Bioengineered Nisin Derivative M17Q Has Enhanced Activity against Staphylococcus epidermidis


Abstract
:1. Introduction
2. Results
2.1. Agar Based Deferred Antagonism and Well Diffusion Assays
2.2. Agar Well Diffusion Assays, Minimum Inhibitory Concentration Assays and Kill Curve
2.3. Inhibition of Biofilm Formation on Plastic
2.4. Biofilm Inhibition on Stainless Steel by M17Q
2.5. Biofilm Assay on Various Catheter Materials
2.6. Testing the Efficacy of Nisin A and M17Q in a Simulated Wound Fluid
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Agar Based Deferred Antagonism Assays
4.3. Purification of Nisin A and The Derivatives M21V and M17Q
4.4. Agar Well Diffusion Assays
4.5. Minimum Inhibitory Concentration (MIC) Assays
4.6. Kill Curve Assay
4.7. Biofilm Assays with Plastic Microtiter Plates
4.8. Biofilm Assays on Stainless Steel
4.9. Biofilm Assay on Various Catheter Materials
4.10. Testing the Efficacy of Nisin A and M17Q in a Simulated Wound Fluid
4.11. Reproducibility and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.Y.H.; Monk, I.R.; Gonçalves da Silva, A.; Seemann, T.; Chua, K.Y.L.; Kearns, A.; Hill, R.; Woodford, N.; Bartels, M.D.; Strommenger, B.; et al. Global Spread of Three Multidrug-Resistant Lineages of Staphylococcus epidermidis. Nat. Microbiol. 2018, 3, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Liu, P.-F.; Liu, Y.-T.; Kuo, S.; Zhang, X.-Q.; Schooley, R.T.; Rohde, H.; Gallo, R.L.; Huang, C.-M. Nasal Commensal Staphylococcus epidermidis Counteracts Influenza Virus. Sci. Rep. 2016, 6, 27870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Center for Disease Prevention and Control. Rapid Risk Assessment: Multidrug-Resistant Staphylococcus epidermidis. 2018. Available online: https://ecdc.europa.eu/sites/portal/files/documents/15-10-2018-RRA-Staphylococcus epidermidis%2C Antimicrobial resistance-World_ZCS9CS.pdf (accessed on 11 March 2020).
- Christensen, G.D.; Bisno, A.L.; Parisi, J.T.; McLaughlin, B.; Hester, M.G.; Luther, R.W. Nosocomial Septicemia Due to Multiply Antibiotic-Resistant Staphylococcus epidermidis. Ann. Intern. Med. 1982, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Büttner, H.; Mack, D.; Rohde, H. Structural Basis of Staphylococcus epidermidis Biofilm Formation: Mechanisms and Molecular Interactions. Front. Cell. Infect. Microbiol. 2015, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Borgwardt, L.; Høiby, N.; Wu, H.; Sørensen, T.S.; Borgwardt, A. Prosthesis Infections after Orthopedic Joint Replacement: The Possible Role of Bacterial Biofilms. Orthop. Rev. (Pavia) 2013, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- van der Meer, J.R.; Polman, J.; Beerthuyzen, M.M.; Siezen, R.J.; Kuipers, O.P.; De Vos, W.M. Characterization of the Lactococcus lactis Nisin a Operon Genes NisP, Encoding a Subtilisin-like Serine Protease Involved in Precursor Processing, and NisR, Encoding a Regulatory Protein Involved in Nisin Biosynthesis. J. Bacteriol. 1993, 175, 2578–2588. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineering Lantibiotics for Therapeutic Success. Front. Microbiol. 2015, 6, 1363. [Google Scholar] [CrossRef] [Green Version]
- Breukink, E.; Wiedemann, I.; van Kraaij, C.; Kuipers, O.P.; Sahl, H.G.; de Kruijff, B. Use of the Cell Wall Precursor Lipid II by a Pore-Forming Peptide Antibiotic. Science 1999, 286, 2361–2364. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Opinion of the Scientific Panel on Food Additives, Flavourings, Processing Aids and Materials in Contact with Food on a Request from the Commission Related to the Use of Nisin (E 234) as a Food Additive. 2006. Available online: http://www.efsa.eu.int/science/afc/afc_opinions/catindex_en.html (accessed on 11 March 2020).
- Campion, A.; Casey, P.G.; Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. In Vivo Activity of Nisin A and Nisin V against Listeria monocytogenes in Mice. BMC Microbiol. 2013, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Bower, C.; Parker, J.; Higgins, A.; Oest, M.; Wilson, J.; Valentine, B.; Bothwell, M.; McGuire, J. Protein Antimicrobial Barriers to Bacterial Adhesion: In Vitro and in Vivo Evaluation of Nisin-Treated Implantable Materials. Colloids Surf. B Biointerfaces 2002, 25, 81–90. [Google Scholar] [CrossRef]
- Piper, C.; Casey, P.G.; Hill, C.; Cotter, P.D.; Ross, R.P. The Lantibiotic lacticin 3147 Prevents Systemic Spread of Staphylococcus aureus in a Murine Infection Model. Int. J. Microbiol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.N.; O’Connor, P.M.; Rea, M.C.; O’Sullivan, O.; Walsh, C.J.; Healy, B.; Mathur, H.; Field, D.; Hill, C.; Ross, R.P. Nisin J, a Novel Natural Nisin Variant, Is Produced by Staphylococcus capitis Sourced from the Human Skin Microbiota. J. Bacteriol. 2020, 202, e00639-19. [Google Scholar] [CrossRef] [Green Version]
- Wirawan, R.E.; Klesse, N.A.; Jack, R.W.; Tagg, J.R. Molecular and Genetic Characterization of a Novel Nisin Variant Produced by Streptococcus uberis. Appl. Environ. Microbiol. 2006, 72, 1148–1156. [Google Scholar] [CrossRef] [Green Version]
- Mulders, J.W.M.; Boerrigter, I.J.; Rollema, H.S.; Siezen, R.J.; Vos, W.M. Identification and Characterization of the Lantibiotic Nisin Z, a Natural Nisin Variant. Eur. J. Biochem. 1991, 201, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Zendo, T.; Fukao, M.; Ueda, K.; Higuchi, T.; Nakayama, J.; Sonomoto, K. Identification of the Lantibiotic Nisin Q, a New Natural Nisin Variant Produced by Lactococcus lactis 61-14 Isolated from a River in Japan. Biosci. Biotechnol. Biochem. 2003, 67, 1616–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.; Quigley, L.; O’Connor, P.M.; Rea, M.C.; Daly, K.; Cotter, P.D.; Hill, C.; Ross, R.P. Studies with Bioengineered Nisin Peptides Highlight the Broad-Spectrum Potency of Nisin V. Microb. Biotechnol. 2010, 3, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Connor, P.M.O.; Cotter, P.D.; Hill, C.; Ross, R.P. The Generation of Nisin Variants with Enhanced Activity against Specific Gram-Positive Pathogens. Mol. Microbiol. 2008, 69, 218–230. [Google Scholar] [CrossRef]
- Field, D.; Gaudin, N.; Lyons, F.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. A Bioengineered Nisin Derivative to Control Biofilms of Staphylococcus pseudintermedius. PLoS ONE 2015, 10, e0119684. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.K.; Draper, L.A.; Hazelhoff, P.-J.; Cotter, P.D.; Ross, R.P.; Hill, C. A Bioengineered Nisin Derivative, M21A, in Combination with Food Grade Additives Eradicates Biofilms of Listeria monocytogenes. Front. Microbiol. 2016, 7, 1939. [Google Scholar] [CrossRef] [Green Version]
- Davison, W.M.; Pitts, B.; Stewart, P.S. Spatial and Temporal Patterns of Biocide Action against Staphylococcus epidermidis Biofilms. Antimicrob. Agents Chemother. 2010, 54, 2920–2927. [Google Scholar] [CrossRef] [Green Version]
- Ghiselli, R.; Giacometti, A.; Cirioni, O.; Dell’Acqua, G.; Mocchegiani, F.; Orlando, F.; D’Amato, G.; Rocchi, M.; Scalise, G.; Saba, V. RNAIII-Inhibiting Peptide And/Or Nisin Inhibit Experimental Vascular Graft Infection with Methicillin-Susceptible and Methicillin-Resistant Staphylococcus epidermidis. Eur. J. Vasc. Endovasc. Surg. 2004, 27, 603–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healy, B.; Field, D.; O’Connor, P.M.; Hill, C.; Cotter, P.D.; Ross, R.P. Intensive Mutagenesis of the Nisin Hinge Leads to the Rational Design of Enhanced Derivatives. PLoS ONE 2013, 8, e79563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouse, S.; Field, D.; Daly, K.M.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineered Nisin Derivatives with Enhanced Activity in Complex Matrices. Microb. Biotechnol. 2012, 5, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Silva, J.; Jekhmane, S.; Paioni, A.L.; Gawarecka, K.; Baldus, M.; Swiezewska, E.; Breukink, E.; Weingarth, M. High-Resolution NMR Studies of Antibiotics in Cellular Membranes. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, D.; Daly, K.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Efficacies of Nisin A and Nisin V Semipurified Preparations Alone and in Combination with Plant Essential Oils for Controlling Listeria monocytogenes. Appl. Environ. Microbiol. 2015, 81, 2762–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.J.; Fatma, T.; Rattan, A. Detection of biofilm formation among the clinical isolates of Staphylococci: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin Microbiota-Host Interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Field, D.; Cotter, P.D.; Ross, R.P.; Hill, C. Bioengineering of the Model Lantibiotic Nisin. Bioengineered 2015, 6, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Dunne, W.M.; Mason, E.O.; Kaplan, S.L., Jr.; Kaplan, S.L. Diffusion of Rifampin and Vancomycin through a Staphylococcus epidermidis Biofilm. Antimicrob. Agents Chemother. 1993, 37, 2522–2526. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, K.K.; Goldmann, D.A.; Pier, G.B. Use of Confocal Microscopy to Analyze the Rate of Vancomycin Penetration through Staphylococcus aureus Biofilms. Antimicrob. Agents Chemother. 2005, 49, 2467–2473. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of Antibiotic Penetration, Oxygen Limitation, and Low Metabolic Activity to Tolerance of Pseudomonas aeruginosa Biofilms to Ciprofloxacin and Tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Zhang, J.; Hu, Y.; Chi, Q.; Mortensen, N.P.; Qu, D.; Molin, S.; Ulstrup, J. Organic Compounds Inhibiting S. epidermidis Adhesion and Biofilm Formation. Ultramicroscopy 2009, 109, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Zendo, T.; Sugimoto, S.; Iwase, T.; Tajima, A.; Yamada, S.; Sonomoto, K. and Mizunoe, Y. Effects of Bacteriocins On Methicillin-Resistant Staphylococcus aureus Biofilm. Antimicrob. Agents Chemother. Am. Soc. Microbiol. J. 2013, 57, 5572–5579. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Connor, R.O.; Cotter, P.D.; Ross, R.P.; Hill, C. In Vitro Activities of Nisin and Nisin Derivatives Alone and in Combination with Antibiotics against Staphylococcus Biofilms. Front. Microbiol. 2016, 7, 508. [Google Scholar] [CrossRef] [Green Version]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Fighting Biofilms with Lantibiotics and Other Groups of Bacteriocins. NPJ Biofilms Microbiomes 2018, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Minei, C.C.; Gomes, B.C.; Ratti, R.P.; D’angelis, C.E.M.; De Martinis, E.C.P. Influence of Peroxyacetic Acid and Nisin And Coculture with Enterococcus faecium On Listeria monocytogenes Biofilm Formation. J. Food Protection. 2008, 71, 634–638. [Google Scholar] [CrossRef]
- Natsis, N.E. and Cohen, P.R. Coagulase-Negative Staphylococcus Skin And Soft Tissue Infections. Am. J. Clin. Dermatol. 2018, 19, 671–677. [Google Scholar] [CrossRef]
- van Staden, A.D.P.; Heunis, T.; Smith, C.; Deane, S.; Dicks, L.M.T. Efficacy of Lantibiotic Treatment of Staphylococcus aureus—Induced Skin Infections, Monitored by In Vivo Bioluminescent Imaging. Antimicrob. Agents Chemother. 2016, 60, 3948–3955. [Google Scholar] [CrossRef] [Green Version]
- Mouritzen, M.V.; Andrea, A.; Qvist, K.; Poulsen, S.S.; Jenssen, H. Immunomodulatory Potential Of Nisin A With Application In Wound Healing. Wound Repair Regen. 2019, 27, 650–660. [Google Scholar] [CrossRef]
- Field, D.; Begley, M.; O’Connor, P.M.; Daly, K.M.; Hugenholtz, F.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineered Nisin A Derivatives with Enhanced Activity against Both Gram Positive and Gram Negative Pathogens. PLoS ONE 2012, 7, e46884. [Google Scholar] [CrossRef] [Green Version]
- Molloy, E.M.; Field, D.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Saturation Mutagenesis of Lysine 12 Leads to the Identification of Derivatives of Nisin A with Enhanced Antimicrobial Activity. PLoS ONE 2013, 8, e58530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, C.; Freeman, R.; Percival, S.L. In Vitro Study of Sustained Antimicrobial Activity of a New Silver Alginate Dressing. J. Am. Col. Certif. Wound Spec. 2009, 1, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]


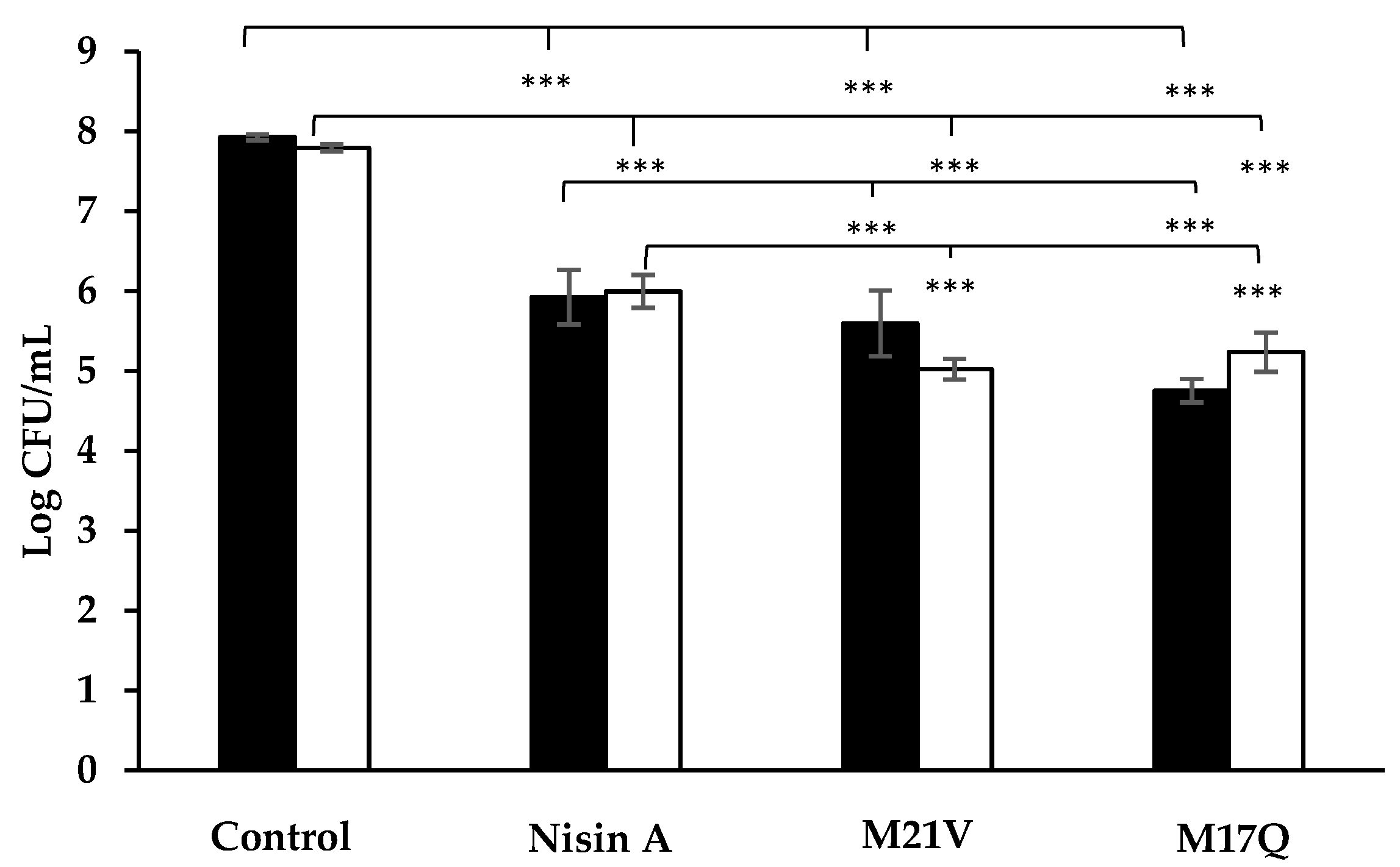
 ) and S. epidermidis 53 (
) and S. epidermidis 53 (  ) after 1 h incubation in tryptone soya broth supplemented with 3.75μM nisin A, M21V or M17Q peptide. (*** = p < 0.001).
) and S. epidermidis 53 ( ) after 1 h incubation in tryptone soya broth supplemented with 3.75μM nisin A, M21V or M17Q peptide. (*** = p < 0.001).
) after 1 h incubation in tryptone soya broth supplemented with 3.75μM nisin A, M21V or M17Q peptide. (*** = p < 0.001).
) and S. epidermidis 53 ( ) after 1 h incubation in tryptone soya broth supplemented with 3.75μM nisin A, M21V or M17Q peptide. (*** = p < 0.001).
 ,
,  ), 3.75 μM, (
), 3.75 μM, (  ,
,  ) and 7.5 μM, (
) and 7.5 μM, (  ,
,  ) nisin A and derivative peptides, M21V and M17Q. Control shows the amount of biofilm produced in the absence of nisin, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-”denotes no statistical significance.
, ), 3.75 μM, ( , ) and 7.5 μM, ( , ) nisin A and derivative peptides, M21V and M17Q. Control shows the amount of biofilm produced in the absence of nisin, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-”denotes no statistical significance.
) nisin A and derivative peptides, M21V and M17Q. Control shows the amount of biofilm produced in the absence of nisin, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-”denotes no statistical significance.
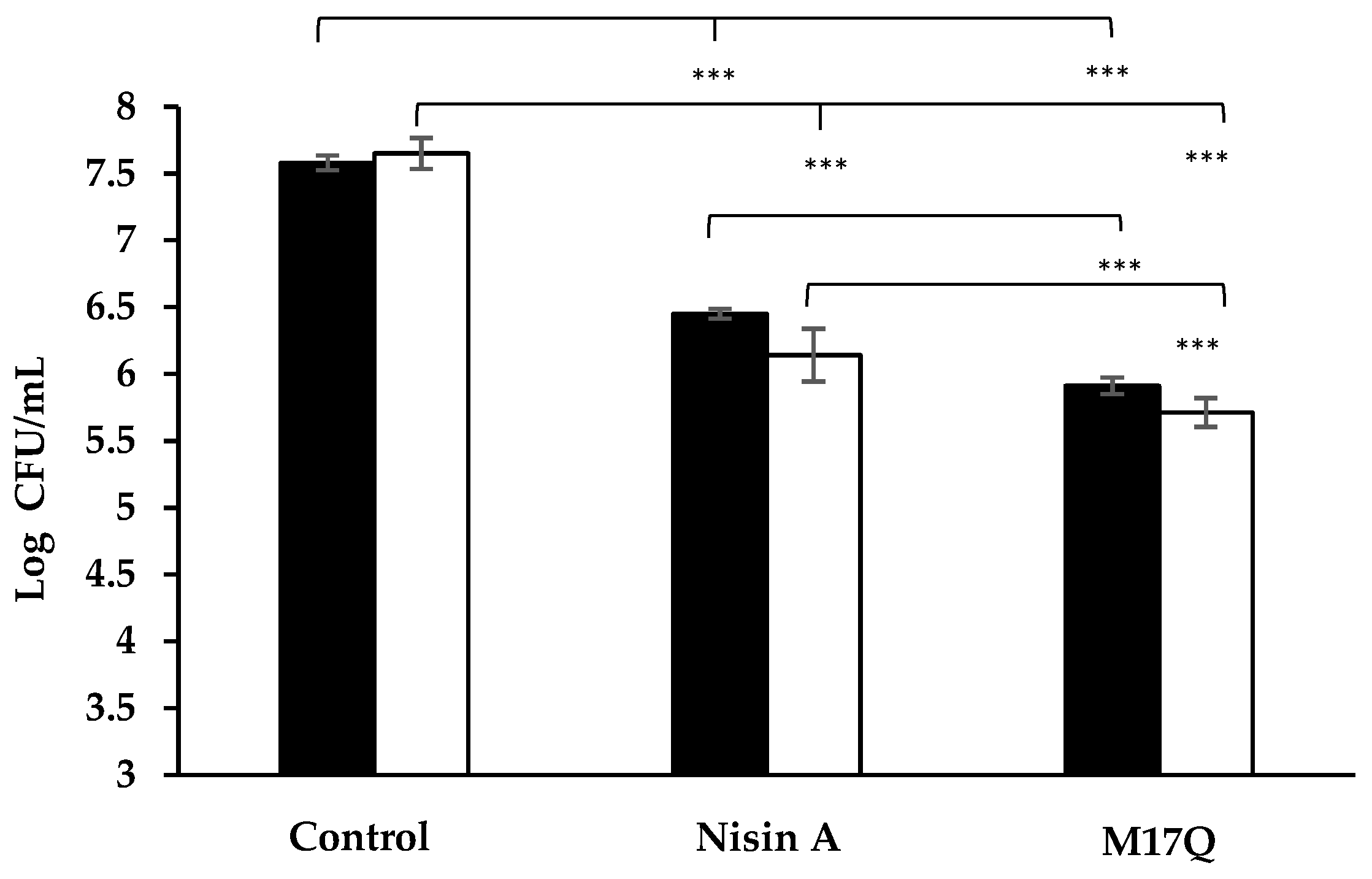
, ), 3.75 μM, ( , ) and 7.5 μM, ( , ) nisin A and derivative peptides, M21V and M17Q. Control shows the amount of biofilm produced in the absence of nisin, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-”denotes no statistical significance. , ), the presence of nisin A at 3.75 μM, ( ,
, ), the presence of nisin A at 3.75 μM, ( ,  ) and derivative peptide M17Q at 3.75 μM, ( ,
) and derivative peptide M17Q at 3.75 μM, ( ,  ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and—shows no statistical significance.
, ), the presence of nisin A at 3.75 μM, ( , ) and derivative peptide M17Q at 3.75 μM, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and—shows no statistical significance.
). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and—shows no statistical significance.
, ), the presence of nisin A at 3.75 μM, ( , ) and derivative peptide M17Q at 3.75 μM, ( , ). *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and—shows no statistical significance.
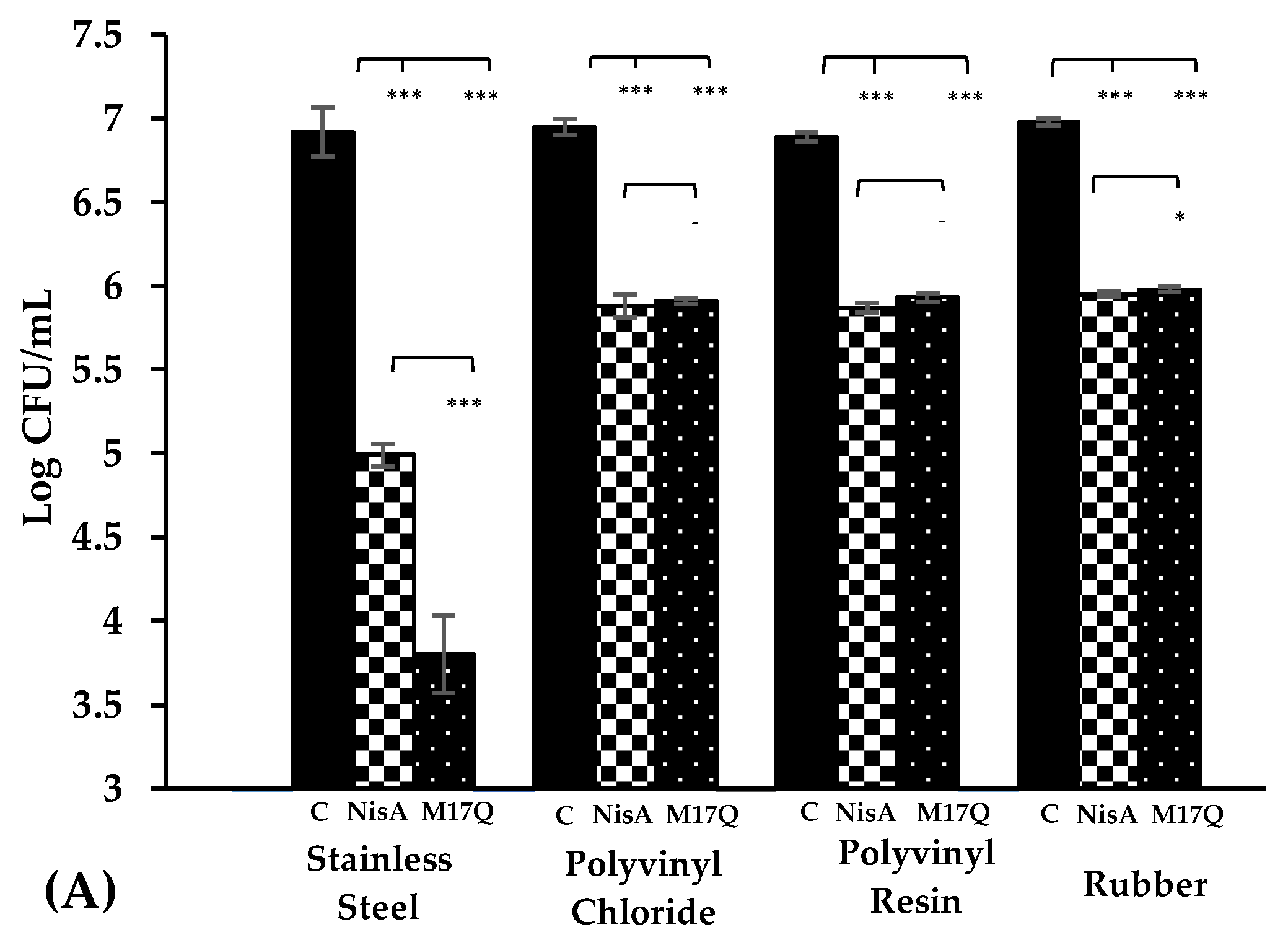
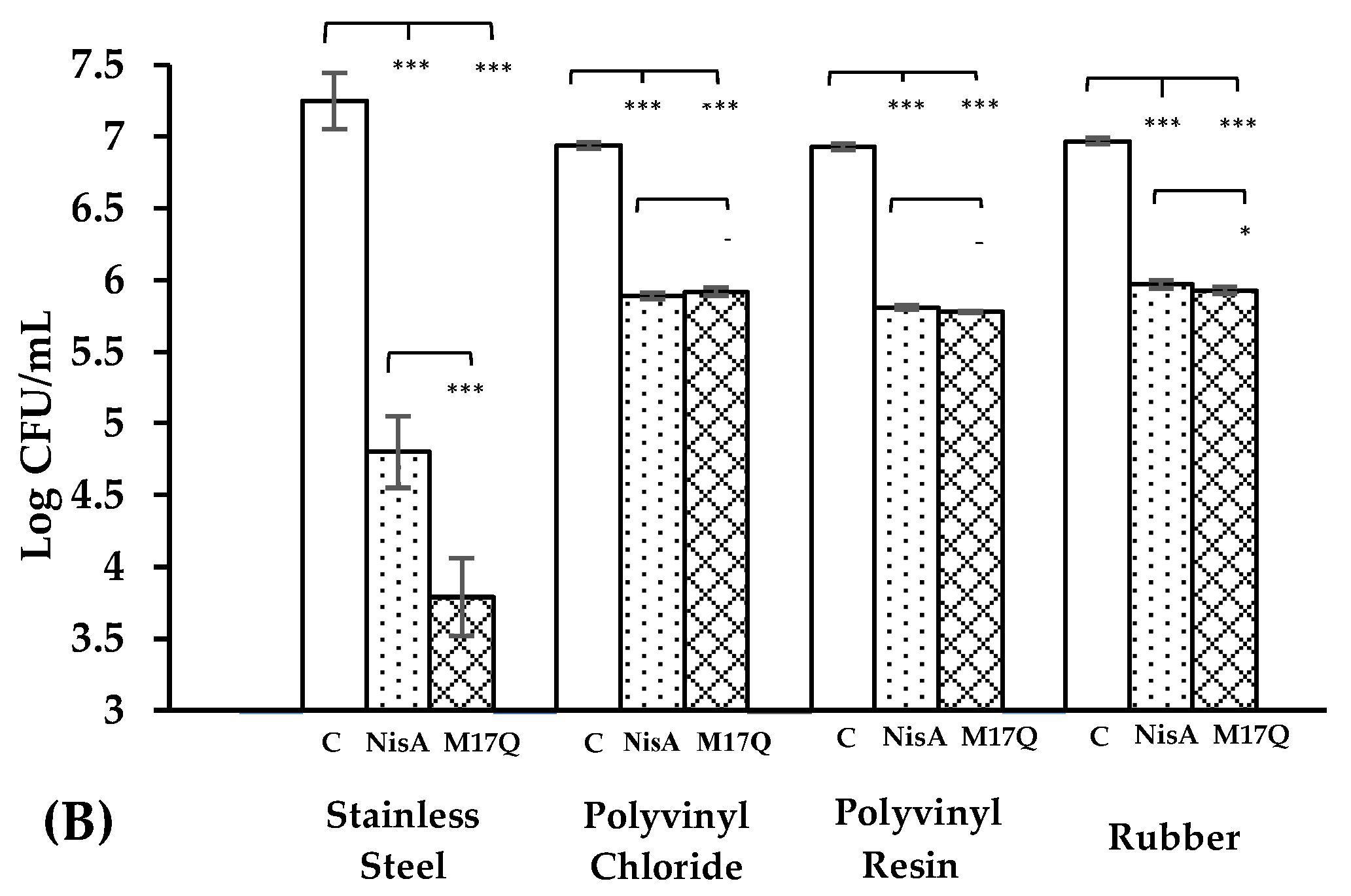
 ) and S. epidermidis 53 ( ) were incubated for 1hr at 37 °C with nisin A and M17Q at a concentration of 3.75 μM. *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-” denotes no statistical significance.
) and S. epidermidis 53 ( ) were incubated for 1hr at 37 °C with nisin A and M17Q at a concentration of 3.75 μM. *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-” denotes no statistical significance.
) and S. epidermidis 53 ( ) were incubated for 1hr at 37 °C with nisin A and M17Q at a concentration of 3.75 μM. *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-” denotes no statistical significance.
) and S. epidermidis 53 ( ) were incubated for 1hr at 37 °C with nisin A and M17Q at a concentration of 3.75 μM. *** denotes p < 0.001, ** denotes p < 0.005, * denotes p < 0.05 and “-” denotes no statistical significance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. epidermidis Strains | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 17 | 22 | 26 | 28 | 53 | 70 | 78 | 80 | 89 | 90 | 92 | 100 | 101 | 102 | 103 | 104 | 106 | |
| WT Nisin | 11 | 14.9 | 12.3 | 16.3 | 11.2 | 11 | 9.8 | 10.8 | 11.7 | 12 | 11 | 11 | 11 | 12.3 | 12.3 | 10.7 | 13.3 | 13.6 |
| Bioengineered L. lactis strains | ||||||||||||||||||
| M21V | 9.5 | 15.9 | 13.7 | 16 | 12.7 | 12.6 | 13 | 12 | 13 | 13 | 12.7 | 12 | 13.1 | 13.7 | 13.3 | 13.6 | 14 | 14 |
| M17Q | 14.3 | 21 | 19.4 | 21 | 19.9 | 15 | 13.1 | 17.7 | 19.4 | 17.8 | 17 | 16.4 | 15.7 | 18.3 | 16.8 | 15 | 20.3 | 19.7 |
| M21S | 11.3 | 16.6 | 12.8 | 16.7 | 13.6 | 12.8 | 11 | 16.7 | 14.3 | 14.2 | 11.2 | 11 | 12.6 | 15.4 | 13 | 12.7 | 14.7 | 13.7 |
| M21V-H31K | 9 | 15.5 | 12.8 | 15.7 | 13 | 12.4 | 9 | 12 | 14 | 11 | 12 | 11 | 11 | 12.1 | 13.2 | 12.5 | 13 | 12.7 |
| M21K | 10 | 13 | 9.8 | 15 | 12 | 10 | 11.3 | 12 | 10 | 14.5 | 11.7 | 9.7 | 11.8 | 15 | 12 | 12.5 | 12.7 | 12.8 |
| VGA | 15.7 | 18.2 | 16.8 | 18 | 14.5 | 17 | 10.2 | 15.7 | 15.2 | 18 | 13.8 | 18 | 15.7 | 15 | 18 | 15 | 19.3 | 20.7 |
| K12A | 10.7 | 13 | 13.4 | 15.9 | 14 | 13.3 | 10.9 | 10 | 15 | 15.2 | 13.3 | 13 | 11.5 | 14 | 15 | 11.9 | 15.3 | 13.7 |
| K12S | 10 | 16 | 12 | 16.7 | 15 | 12 | 8 | 17 | 14 | 13.4 | 12.3 | 11.4 | 13 | 14.5 | 14.6 | 12.5 | 17.7 | 14.7 |
| K12T | 10 | 17.7 | 10.5 | 15.1 | 10 | 10 | 8 | 10 | 9 | 10 | 12 | 9 | 11 | 10 | 12.5 | 12.3 | 13.8 | 11.3 |
| K22T | 10 | 16.5 | 11.2 | 18.2 | 13.6 | 11.5 | 13.6 | 10.8 | 13.3 | 14.6 | 13.3 | 10 | 11.4 | 15 | 17 | 12.7 | 16 | 13.6 |
| VGV | 10.1 | 14.9 | 13.6 | 16.1 | 13.3 | 13.3 | 11 | 9 | 12.4 | 8 | 12 | 15.7 | 11 | 13.7 | 11.2 | 13.5 | 18.5 | 18 |
| VGT | 12.2 | 18 | 16.4 | 18 | 14.2 | 16 | 11 | 11 | 14 | 15.1 | 14.3 | 11.7 | 11.7 | 13.3 | 15.8 | 13 | 19 | 14.6 |
| PIT | 11.7 | 15.7 | 11.3 | 17 | 13 | 13.5 | 11 | 11 | 10 | 16.2 | 11.6 | 10 | 12.7 | 11 | 15.2 | 12.1 | 16.3 | 16.1 |
| PGA | 13 | 19.7 | 14.3 | 19.4 | 19.6 | 14 | 12.7 | 10.6 | 16.4 | 9 | 16.3 | 11.5 | 14.7 | 15 | 15.5 | 14.9 | 18 | 15.7 |
| T2L | 11 | 10 | 9.5 | 12.2 | 10.5 | 10 | 8 | 11.8 | 10 | 12 | 11.8 | 16 | 14.1 | 10.7 | 14.3 | 14 | 14.9 | 15.1 |
| G18Dhb | 12.2 | 11 | 8 | 14.7 | 11 | 10.3 | 8 | 10 | 16.3 | 11.7 | 10 | 10 | 12 | 13 | 12 | 12 | 11.9 | 12.7 |
| SGK | 13.4 | 17 | 13.7 | 18.8 | 15.3 | 13.7 | 10 | 11 | 13.7 | 13.7 | 15 | 13.4 | 14 | 13 | 13.7 | 13.7 | 16 | 16.5 |
| PAQ | 10 | 12.5 | 11.7 | 18 | 12 | 11.8 | 10 | 14 | 9 | 13 | 14 | 12 | 11 | 12.7 | 14.3 | 12.3 | 17 | 15.3 |
| HTK | 11.6 | 16.5 | 10.4 | 17 | 14 | 10 | 10 | 14.3 | 14.5 | 13.7 | 12 | 11.7 | 12.3 | 13 | 16 | 12.3 | 15.9 | 15.9 |
| I4V | 0 | 14.6 | 11.7 | 18 | 15.9 | 11.5 | 9 | 11 | 10.7 | 12.7 | 13 | 12 | 13 | 11.7 | 13.7 | 11 | 15 | 14.7 |
| N2OP | 10 | 15.5 | 11.8 | 18 | 14.5 | 11.3 | 10 | 12.3 | 16.9 | 14.7 | 16 | 13.6 | 11.5 | 15 | 13.7 | 13.5 | 16.9 | 16.4 |
| S29R | 10 | 14 | 10.8 | 15.6 | 12.6 | 11 | 9 | 9 | 10.5 | 14 | 12.7 | 10 | 7 | 9 | 11.4 | 11 | 13.4 | 14 |
| S29E | 10 | 15.4 | 11.3 | 16.7 | 11.7 | 11 | 9 | 11 | 10 | 13.6 | 11.7 | 11 | 11 | 12 | 17 | 16.9 | 12 | 14.2 |
| S29A | 10 | 16 | 11.7 | 17.3 | 14 | 11.4 | 10 | 11 | 11.6 | 14.7 | 12.7 | 12 | 11.9 | 13.6 | 14 | 13 | 15.3 | 14 |
| S29D | 9 | 10.4 | 14 | 13.2 | 11.9 | 9.5 | 11.5 | 11 | 10.9 | 10 | 11.3 | 10.7 | 8 | 11 | 14.3 | 10 | 13 | 12.2 |
| M21A | 12 | 17.4 | 18.3 | 19.3 | 14.4 | 14.7 | 11 | 11.5 | 17.3 | 15.5 | 14 | 16.7 | 15 | 15.5 | 15.2 | 13.2 | 16.7 | 16.7 |
| H31K | 10.5 | 10.9 | 14 | 13 | 14 | 11.3 | 10.1 | 8.4 | 10 | 9.3 | 11.9 | 10.3 | 8 | 12 | 12.5 | 10.4 | 12.7 | 14 |
| AAK | 12.5 | 16 | 13.2 | 15.7 | 11.3 | 11.4 | 9 | 12 | 12.5 | 15 | 13.3 | 13.8 | 12.3 | 13.7 | 16.3 | 12 | 13.7 | 15 |
| AAA | 15 | 19.9 | 21 | 21 | 20 | 14.6 | 15.9 | 17.5 | 16 | 19 | 15.8 | 17 | 16 | 19 | 19.3 | 14 | 20 | 19 |
| Nisin A | M21V | M17Q | |
|---|---|---|---|
| S. epidermidis 28 | 3.75 | 1.875 | 1.875 |
| S. epidermidis 53 | 7.5 | 3.75 | 3.75 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Twomey, E.; Hill, C.; Field, D.; Begley, M. Bioengineered Nisin Derivative M17Q Has Enhanced Activity against Staphylococcus epidermidis. Antibiotics 2020, 9, 305. https://doi.org/10.3390/antibiotics9060305
Twomey E, Hill C, Field D, Begley M. Bioengineered Nisin Derivative M17Q Has Enhanced Activity against Staphylococcus epidermidis. Antibiotics. 2020; 9(6):305. https://doi.org/10.3390/antibiotics9060305
Chicago/Turabian StyleTwomey, Ellen, Colin Hill, Des Field, and Maire Begley. 2020. "Bioengineered Nisin Derivative M17Q Has Enhanced Activity against Staphylococcus epidermidis" Antibiotics 9, no. 6: 305. https://doi.org/10.3390/antibiotics9060305
APA StyleTwomey, E., Hill, C., Field, D., & Begley, M. (2020). Bioengineered Nisin Derivative M17Q Has Enhanced Activity against Staphylococcus epidermidis. Antibiotics, 9(6), 305. https://doi.org/10.3390/antibiotics9060305