly, D.; Sanada, S.; O’Reilly, J.; Polonky, J.; Prangé, T.; Pascard, C. Isolation and structure (X-ray analysis) of the orsellinate of armillol, a new antibacterial metabolite from Armillaria mellea. J. Chem. Soc. Chem. Commun. 1982, 135–137. [Google Scholar] [CrossRef]
- Donnelly, D.M.X.; Abe, F.; Coveney, D.; Fukuda, N.; O’Reilly, J.; Polonsky, J.; Prangé, T. Antibacterial Sesquiterpene Aryl Esters from Armillaria mellea. J. Nat. Prod. 1985, 48, 10–16. [Google Scholar] [CrossRef]
- Donnelly, D.M.X.; Coveney, D.J.; Fukuda, N.; Polonsky, J. New Sesquiterpene Aryl Esters from Armillaria mellea. J. Nat. Prod. 1986, 49, 111–116. [Google Scholar] [CrossRef]
- Yang, J.-S.; Cong, P.-Z. Mass Spectrometric Studies on the Sesquiterpenol Aromatic Esters from Mycelium of Armillaria Mellea. Huaxue Xuebao 1988, 46, 1093–1100. [Google Scholar]
- Yang, J.S.; Chen, Y.W.; Feng, X.Z.; Yu, D.Q.; He, C.H.; Zheng, Q.T.; Liang, X.T. Isolation and Structure Elucidation of Armillaricin. Planta Med. 2007, 55, 564–565. [Google Scholar] [CrossRef]
- Misiek, M.; Williams, J.; Schmich, K.; Hüttel, W.; Merfort, I.; Salomon, C.E.; Aldrich, C.C.; Hoffmeister, D. Structure and Cytotoxicity of Arnamial and Related Fungal Sesquiterpene Aryl Esters. J. Nat. Prod. 2009, 72, 1888–1891. [Google Scholar] [CrossRef]
- Bohnert, M.; Miethbauer, S.; Dahse, H.-M.; Ziemen, J.; Nett, M.; Hoffmeister, D. In vitro cytotoxicity of melleolide antibiotics: Structural and mechanistic aspects. Bioorganic Med. Chem. Lett. 2011, 21, 2003–2006. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, M.; Nuetzmann, H.-W.; Eschroeckh, V.; Horn, F.; Dahse, H.-M.; Brakhage, A.A.; Hoffmeister, D. Cytotoxic and antifungal activities of melleolide antibiotics follow dissimilar structure–activity relationships. Phytochemistry 2014, 105, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.M.X.; Polonsky, J.; Prangé, T.; Snatzke, G.; Wagner, U. The absolute configuration of the orsellinate of armillol; application of the coupled oscillator theory. J. Chem. Soc. Chem. Commun. 1984, 222–223. [Google Scholar] [CrossRef]
- Cremin, P.; Donnelly, D.M.X.; Guiry, P.J.; Wolfender, J.-L.; Hostettmann, K. A liquid chromatography–thermospray ionisation–mass spectrometry guided isolation of a new sesquiterpene aryl ester from Armillaria novae-zelandiae. J. Chem. Soc. Perkin Trans. 1 2000, 1, 2325–2329. [Google Scholar] [CrossRef]
- Misiek, M.; Hoffmeister, D. Sesquiterpene aryl ester natural products in North American Armillaria species. Mycol. Prog. 2010, 11, 7–15. [Google Scholar] [CrossRef]
- Chen, C.-C.; Kuo, Y.-H.; Cheng, J.-J.; Sung, P.-J.; Ni, C.-L.; Chen, C.-C.; Shen, C.-C. Three New Sesquiterpene Aryl Esters from the Mycelium of Armillaria mellea. Molecules 2015, 20, 9994–10003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midland, S.L.; Izac, R.R.; Wing, R.M.; Zaki, A.I.; Munnecke, D.E.; Sims, J.J. Melleolide, A new Antibiotic from Armillaria mellea. Tet. Lett. 1982, 23, 2515–2518. [Google Scholar] [CrossRef]
- Arnone, A.; Cardillo, R.; Nasini, G. Structures of melleolides B-D, three antibacterial sesquiterpenoids from Armillaria mellea. Phytochemistry 1986, 25, 471–474. [Google Scholar] [CrossRef]
- Arnone, A.; Cardillo, R.; Di Modugno, V.; Nasini, G. Secondary mould metabolites. XXII. Isolation and structure elucidation of melledonals D and E and melleolides E-H, novel sesquiterpenoid aryl esters from Clitocybe elegans and Armillaria mellea. Gazz. Chim. Ital. 1988, 118, 517–521. [Google Scholar]
- Momose, I.; Sekizawa, R.; Hosokawa, N.; Iinuma, H.; Maisui, S.; Nakamura, H.; Naganawa, H.; Hamada, M.; Takeuchi, T. Melleolides K, L and M, New Melleolides from Armillariella mellea. J. Antibiot. 2000, 53, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-C.; Cheng, J.-J.; Shen, C.-C. Protoilludane Norsesquiterpenoid Esters and Uses Thereof. U.S. Patent 20110262561, 27 October 2011. [Google Scholar]
- Arnone, A.; Cardillo, R.; Nasini, G. Secondary mould metabolites. XXIII. Isolation and structure elucidation of melleolides I and J and armellides A and B, novel sesquiterpenoid aryl esters from Armillaria novae-zelandiae. Gazz. Chim. Ital. 1988, 118, 523–527. [Google Scholar]
- Yang, J.; Su, Y.; Wang, Y.; Feng, X.; Yu, D.; Cong, P.; Tamai, M.; Obuchi, T.; Kondoh, H.; Liang, X. Isolation and Structures of Two New Sesquiterpenoid Aromatic Esters: Armillarigin and Armillarikin1. Planta Medica 1989, 55, 479–481. [Google Scholar] [CrossRef]
- Arnone, A.; Cardillo, R.; Nasini, G.; Meille, S.V. Secondary mould metabolites. Part 19. Structure elucidation and absolute configuration of melledonals B and C, novel antibacterial sesquiterpenoids from Armillaria mellea. X-Ray molecular structure of melledonal C. J. Chem. Soc. Perkin Trans. 1 1988, 1, 503. [Google Scholar] [CrossRef]
- Kobori, H.; Sekiya, A.; Suzuki, T.; Choi, J.-H.; Hirai, H.; Kawagishi, H. Bioactive Sesquiterpene Aryl Esters from the Culture Broth of Armillaria sp. J. Nat. Prod. 2015, 78, 163–167. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Jiang, B.; Li, W.; Zheng, L.; Yang, X.; Bao, Y.; Sun, L.; Huang, Y.-X.; Li, Y. Structure, cytotoxic activity and mechanism of protoilludane sesquiterpene aryl esters from the mycelium of Armillaria mellea. J. Ethnopharmacol. 2016, 184, 119–127. [Google Scholar] [CrossRef]
- Yin, X.; Feng, T.; Liu, J.-K. Structures and cytotoxicities of three new sesquiterpenes from cultures of Armillaria sp. Nat. Prod. Bioprospect. 2012, 2, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Cremin, P.; Donnelly, D.M.; Wolfender, J.-L.; Hostettmann, K. Liquid chromatographic-thermospray mass spectrometric analysis of sesquiterpenes of Armillaria (Eumycota: Basidiomycotina) species. J. Chromatogr. A 1995, 710, 273–285. [Google Scholar] [CrossRef]
- Donnelly, D.M.; Hutchinson, R.M.; Coveney, D.; Yonemitsu, M. Sesquiterpene aryl esters from Armillaria mellea. Phytochemistry 1990, 29, 2569–2572. [Google Scholar] [CrossRef]
- Donnelly, D.M.; Konishi, T.; Dunne, O.; Cremin, P. Sesquiterpene aryl esters from Armillaria tabescens. Phytochemistry 1997, 44, 1473–1478. [Google Scholar] [CrossRef]
- Donnelly, D.M.; Coveney, D.J.; Polonsky, J. Melledonal and melledonol, sesquiterpene esters from Armillaria mellea. Tetrahedron Lett. 1985, 26, 5343–5344. [Google Scholar] [CrossRef]
- Donnelly, D.M.; Quigley, P.F.; Coveney, D.J.; Polonsky, J. Two new sesquiterpene esters from Armillaria mellea. Phytochemistry 1987, 26, 3075–3077. [Google Scholar] [CrossRef]
- Yang, J.; Yuwu, C.; Xiaozhang, F.; Dequan, Y.; XiaoTian, L. Chemical Constituents of Armillaria mellea Mycelium I. Isolation and Characterization of Armillarin and Armillaridin. Planta Medica 1984, 50, 288–290. [Google Scholar] [CrossRef]
- Yang, J.S.; Su, Y.L.; Wang, Y.L.; Feng, X.Z.; Yu, D.Q.; Liang, X.T. Studies on the chemical constituents of Armillaria mellea mycelium. V. Isolation and characterization of armillarilin and armillarinin. Yao Xue Xue Bao 1990, 25, 24–28. [Google Scholar]
- Yang, J.S.; Su, Y.L.; Wang, Y.L.; Feng, X.Z.; Yu, D.Q.; Liang, X.T.; He, C.H.; Zheng, Q.T. Chemical constituents of Armillaria mellea mycelium. VI. Isolation and structure of armillaripin. Yao Xue Xue Bao 1990, 25, 353–356. [Google Scholar]
- Yang, J.S.; Su, Y.L.; Wang, Y.L.; Feng, X.Z.; Yu, D.Q.; Liang, X.T. Chemical constituents of Armillaria mellea mycelium. VII. Isolation and characterization of chemical constituents of the acetone extract. Yao Xue Xue Bao 1991, 26, 117–122. [Google Scholar]
- Yang, J.; Su, Y.; Wang, Y.; Feng, X.; Yu, D.; Liang, X. Two Novel Protoilludane Norsesquiterpenoid Esters, Armillasin and Armillatin, from Armillaria mellea. Planta Med. 1991, 57, 478–480. [Google Scholar] [CrossRef]
- Yang, J.S.; Watube, S. Antibiotic Armillaric Acid and Its Manufacture with Armillaria mellea. CN Patent 1034956, 23 August 1989. [Google Scholar]
- Donnelly, D.M.; Hutchinson, R.M. Armillane, a saturated sesquiterpene ester from Armillaria mellea. Phytochemistry 1990, 29, 179–182. [Google Scholar] [CrossRef]
- Obuchi, T.; Kondoh, H.; Watanabe, N.; Tamai, M.; Imura, S.; Jun-Shan, Y.; Xiao-Tian, L. Armillaric Acid, A New Antibiotic Produced by Armillaria mellea. Planta Med. 1990, 56, 198–201. [Google Scholar] [CrossRef]
- Sonnenbichler, J.; Guillaumin, J.-J.; Peipp, H.; Schwarz, D. Secondary metabolites from dual cultures of genetically different Armillaria isolates. For. Pathol. 1997, 27, 241–249. [Google Scholar] [CrossRef]
- Hanssen, H.-P.; Sprecher, E.; Abraham, W.-R. 6-Protoilludene, the major volatile metabolite from ceratocystis piceae liquid cultures. Phytochemistry 1986, 25, 1979–1980. [Google Scholar] [CrossRef]
- Anderson, J.R.; Briant, C.E.; Edwards, R.L.; Mabelis, R.P.; Poyser, J.P.; Spencer, H.; Whalley, A.J.S. Punctatin A (antibiotic M95464): X-ray crystal structure of a sesquiterpene alcohol with a new carbon skeleton from the fungus, Paronia punctata. J. Chem. Soc., Chem. Commun. 1984, 7, 405–406. [Google Scholar] [CrossRef]
- Poyser, J.P.; Edwards, R.L.; Anderson, J.R.; Hursthouse, M.B.; Walker, N.P.C.; Sheldrick, G.M.; Whalley, A.J.S. Punctatins A,D,E, and F (antibiotics M95464, M167906, M171950, and M189122), isomeric allylic alcohols from the fungus Poronia punctata: X-ray crystal structures of D and of E acetonide. J. Antibiot. 1986, 39, 167–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.R.; Edwards, R.L.; Poyser, J.P.; Whalley, A.J.S. Metabolites of the higher fungi. Part 23. The punctaporonins. Novel bi-, tri-, and tetra-cyclic sesquiterpenes related to caryophyllene, from the fungus Poronia punctate (Linnaeus:Fries) Fries. J. Chem. Soc. Perkin Trans. 1 1988, 1, 823. [Google Scholar] [CrossRef]
- Sugimura, T.; Paquette, L.A. Enantiospecific total synthesis of the sesquiterpene antibiotics (-)-punctatin A and (+)-punctatin D. J. Am. Chem. Soc. 1987, 109, 3017–3024. [Google Scholar] [CrossRef]
- Deyrup, S.T.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T. Caryophyllene Sesquiterpenoids from a Fungicolous Isolate of Pestalotiopsis disseminata. J. Nat. Prod. 2006, 69, 608–611. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Q.; Liu, X.; Che, Y. Punctaporonins N–S, New Caryophyllene Sesquiterpenoids from Cytospora sp. BioMed Res. Int. 2017, 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.-H.; Liu, D.; Proksch, P.; Guo, P.; Lin, W. Punctaporonins H–M: Caryophyllene-Type Sesquiterpenoids from the Sponge-Associated Fungus Hansfordia sinuosae. Mar. Drugs 2014, 12, 3904–3916. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.H.; Wicklow, D.T.; Gloer, J.B. New punctaporonins from two fungicolous isolates of Pestalotiopsis sp. Phytochem. Lett. 2016, 16, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Wu, Y.; Qiao, Y.; Guo, Y.; Wang, J.; Hu, Z.; Zhang, Q.; Li, X.; Huang, J.; Zhou, Q.; et al. Protoilludane, Illudalane, and Botryane Sesquiterpenoids from the Endophytic Fungus Phomopsis sp. TJ507A. J. Nat. Prod. 2018, 81, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Brasco, M.F.R.; Seldes, A.M.; Palermo, J.A. Paesslerins A and B: Novel Tricyclic Sesquiterpenoids from the Soft Coral Alcyonium aessleri. Org. Lett. 2001, 3, 1415–1417. [Google Scholar] [CrossRef] [PubMed]
- Inanaga, K.; Takasu, K.; Ihara, M. Rapid Assembly of Polycyclic Substances by a Multicomponent Cascade (4 + 2)−(2 + 2) Cycloadditions: Total Synthesis of the Proposed Structure of Paesslerin A. J. Am. Chem. Soc. 2004, 126, 1352–1353. [Google Scholar] [CrossRef] [PubMed]
- Mogi, Y.; Inanaga, K.; Tokuyama, H.; Ihara, M.; Yamaoka, Y.; Yamada, K.-I.; Takasu, K. Rapid Assembly of Protoilludane Skeleton through Tandem Catalysis: Total Synthesis of Paesslerin A and Its Structural Revision. Org. Lett. 2019, 21, 3954–3958. [Google Scholar] [CrossRef] [PubMed]
- Castillo, U.F.; Sakagami, Y.; Alonso-Amelot, M.; Ojika, M. Pteridanoside, the first protoilludane sesquiterpene glucoside as a toxic component of the neotropical bracken fern Pteridium aquilinum var. caudatum. Tetrahedron 1999, 55, 12295–12300. [Google Scholar] [CrossRef]
- Ma, C.M.; Nakamura, N.; Nawawi, A.; Hattori, M.; Cai, S.Q. A Novel Protoilludane Sesquiterpene from the Wood of Xanthoceras sorbifolia. Chin. Chem. Lett. 2004, 15, 65–67. [Google Scholar]
- Wang, J.; Zhang, L.-X.; Zhao, Y.; Chen, W.-X.; Yang, Q.; Wang, Y.-X. Volatile Oil Contents and Their Variation Law of RADIX ET RHIZOMA GINSENG in Changbai Mountain Areas. Med. Plant 2011, 2, 40–43, 51. [Google Scholar]
- Jiao, H.-Y.; Wang, Y.-S.; Mo, X.-L.; Zhang, X.-T.; Zeng, Q.-Q. Analysis of volatile components of Pogostemon cablin from Indonesia and China. Jinri Yaoxue 2013, 23, 1–3, 10. [Google Scholar]
- Tong, H.-F.; Xue, J.; Tong, Y.-L. Analysis of volatile components from ginseng and American ginseng for identification by GC-MS. Zhongyiyao Xuebao 2013, 41, 49–54. [Google Scholar]
- Wang, W.; Liu, X.; Liu, J.; Cai, E.; Zhao, Y.; Li, H.; Zhang, L.; Li, P.; Gao, Y. Sesquiterpenoids from the Root of Panax ginseng Attenuates Lipopolysaccharide-Induced Depressive-Like Behavior through the Brain-Derived Neurotrophic Factor/Tropomyosin-Related Kinase B and Sirtuin Type 1/Nuclear Factor-κB Signaling Pathways. J. Agric. Food Chem. 2017, 66, 265–271. [Google Scholar] [CrossRef]
- Wu, J. Protoilludane Sesquiterpenoid Compound, Preparation Method and Medical Applications Thereof. CN Patent 105418544, 23 March 2016. [Google Scholar]
- Sander, T.; Freyss, J.; Von Korff, M.; Rufener, C. DataWarrior: An Open-Source Program for Chemistry Aware Data Visualization and Analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef] [PubMed]































Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadelis, M.M.; Copp, B.R.; Wiles, S. A Review of Fungal Protoilludane Sesquiterpenoid Natural Products. Antibiotics 2020, 9, 928. https://doi.org/10.3390/antibiotics9120928
Cadelis MM, Copp BR, Wiles S. A Review of Fungal Protoilludane Sesquiterpenoid Natural Products. Antibiotics. 2020; 9(12):928. https://doi.org/10.3390/antibiotics9120928
Chicago/Turabian StyleCadelis, Melissa M., Brent R. Copp, and Siouxsie Wiles. 2020. "A Review of Fungal Protoilludane Sesquiterpenoid Natural Products" Antibiotics 9, no. 12: 928. https://doi.org/10.3390/antibiotics9120928
APA StyleCadelis, M. M., Copp, B. R., & Wiles, S. (2020). A Review of Fungal Protoilludane Sesquiterpenoid Natural Products. Antibiotics, 9(12), 928. https://doi.org/10.3390/antibiotics9120928