Fosfomycin Resistance in Escherichia coli Isolates from South Korea and in vitro Activity of Fosfomycin Alone and in Combination with Other Antibiotics



Abstract
:1. Introduction
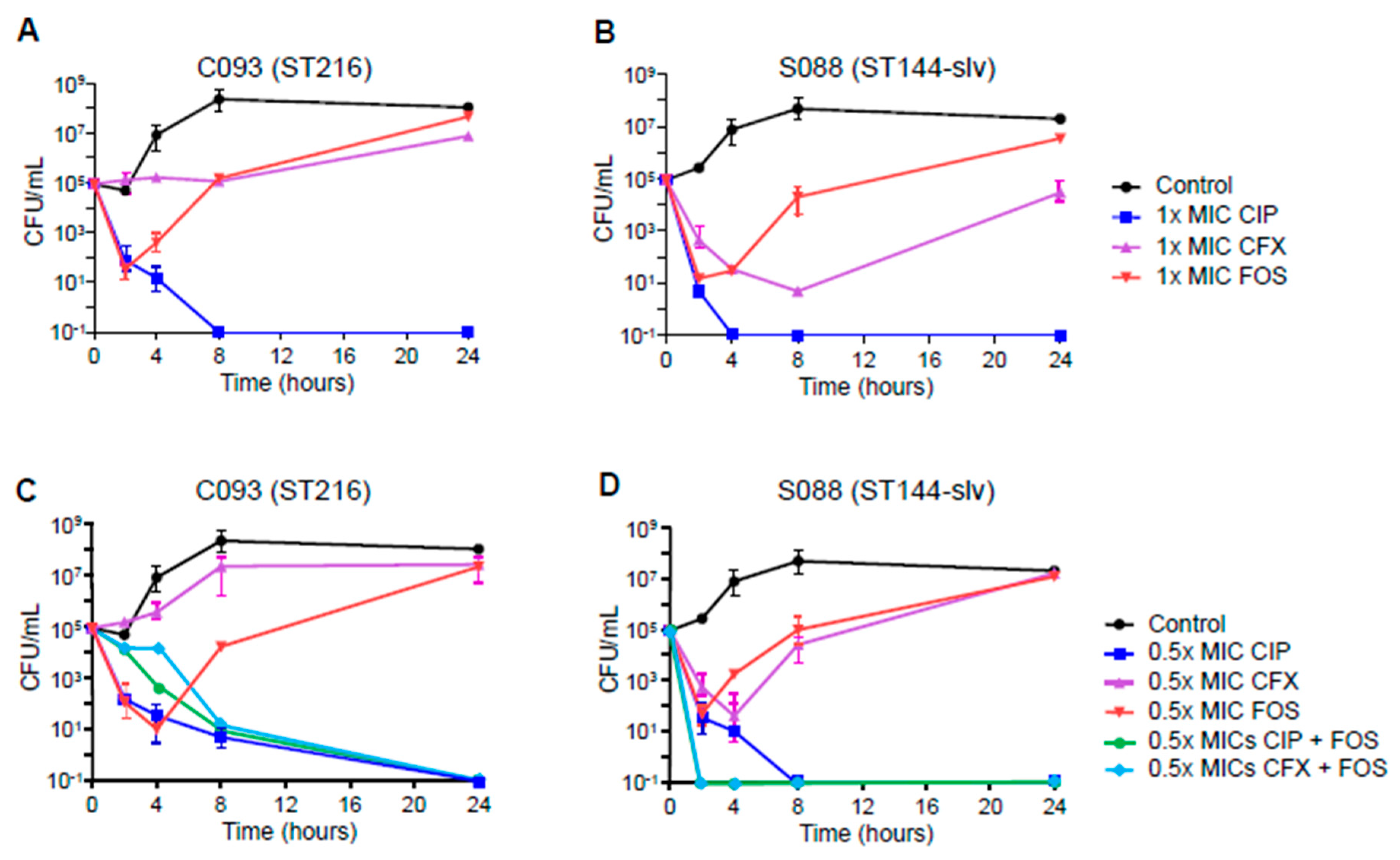
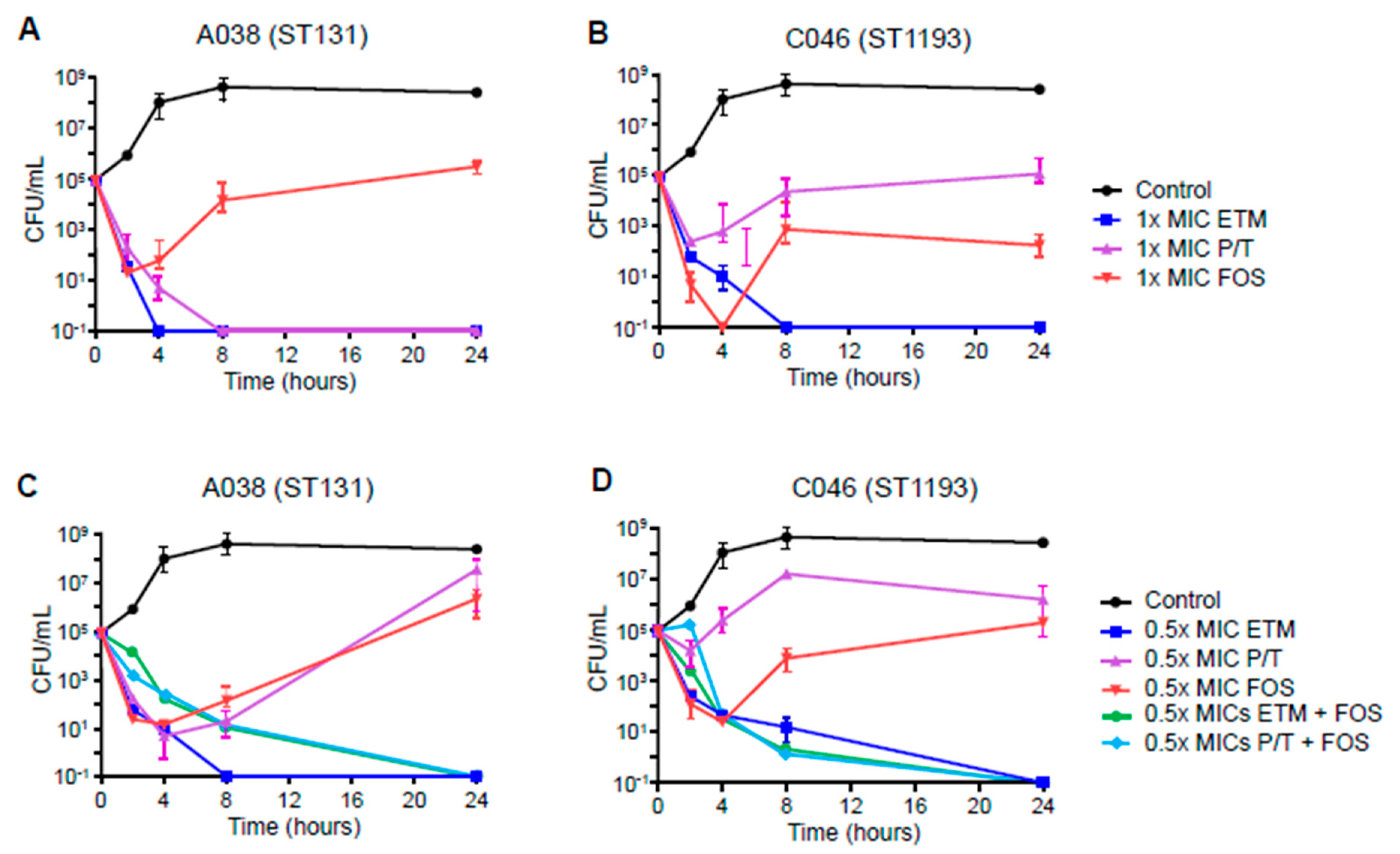
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Genotyping
4.4. Fosfomycin Resistance Mechanisms
4.5. In vitro Time-Kill Assays
4.6. Definition of Infection Onset
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bouchillon, S.K.; Badal, R.E.; Hoban, D.J.; Hawser, S.P. Antimicrobial susceptibility of inpatient urinary tract isolates of gram-negative bacilli in the United States: Results from the study for monitoring antimicrobial resistance trends (SMART) program: 2009–2011. Clin. Ther. 2013, 35, 872–877. [Google Scholar] [CrossRef]
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113, 5–13. [Google Scholar] [CrossRef]
- Vila, J.; Saez-Lopez, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giske, C.G. Contemporary resistance trends and mechanisms for the old antibiotics colistin, temocillin, fosfomycin, mecillinam and nitrofurantoin. Clin. Microbiol. Infect. 2015, 21, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahan, F.M.; Kahan, J.S.; Cassidy, P.J.; Kropp, H. The mechanism of action of fosfomycin (phosphonomycin). Ann. N. Y. Acad. Sci. 1974, 235, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Sastry, S.; Doi, Y. Fosfomycin: Resurgence of an old companion. J. Infect. Chemother. 2016, 22, 273–280. [Google Scholar] [CrossRef]
- Falagas, M.E.; Giannopoulou, K.P.; Kokolakis, G.N.; Rafailidis, P.I. Fosfomycin: Use beyond urinary tract and gastrointestinal infections. Clin. Infect. Dis. 2008, 46, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Castaneda-Garcia, A.; Blazquez, J.; Rodriguez-Rojas, A. Molecular Mechanisms and Clinical Impact of Acquired and Intrinsic Fosfomycin Resistance. Antibiotics (Basel) 2013, 2, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Athanasaki, F.A.; Voulgaris, G.L.; Triarides, N.A.; Vardakas, K.Z. Resistance to fosfomycin: Mechanisms, frequency and clinical consequences. Int. J. Antimicrob. Agents 2019, 53, 22–28. [Google Scholar] [CrossRef]
- Takahata, S.; Ida, T.; Hiraishi, T.; Sakakibara, S.; Maebashi, K.; Terada, S.; Muratani, T.; Matsumoto, T.; Nakahama, C.; Tomono, K. Molecular mechanisms of fosfomycin resistance in clinical isolates of Escherichia coli. Int. J. Antimicrob. Agents 2010, 35, 333–337. [Google Scholar] [CrossRef]
- Kresken, M.; Pfeifer, Y.; Hafner, D.; Wresch, R.; Korber-Irrgang, B. Occurrence of multidrug resistance to oral antibiotics among Escherichia coli urine isolates from outpatient departments in Germany: Extended-spectrum beta-lactamases and the role of fosfomycin. Int. J. Antimicrob. Agents 2014, 44, 295–300. [Google Scholar] [CrossRef]
- Mendes, A.C.; Rodrigues, C.; Pires, J.; Amorim, J.; Ramos, M.H.; Novais, Â.; Peixe, L. Importation of fosfomycin resistance fosA3 gene to Europe. Emerg. Infect. Dis. 2016, 22, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, B.; Li, Y.; Zhu, S.; Xue, F.; Liu, J. Antimicrobial susceptibility and molecular mechanisms of fosfomycin resistance in clinical Escherichia coli isolates in mainland China. PLoS ONE 2015, 10, e0135269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, K.S.; Suh, J.Y.; Peck, K.R.; Lee, M.Y.; Oh, W.S.; Kwon, K.T.; Jung, D.S.; Lee, N.Y.; Song, J.H. In vitro activity of fosfomycin against ciprofloxacin-resistant or extended-spectrum beta-lactamase-producing Escherichia coli isolated from urine and blood. Diagn. Microbiol. Infect. Dis. 2007, 58, 111–115. [Google Scholar] [PubMed]
- Lee, S.Y.; Park, Y.J.; Yu, J.K.; Jung, S.; Kim, Y.; Jeong, S.H.; Arakawa, Y. Prevalence of acquired fosfomycin resistance among extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae clinical isolates in Korea and IS26-composite transposon surrounding fosA3. J. Antimicrob. Chemother. 2012, 67, 2843–2847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.R.; Kim, S.J.; Kim, Y.; Kim, J.; Choi, T.Y.; Kang, J.O.; Wie, S.H.; Ki, M.; Cho, Y.K.; Lim, S.K.; et al. Susceptibility of Escherichia coli from community-acquired urinary tract infection to fosfomycin, nitrofurantoin, and temocillin in Korea. J. Korean Med. Sci. 2014, 29, 1178–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.H.; Jung, S.I.; Chung, H.S.; Yu, H.S.; Hwang, E.C.; Kim, S.O.; Kang, T.W.; Kwon, D.D.; Park, K. Antimicrobial susceptibilities of extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in health care-associated urinary tract infection: Focus on susceptibility to fosfomycin. Int. Urol. Nephrol. 2015, 47, 1059–1066. [Google Scholar] [CrossRef]
- Johnson, J.R.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Castanheira, M. Escherichia coli sequence type ST131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin. Infect. Dis. 2010, 51, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.Y.; Choi, H.J.; Choi, J.Y.; Song, M.; Song, Y.; Kim, S.W.; Chang, H.H.; Jung, S.I.; Kim, Y.S.; Ki, H.K.; et al. Dissemination of ST131 and ST393 community-onset, ciprofloxacin-resistant Escherichia coli clones causing urinary tract infections in Korea. J. Infect. 2010, 60, 146–153. [Google Scholar] [CrossRef]
- Colpan, A.; Johnston, B.; Porter, S.; Clabots, C.; Anway, R.; Thao, L.; Kuskowski, M.A.; Tchesnokova, V.; Sokurenko, E.V.; Johnson, J.R. VICTORY (Veterans Influence of Clonal Types on Resistance: Year 2011) Investigators. Escherichia coli sequence type 131 (ST131) subclone H30 as an emergent multidrug-resistant pathogen among US veterans. Clin. Infect. Dis. 2013, 57, 1256–1265. [Google Scholar] [CrossRef] [Green Version]
- Silver, L.L. Fosfomycin: Mechanism and resistance. Cold Spring Harb. Perspect. Med. 2017, 7, a025262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Aguilar, M.; Cantón, R. New microbiological aspects of fosfomycin. Revista Española de Quimioterapia 2019, 32 (Suppl 1), 8–18. [Google Scholar]
- Nilsson, A.I.; Berg, O.G.; Aspevall, O.; Kahlmeter, G.; Andersson, D.I. Biological costs and mechanisms of fosfomycin resistance in Escherichia coli. Antimicrob. Agents Chemother. 2003, 47, 2850–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karageorgopoulos, D.E.; Wang, R.; Yu, X.H.; Falagas, M.E. Fosfomycin: Evaluation of the published evidence on the emergence of antimicrobial resistance in Gram-negative pathogens. J. Antimicrob. Chemother. 2012, 67, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: Twenty-seventh Informational Supplement M100-S28; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Wirth, T.; Falush, D.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; Achtman, M. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.Y.; Yang, Y.; Han, H.; Betzi, S.; Olesen, S.H.; Marsilio, F.; Schönbrunn, E. Functional consequence of covalent reaction of phosphoenolpyruvate with UDP-N-acetylglucosamine 1-carboxyvinyltransferase (MurA). J. Biol. Chem. 2012, 287, 12657–12667. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Huang, X.; Deng, Y.; He, L.; Yang, T.; Zeng, Z.; Chen, Z.; Liu, J.H. Dissemination of the fosfomycin resistance gene fosA3 with CTX-M beta-lactamase genes and rmtB carried on IncFII plasmids among Escherichia coli isolates from pets in China. Antimicrob. Agents Chemother. 2012, 56, 2135–2138. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lim, Y.M.; Rheem, I.; Lee, Y.; Lee, J.C.; Seol, S.Y.; Lee, Y.C.; Cho, D.T. CTX-M and SHV-12 beta-lactamases are the most common extended-spectrum enzymes in clinical isolates of Escherichia coli and Klebsiella pneumoniae collected from 3 university hospitals within Korea. FEMS Microbiol. Lett. 2005, 245, 93–98. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antimicrobial Agents | Number of Resistant Isolates (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total (n = 283) | Mode of Acquisition | Site of Infection c | Facility e | |||||||||
| Community-onset (n = 150) | Hospital-onset (n = 102) | LTCF b-onset (n = 31) | P | UTI d (n = 129) | Non-UTI (n = 154) | P | SMC (n = 114) | SCH (n = 107) | KUAH (n = 62) | P | ||
| Fosfomycin | 19 (6.7) | 9 (6.0) | 7 (6.9) | 3 (10.0) | 0.384 | 5 (3.9) | 14 (9.1) | 0.042 | 4 (3.5) | 9 (8.4) | 6 (9.7) | 0.054 |
| Ciprofloxacin | 181 (64.0) | 72 (48.0) | 70 (68.6) | 22 (71.0) | 0.285 | 70 (72.9) | 111 (59.4) | 0.023 | 67 (58.8) | 62 (57.9) | 52 (83.9) | 0.008 |
| Cefepime | 117 (41.3) | 48 (32.0) | 55 (53.9) | 14 (45.2) | 0.008 | 45 (34.9) | 72 (46.8) | 0.115 | 45 (39.5) | 40 (37.4) | 32 (51.6) | 0.022 |
| Cefixime | 131 (46.3) | 58 (38.7) | 57 (55.9) | 16 (51.6) | 0.033 | 54 (48.9) | 77 (50.0) | 0.363 | 48 (42.1) | 45 (42.1) | 38 (61.3) | 0.062 |
| P/T a | 88 (31.1) | 38 (25.4) | 42 (41.2) | 8 (25.8) | 0.025 | 42 (32.6) | 46 (29.9) | 0.813 | 49 (43.0) | 34 (31.8) | 5 (8.1) | <0.001 |
| Amikacin | 6 (2.1) | 0 | 4 (3.9) | 2 (6.5) | 0.017 | 4 (3.1) | 2 (1.3) | 0.534 | 5 (4.4) | 0 | 1 (1.6) | 0.116 |
| Ertapenem | 7 (2.5) | 1 (0.7) | 6 (5.9) | 0 | 0.012 | 4 (3.1) | 3 (1.9) | 0.880 | 1 (0.9) | 3 (2.8) | 3 (4.8) | 0.244 |
| Colistin | 30 (10.6) | 12 (8.0) | 15 (14.7) | 3 (9.7) | 0.219 | 18 (14.0) | 12 (7.8) | 0.121 | 20 (17.5) | 7 (6.5) | 3 (4.8) | 0.007 |
| Tigecycline | 3 (1.1) | 0 | 2 (2.0) | 1 (3.2) | 0.075 | 2 (1.5) | 1 (0.6) | 0.593 | 0 | 0 | 2 (3.2) | 0.117 |
| Isolate No. | CC a | ST a | Allele no. b | Specimen | Site of Infection c | Mode of Acquisition d | Amino Acid Alterations | |||
|---|---|---|---|---|---|---|---|---|---|---|
| fos | GlpT | UhpT | MurA | |||||||
| S020 | CC131 | ST131 | 53-40-47-13-36-28-29 | Blood | IAI | Hospital | ||||
| S074 | ST131 | 53-40-47-13-36-28-29 | Blood | UTI | Hospital | D220N | ||||
| C072 | ST131-slv1 | 53-40-193-13-36-28-29 | Blood | Cholangitis | Community | |||||
| C073 | ST131-slv2 | 53-40-47-200-36-28-29 | Blood | UTI | LTCF | |||||
| A011 | ST131-dlv1 | 53-35-47-13-36-5-29 | Blood | IAI | Hospital | |||||
| C025 | CC14 | ST1193 | 14-14-10-200-17-7-10 | Urine | UTI | Community | ||||
| C036 | ST1193 | 14-14-10-200-17-7-10 | Blood | Cholangitis | Community | G168R | ||||
| A049 | ST1193 | 14-14-10-200-17-7-10 | Blood | UTI | Community | M136K | ||||
| S019 | ST1193 | 14-14-10-200-17-7-10 | Blood | Prostatitis | LTCF | |||||
| C078 | ST1193-slv | 14-40-10-200-17-7-10 | Blood | UTI | Community | |||||
| C106 | CC69 | ST106 | 21-38-27-6-5-8-4 | Blood | UTI | Community | ||||
| A004 | ST106-dlv | 21-88-27-6-5-79-4 | Blood | UTI | Community | fosA3 | ||||
| A031 | CC95 | ST1531-slv | 37-35-19-37-17-5-26 | Blood | UTI | Community | A16T | |||
| A041 | ST1531-slv | 37-35-19-37-17-5-26 | Blood | UTI | LTCF | A16T | ||||
| C011 | CC155 | ST58 | 64-4-16-24-8-14 | Blood | Liver abscess | Community | Y60F | |||
| C045 | CC38 | ST38 | 4-26-2-25-5-5-19 | Urine | UTI | Hospital | Ins. of DG139 | |||
| C063 | CC10 | ST10 | 10-11-4-8-8-8-2 | Urine | UTI | Hospital | fosA3 | G168R | P99S | |
| A043 | CC398 | ST398-slv | 64-40-1-1-8-8-6 | Blood | Cholangitis | Hospital | A154T | |||
| S050 | CC95 | ST95-slv | 37-38-34-37-17-11-26 | Blood | NF | Hospital | A16T | |||
| Isolate No. | ST a | ESBL Type b | MIC (mg/L) c | Site of Infection d | Mode of Acquisition | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FOS | CIP | CFM | CFX | P/T | AMK | ETM | COL | TGC | |||||
| C093 | ST216 | - | 4 | 0.06 | 0.06 | 0.25 | 16/4 | 4 | 0.06 | 1 | 2 | Cholangitis | Community |
| S088 | ST144-slv | - | 8 | 0.06 | 0.06 | 0.06 | 1/4 | 4 | 0.06 | 0.5 | 1 | Cholangitis | Community |
| A038 | ST131 | CTX-M-15 | 8 | >64 | >64 | >64 | 16/4 | 4 | 0.06 | 1 | 4 | UTI | Community |
| C046 | ST1193 | CTX-M-14 | 16 | >64 | >64 | >64 | 4/4 | 4 | 0.06 | 1 | 1 | UTI | Community |
| Control Strains | |||||||||||||
| E. coli ATCC 25922 | 0.5 | 0.06 | 0.06 | 0.5 | 2/4 | 1 | 0.06 | 0.5 | 0.06 | ||||
| P. aeruginosa ATCC 27853 | 2 | 0.25 | 2 | NAe | 4/4 | 1 | 0.5 | 0.5 | NA | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seok, H.; Choi, J.Y.; Wi, Y.M.; Park, D.W.; Peck, K.R.; Ko, K.S. Fosfomycin Resistance in Escherichia coli Isolates from South Korea and in vitro Activity of Fosfomycin Alone and in Combination with Other Antibiotics. Antibiotics 2020, 9, 112. https://doi.org/10.3390/antibiotics9030112
Seok H, Choi JY, Wi YM, Park DW, Peck KR, Ko KS. Fosfomycin Resistance in Escherichia coli Isolates from South Korea and in vitro Activity of Fosfomycin Alone and in Combination with Other Antibiotics. Antibiotics. 2020; 9(3):112. https://doi.org/10.3390/antibiotics9030112
Chicago/Turabian StyleSeok, Hyeri, Ji Young Choi, Yu Mi Wi, Dae Won Park, Kyong Ran Peck, and Kwan Soo Ko. 2020. "Fosfomycin Resistance in Escherichia coli Isolates from South Korea and in vitro Activity of Fosfomycin Alone and in Combination with Other Antibiotics" Antibiotics 9, no. 3: 112. https://doi.org/10.3390/antibiotics9030112
APA StyleSeok, H., Choi, J. Y., Wi, Y. M., Park, D. W., Peck, K. R., & Ko, K. S. (2020). Fosfomycin Resistance in Escherichia coli Isolates from South Korea and in vitro Activity of Fosfomycin Alone and in Combination with Other Antibiotics. Antibiotics, 9(3), 112. https://doi.org/10.3390/antibiotics9030112