Analysis of Gastrointestinal Responses Revealed Both Shared and Specific Targets of Zinc Oxide and Carbadox in Weaned Pigs

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
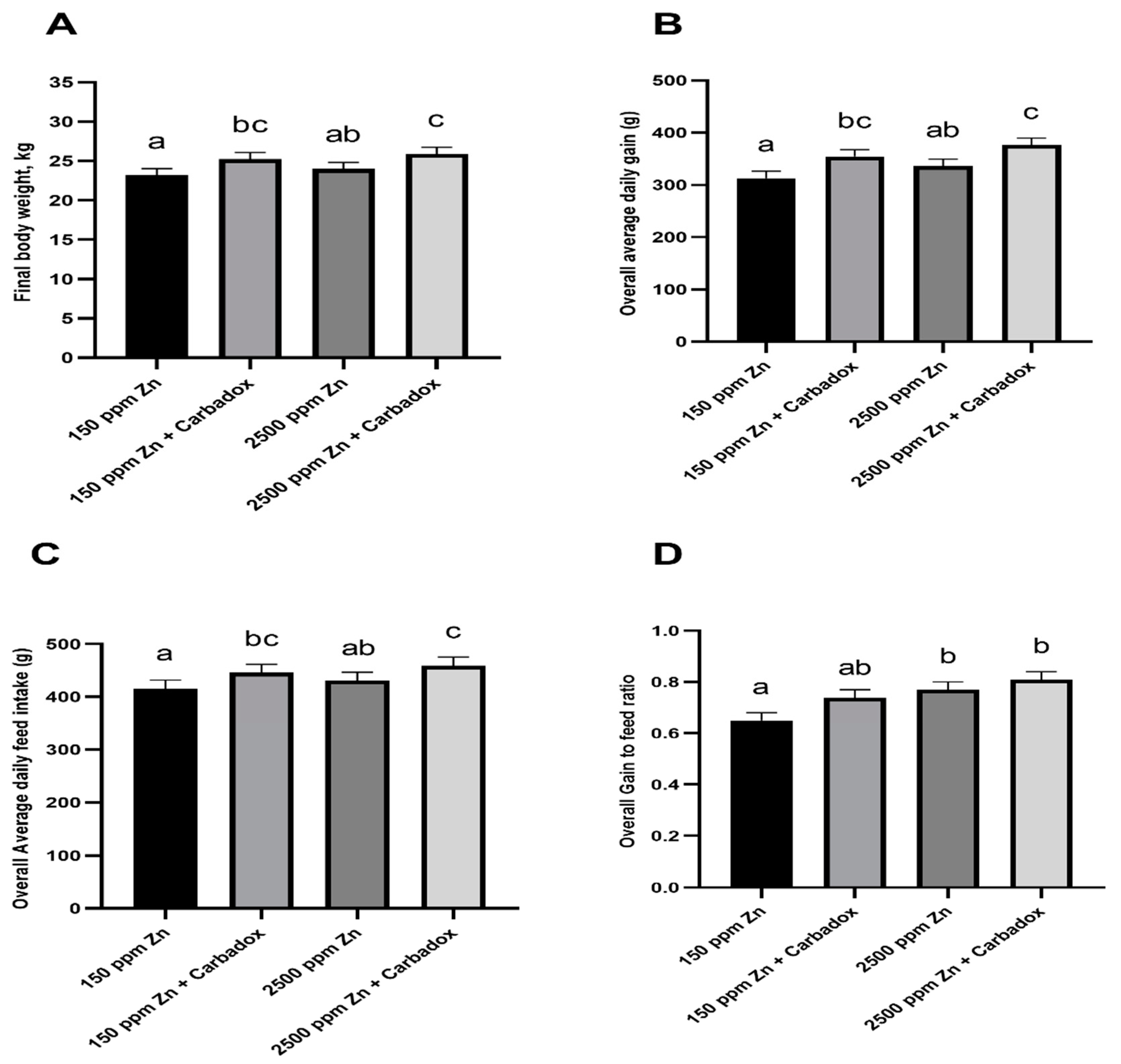
2.1. Effects of Dietary Zn and Carbadox on Growth Performance
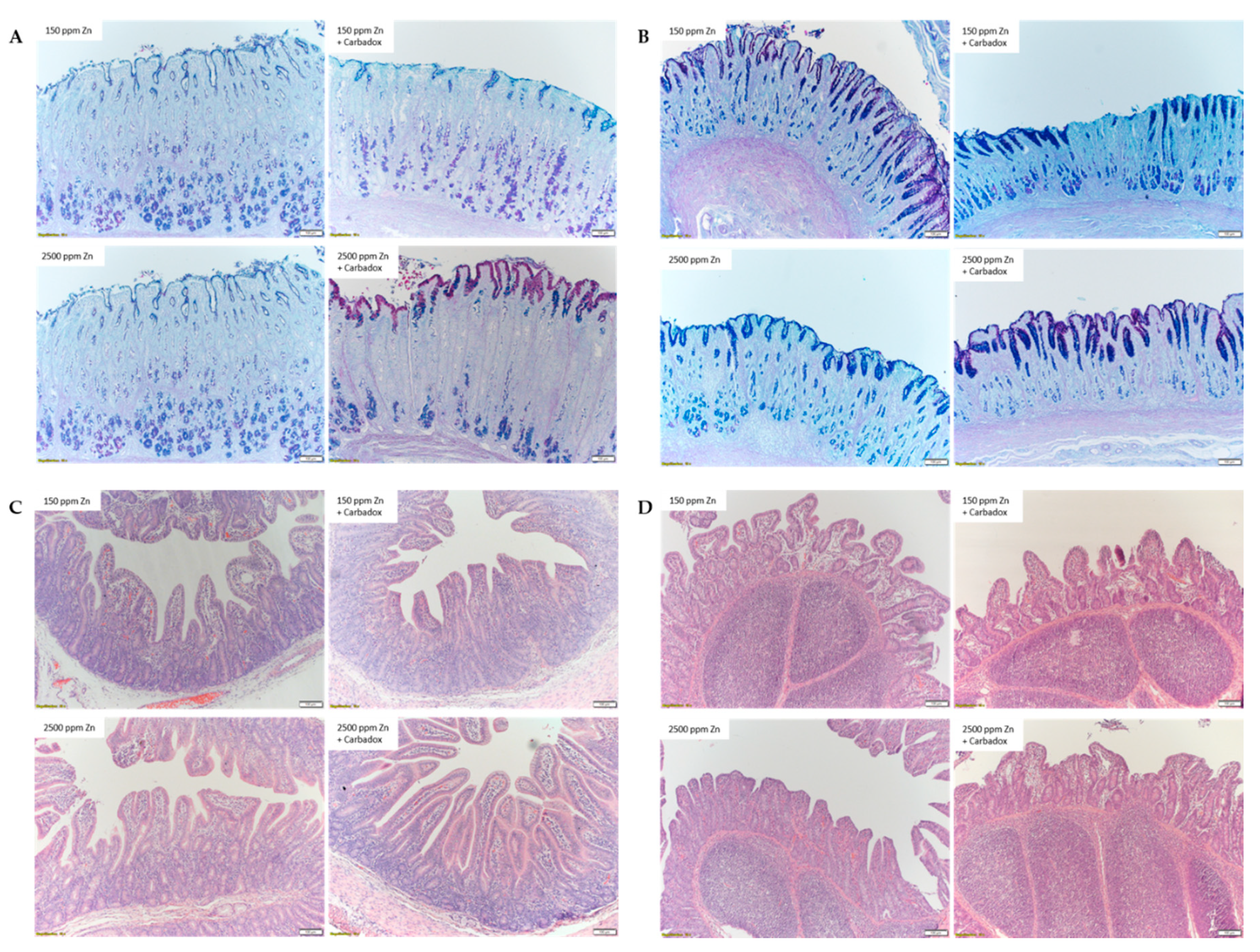
2.2. Effects of Dietary Zn and Carbadox on Gastric Acid Production and GI Morphology
2.3. Effects of Dietary Zn and Carbadox on Ileal Cytokines
2.4. Effects of Dietary Zn and Carbadox on Free Amino Acid (AA), Bile Acid (BA), and Fatty Acid (FA) in Ileal Digesta
2.5. Effects of Dietary Zn and Carbadox on Free AA, BA, and FA in Feces
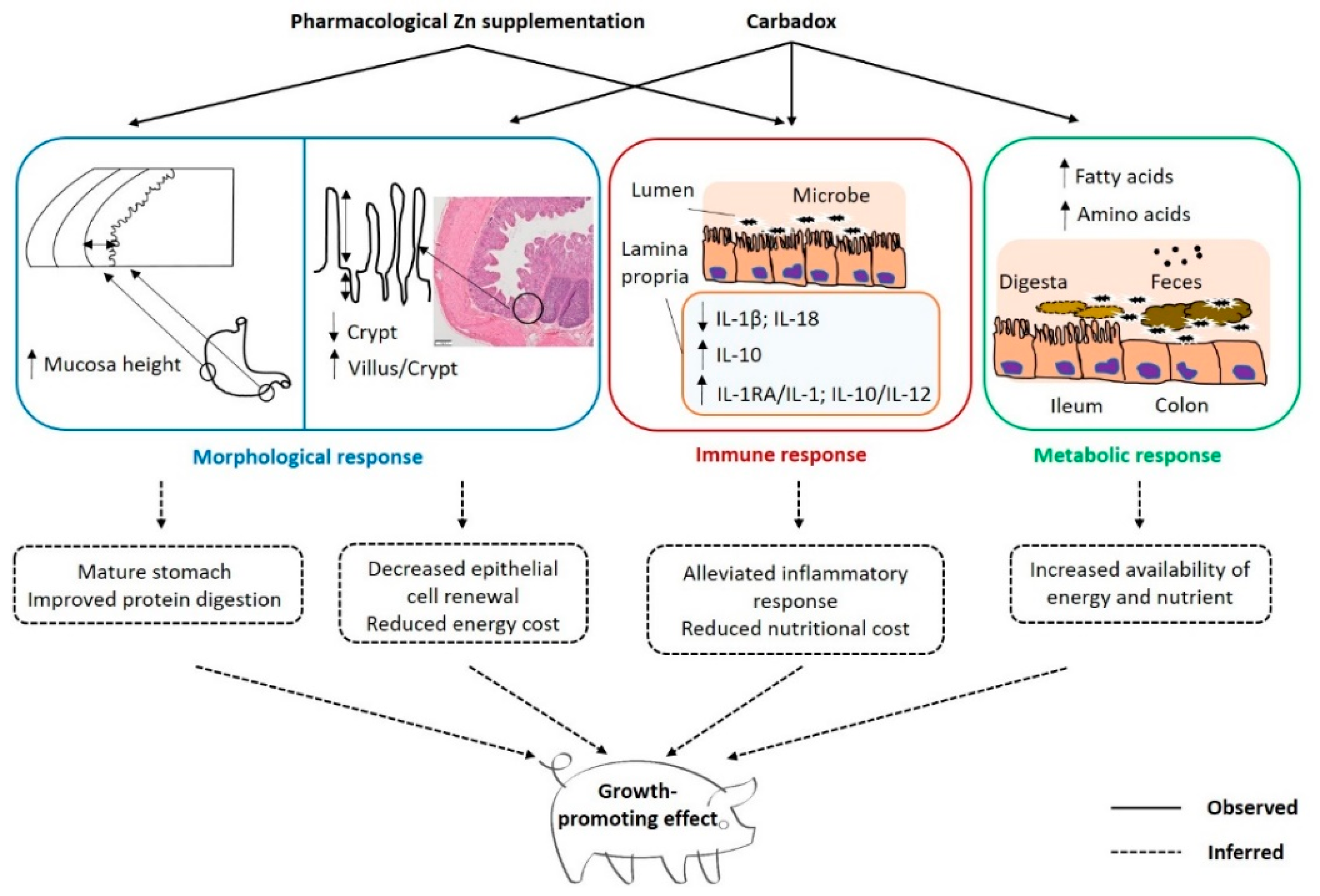
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Data and Sample Collection
4.3. Gastric Acid Titration
4.4. Histological Analysis
4.5. Tissue Cytokine Analysis
4.6. Quantitative Analysis of Free Amino Acids, Fatty Acids, and Bile Acids in Ileal Digesta and Feces
4.7. Statistical Analysis
4.8. Ethical Approval
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cogliani, C.; Goossens, H.; Greko, C. Restricting antimicrobial use in food animals: Lessons from Europe. Microbe 2011, 6, 274–279. [Google Scholar] [CrossRef]
- Vahjen, W.; Pietruszyńska, D.; Starke, I.C.; Zentek, J. High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog. 2015, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Dȩbski, B. Supplementation of pigs diet with zinc and copper as alternative to conventional antimicrobials. Pol. J. Vet. Sci. 2016, 19, 917–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.; Larsen, M.M.; Bak, J. National monitoring study in Denmark finds increased and critical levels of copper and zinc in arable soils fertilized with pig slurry. Environ. Pollut. 2016, 214, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration Veterinary Feed Directive. Fed. Regist. 2015, 80, 31708–31735.
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Gabler, N.K.; Schweer, W.P. Alternatives to in-feed antibiotics for nursery pigs. In Proceedings of the 78th Minnesota Nutrition Conference, University of Minnesota, Mankato, Minnesota, 20–21 September 2018; pp. 191–198. [Google Scholar]
- Poulsen, H.D. Zinc oxide for weanling piglets. Acta Agric. Scand. Sect. A Anim. Sci. 1995, 45, 159–167. [Google Scholar] [CrossRef]
- Sales, J. Effects of pharmacological concentrations of dietary zinc oxide on growth of post-weaning pigs: A meta-analysis. Biol. Trace Elem. Res. 2013, 152, 343–349. [Google Scholar] [CrossRef]
- Yen, J.T.; Nienaber, J.A.; Pond, W.G.; Varel, V.H. Effect of carbadox on growth, fasting metabolism, thyroid function and gastrointestinal tract in young pigs. J. Nutr. 1985, 115, 970–979. [Google Scholar] [CrossRef]
- Thrasher, G.W.; Shively, J.E.; Askelson, C.E.; Babcock, W.E.; Haute, T.; Procedure, E. Effects of feeding carbadox upon the growth and performance of young pigs. J. Anim. Sci. 1969, 28, 208–215. [Google Scholar] [CrossRef]
- Hill, G.M.; Mahan, D.C.; Carter, S.D.; Cromwell, G.L.; Ewan, R.C.; Harrold, R.L.; Lewis, A.J.; Miller, P.S.; Shurson, G.C.; Veum, T.L. Effect of pharmacological concentrations of zinc oxide with or without the inclusion of an antibacterial agent on nursery pig performance. J. Anim. Sci. 2001, 79, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Niewold, T.A. Why anti-inflammatory compounds are the solution for the problem with in feed antibiotics. Qual. Assur. Saf. Crops Foods 2014, 6, 119–122. [Google Scholar] [CrossRef]
- Højberg, O.; Canibe, N.; Poulsen, H.D.; Hedemann, M.S.; Jensen, B.B. Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Appl. Environ. Microbiol. 2005, 71, 2267–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Li, X.; Li, D.; Yue, T.; Fang, Q.; Ni, J.; Zhou, X.; Wu, G. Dietary supplementation with zinc oxide stimulates ghrelin secretion from the stomach of young pigs. J. Nutr. Biochem. 2009, 20, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chen, Y.; Wang, Z.; Zhou, A.; He, M.; Mao, L.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; et al. Coated zinc oxide improves intestinal immunity function and regulates microbiota composition in weaned piglets. Br. J. Nutr. 2014, 111, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhu, C.; Chen, S.; Gao, L.; Lv, H.; Feng, R.; Zhu, Q.; Xu, J.; Chen, Z.; Jiang, Z. Dietary high zinc oxide modulates the microbiome of ileum and colon in weaned piglets. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.L.; Pluske, J.R.; Morrison, R.S.; McDonald, T.N.; Smits, R.J.; Henman, D.J.; Stensland, I.; Dunshea, F.R. Post-weaning and whole-of-life performance of pigs is determined by live weight at weaning and the complexity of the diet fed after weaning. Anim. Nutr. 2017, 3, 372–379. [Google Scholar] [CrossRef]
- Hill, G.M.; Cromwell, G.L.; Crenshaw, T.D.; Dove, C.R.; Ewan, R.C.; Knabe, D.A.; Lewis, A.J.; Libal, G.W.; Mahan, D.C.; Shurson, G.C.; et al. Growth promotion effects and plasma changes from feeding high dietary concentrations of zinc and copper to weanling pigs (regional study). J. Anim. Sci. 2000, 78, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Mahan, C.; Carter, S.; Cromwell, G.; Hill, G.M.; Harrold, R.; Lewis, A.J.; Veum, T.L. Efficacy of added zinc oxide levels with or without an antibacterial agent in the postweaning diets of pigs. J. Anim. Sci. 2000, 78, 61. [Google Scholar]
- Ghoshal, N.G.; Bal, H.S. Comparative morphology of the stomach of some laboratory mammals. Lab. Anim. 1989, 23, 21–29. [Google Scholar] [CrossRef]
- Feher, J. The Stomach. In Quantitative Human Physiology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 785–795. ISBN 9780123821638. [Google Scholar]
- Carlson, D.; Poulsen, H.D.; Vestergaard, M. Additional dietary zinc for weaning piglets is associated with elevated concentrations of serum IGF-I. J. Anim. Physiol. Anim. Nutr. 2004, 88, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yin, J.; Li, D.; Chen, X.; Zang, J.; Zhou, X. Dietary supplementation with zinc oxide increases IGF-I and IGF-I receptor gene expression in the small intestine of weanling piglets. J. Nutr. 2006, 136, 1786–1791. [Google Scholar] [CrossRef] [PubMed]
- Li, B.T.; Van Kessel, A.G.; Caine, W.R.; Huang, S.X.; Kirkwood, R.N. Small intestinal morphology and bacterial populations in ileal digesta and feces of newly weaned pigs receiving a high dietary level of zinc oxide. Can. J. Anim. Sci. 2011, 81, 511–516. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Kim, Y.M.; Lee, K.Y.; Kim, I.H. Use of protected zinc oxide in lower doses in weaned pigs in substitution for the conventional high dose zinc oxide. Anim. Feed Sci. Technol. 2018, 240, 1–10. [Google Scholar] [CrossRef]
- Hedemann, M.S.; Jensen, B.B.; Poulsen, H.D. Influence of dietary zinc and copper on digestive enzyme activity and intestinal morphology in weaned pigs. J. Anim. Sci. 2006, 84, 3310–3320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovicv, R.; Šefer, D.; Krsticv, M.; Petrujkicv, B. Effect of different growth promoters on broiler performance and gut morphology. Arch. Med. Vet. 2009, 41, 163–169. [Google Scholar]
- Looft, T.; Allen, H.K.; Casey, T.A.; Alt, D.P.; Stanton, T.B. Carbadox has both temporary and lasting effects on the swine gut microbiota. Front. Microbiol. 2014, 5, 276. [Google Scholar] [CrossRef]
- Gao, Y.; Han, F.; Huang, X.; Rong, Y.; Yi, H.; Wang, Y. Changes in gut microbial populations, intestinal morphology, expression of tight junction proteins, and cytokine production between two pig breeds after challenge with Escherichia coli K88: A comparative study. J. Anim. Sci. 2013, 91, 5614–5625. [Google Scholar] [CrossRef] [Green Version]
- Clatworthy, J.P.; Subramanian, V. Stem cells and the regulation of proliferation, differentiation and patterning in the intestinal epithelium: Emerging insights from gene expression patterns, transgenic and gene ablation studies. Mech. Dev. 2001, 101, 3–9. [Google Scholar] [CrossRef]
- Niewold, T.A. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult. Sci. 2007, 86, 605–609. [Google Scholar] [CrossRef]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, X.; Xiao, Z.; Liu, L.; Wang, G.; Tao, W.; Wang, M.; Zou, J.; Leng, D. Effects of dietary zinc oxide nanoparticles supplementation on growth performance, zinc status, intestinal morphology, microflora population, and immune response in weaned pigs. J. Sci. Food Agric. 2019, 99, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Klasing, K.C. Nutrition and the immune system. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczek, O.; Vannier, E.; Borggraefe, I.; Kaser, A.; Siegmund, B.; Dinarello, C.A.; Tilg, H. Imbalance between interleukin-1 agonists and antagonists: Relationship to severity of inflammatory bowel disease. Clin. Exp. Immunol. 2004, 138, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Casini-Raggi, V.; Kam, L.; Chong, Y.J.; Fiocchi, C.; Pizarro, T.T.; Cominelli, F. Mucosal imbalance of IL-1 and IL-1 receptor antagonist in inflammatory bowel disease. A novel mechanism of chronic intestinal inflammation. J. Immunol. 1995, 154, 2434–2440. [Google Scholar] [PubMed]
- Laplace, J.P.; Darcy-Vrillon, B.; Duval-Iflah, Y.; Raibaud, P. Proteins in the digesta of the pig: Amino acid composition of endogenous, bacterial and fecal fractions. Reprod. Nutr. Dev. 1985, 25, 1083–1099. [Google Scholar] [CrossRef] [Green Version]
- Choo, J.M.; Kanno, T.; Zain, N.M.M.; Leong, L.E.X.; Abell, G.C.J.; Keeble, J.E.; Bruce, K.D.; Mason, A.J.; Rogers, G.B. Divergent relationships between fecal microbiota and metabolome following distinct antibiotic-induced disruptions. Am. Soc. Microbiol. 2017, 2, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Torrallardona, D.; Harris, C.I.; Fuller, M.F. Pigs’ gastrointestinal microflora provide them with essential amino acids. J. Nutr. 2003, 133, 1127–1131. [Google Scholar] [CrossRef] [Green Version]
- Looft, T.; Allen, H.K.; Cantarel, B.L.; Levine, U.Y.; Bayles, D.O.; Alt, D.P.; Henrissat, B.; Stanton, T.B. Bacteria, phages and pigs: The effects of in-feed antibiotics on the microbiome at different gut locations. ISME J. 2014, 8, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- van der Wielen, N.; Moughan, P.J.; Mensink, M. Amino acid absorption in the large intestine of humans and porcine models. J. Nutr. 2017, 147, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Wu, G.; Zhu, W. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [Green Version]
- Trudeau, M.P.; Zhou, Y.; Leite, F.L.; Gomez, A.; Urriola, P.E.; Shurson, G.C.; Chen, C.; Isaacson, R.E. Fecal hyodeoxycholic acid is correlated with tylosin-induced microbiome changes in growing pigs. Front. Vet. Sci. 2018, 5, 1–7. [Google Scholar] [CrossRef]
- Pols, T.W.H.; Puchner, T.; Korkmaz, H.I.; Vos, M.; Soeters, M.R.; de Vries, C.J.M. Lithocholic acid controls adaptive immune responses by inhibition of Th1 activation through the Vitamin D receptor. PLoS ONE 2017, 12, e0176715. [Google Scholar] [CrossRef] [PubMed]
- Waghray, M.; Zavros, Y.; Saqui–Salces, M.; El–Zaatari, M.; Alamelumangapuram, C.B.; Todisco, A.; Eaton, K.A.; Merchant, J.L. Interleukin-1β promotes gastric atrophy through suppression of sonic hedgehog. Gastroenterology 2010, 138, 562–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saqui-Salces, M.; Huang, Z.; Ferrandis Vila, M.; Li, J.; Mielke, J.A.; Urriola, P.E.; Shurson, G.C. Modulation of intestinal cell differentiation in growing pigs is dependent on the fiber source in the diet. J. Anim. Sci. 2017, 95, 1179–1190. [Google Scholar] [CrossRef]
- Vila, M.F.; Trudeau, M.P.; Hung, Y.T.; Zeng, Z.; Urriola, P.E.; Shurson, G.C.; Saqui-Salces, M. Dietary fiber sources and non-starch polysaccharide-degrading enzymes modify mucin expression and the immune profile of the swine ileum. PLoS ONE 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, W.; Chen, P.; Urriola, P.E.; Shurson, G.C.; Ruan, R.; Chen, C. Metabolomic evaluation of scenedesmus sp. as a feed ingredient revealed dose-dependent effects on redox balance, intermediary and microbial metabolism in a mouse model. Nutrients 2019, 11, 1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Yao, D.; Chen, C.; Lu, Y.; Yao, D.; Chen, C. 2-Hydrazinoquinoline as a derivatization agent for LC-MS-based metabolomic investigation of diabetic ketoacidosis. Metabolites 2013, 3, 993–1010. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Zn | 150 | 2500 | SE 1 | p-values | ||||
|---|---|---|---|---|---|---|---|---|
| Carbadox | No | Yes | No | Yes | Zn | AB 2 | Zn × AB 3 | |
| Stomach | ||||||||
| Mucosal height of corpus, µm | 671.5 | 606.7 | 736.3 | 723.2 | 27.52 | <0.01 | 0.13 | 0.33 |
| Mucosal height of antrum, µm | 496.8 | 489.6 | 561.8 | 534.5 | 15.11 | <0.01 | 0.25 | 0.50 |
| Gastric acid, mEq/mL | 0.065 | 0.054 | 0.058 | 0.052 | 0.006 | 0.49 | 0.19 | 0.71 |
| Jejunum | ||||||||
| Villus width, μm | 93.4 | 85.0 | 90.7 | 93.0 | 5.35 | 0.56 | 0.50 | 0.25 |
| Villus height, μm | 535.6 | 500.8 | 551.6 | 572.3 | 23.74 | 0.07 | 0.77 | 0.25 |
| Crypt depth, μm | 143.1 | 113.9 | 139.3 | 137.7 | 5.63 | 0.08 | 0.01 | 0.02 |
| Villus:crypt ratio | 3.74 a | 4.40 b | 3.99 ab | 4.17 ab | 0.09 | 0.94 | <0.01 | 0.06 |
| Ileum | ||||||||
| Villus width, μm | 71.1 | 69.3 | 68.2 | 73.5 | 2.7 | 0.81 | 0.51 | 0.19 |
| Villus height, μm | 280.4 | 286.9 | 287.8 | 308.6 | 16.3 | 0.38 | 0.41 | 0.67 |
| Crypt depth, μm | 111.0 b | 90.1 a | 98.2 ab | 99.4 ab | 5.4 | 0.75 | 0.07 | 0.05 |
| Villus:Crypt ratio | 2.64 | 3.22 | 2.93 | 3.10 | 0.17 | 0.61 | 0.03 | 0.22 |
| Peyer’s patch, μm 2 | 163,673 | 169,555 | 171,699 | 163341 | 9816 | 0.93 | 0.90 | 0.47 |
| Zn | 150 | 2500 | SE 2 | p-values | ||||
|---|---|---|---|---|---|---|---|---|
| Carbadox | No | Yes | No | Yes | Zn | AB 3 | Zn × AB 4 | |
| IFNγ | 96.70 | 85.24 | 91.06 | 84.39 | 10.04 | 0.75 | 0.38 | 0.81 |
| IL-1α | 1.19 | 1.18 | 1.16 | 0.90 | 0.19 | 0.38 | 0.43 | 0.46 |
| IL-1β | 85.27 | 44.33 | 51.78 | 34.48 | 9.75 | 0.03 | <0.01 | 0.23 |
| IL-1RA | 13.21 ab | 12.89 ab | 18.1 b | 11.24 a | 1.48 | 0.28 | 0.02 | 0.03 |
| IL-1RA:IL-1 | 0.20 | 0.32 | 0.39 | 0.35 | 0.05 | 0.04 | 0.41 | 0.13 |
| IL-4 | 19.74 | 15.07 | 15.39 | 8.48 | 7.05 | 0.46 | 0.43 | 0.88 |
| IL-6 | 1.27 a | 2.29 ab | 3.32 b | 1.34 a | 0.68 | 0.28 | 0.33 | 0.01 |
| IL-8 | 270.47 | 234.71 | 224.13 | 170.08 | 49.36 | 0.12 | 0.20 | 0.80 |
| IL-10 | 0.36 | 0.54 | 0.31 | 0.58 | 0.12 | 0.98 | 0.09 | 0.69 |
| IL-12 | 2.67 | 2.36 | 2.89 | 2.12 | 0.56 | 0.98 | 0.34 | 0.68 |
| IL-10:IL-12 | 0.14 | 0.26 | 0.15 | 0.23 | 0.05 | 0.83 | 0.07 | 0.67 |
| IL-18 | 931.05 | 889.50 | 856.91 | 765.01 | 57.45 | 0.09 | 0.25 | 0.66 |
| Zn | 150 | 2500 | SE 1 | p-values | ADG (r Value) 4 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Carbadox | No | Yes | No | Yes | Zn | AB 2 | Zn × AB 3 | ||
| Amino acids, mg/g | |||||||||
| Alanine | 0.257 | 0.252 | 0.221 | 0.270 | 0.030 | 0.78 | 0.51 | 0.41 | 0.33 |
| Arginine | 0.321 | 0.282 | 0.290 | 0.595 | 0.108 | 0.20 | 0.23 | 0.12 | 0.38 |
| Asparagine | 0.035 | 0.081 | 0.070 | 0.080 | 0.027 | 0.53 | 0.28 | 0.49 | 0.32 |
| Aspartic acid | 0.131 | 0.169 | 0.149 | 0.220 | 0.034 | 0.31 | 0.11 | 0.64 | - |
| Glutamic acid | 1.011 | 1.087 | 0.965 | 1.112 | 0.174 | 0.95 | 0.53 | 0.84 | - |
| Glutamine | 0.123 | 0.142 | 0.113 | 0.184 | 0.035 | 0.64 | 0.20 | 0.45 | 0.33 |
| Glycine | 0.211 | 0.255 | 0.191 | 0.204 | 0.035 | 0.31 | 0.42 | 0.66 | - |
| Histidine | 0.070 | 0.073 | 0.064 | 0.093 | 0.015 | 0.63 | 0.29 | 0.38 | 0.36 |
| Leucine/Isoleucine | 0.195 | 0.216 | 0.152 | 0.265 | 0.039 | 0.94 | 0.09 | 0.25 | 0.40 |
| Lysine | 0.729 | 0.612 | 0.687 | 0.818 | 0.132 | 0.54 | 0.96 | 0.35 | - |
| Methionine | 0.021 | 0.028 | 0.016 | 0.033 | 0.009 | 0.94 | 0.19 | 0.57 | 0.37 |
| Phenylalanine | 0.259 | 0.257 | 0.192 | 0.289 | 0.038 | 0.64 | 0.22 | 0.20 | - |
| Proline | 0.201 | 0.163 | 0.157 | 0.199 | 0.024 | 0.87 | 0.92 | 0.11 | 0.30 |
| Serine | 0.086 | 0.128 | 0.113 | 0.163 | 0.036 | 0.39 | 0.21 | 0.91 | 0.38 |
| Threonine | 0.073 | 0.103 | 0.075 | 0.133 | 0.023 | 0.45 | 0.07 | 0.55 | 0.44 |
| Tryptophan | 0.043 | 0.063 | 0.047 | 0.095 | 0.019 | 0.35 | 0.08 | 0.46 | 0.46 |
| Tyrosine | 0.304 | 0.294 | 0.236 | 0.391 | 0.069 | 0.84 | 0.30 | 0.24 | 0.35 |
| Valine | 0.174 | 0.192 | 0.141 | 0.241 | 0.035 | 0.83 | 0.11 | 0.26 | 0.41 |
| Fatty acids, mg/g | |||||||||
| Acetic acid | 0.376 | 0.159 | 0.148 | 0.221 | 0.069 | 0.20 | 0.27 | 0.03 | - |
| Propionic acid | 0.115 | 0.008 | 0.002 | 0.001 | 0.054 | 0.35 | 0.32 | 0.34 | - |
| Butyric acid | 0.638 | 0.018 | 0.010 | 0.013 | 0.311 | 0.32 | 0.33 | 0.33 | - |
| C6:0 | 0.130 | 0.093 | 0.098 | 0.085 | 0.020 | 0.32 | 0.22 | 0.53 | - |
| C8:0 | 0.008 | 0.007 | 0.009 | 0.008 | 0.001 | 0.35 | 0.45 | 0.71 | - |
| C10:0 | 0.002 | 0.002 | 0.009 | 0.010 | 0.002 | 0.24 | 0.73 | 0.73 | - |
| C12:0 | 0.060 | 0.061 | 0.049 | 0.029 | 0.027 | 0.42 | 0.73 | 0.70 | - |
| C14:0 | 0.035 | 0.061 | 0.024 | 0.035 | 0.015 | 0.25 | 0.23 | 0.65 | - |
| C14:1 | 0.009 | 0.005 | 0.004 | 0.006 | 0.003 | 0.57 | 0.78 | 0.30 | - |
| C15:0 | 0.009 | 0.016 | 0.008 | 0.009 | 0.003 | 0.24 | 0.25 | 0.41 | - |
| C16:0 | 1.944 | 2.371 | 2.214 | 2.419 | 0.186 | 0.38 | 0.09 | 0.55 | - |
| C16:1 | 1.044 | 0.895 | 0.498 | 0.825 | 0.309 | 0.33 | 0.78 | 0.45 | - |
| C17:0 | 0.019 | 0.042 | 0.024 | 0.023 | 0.011 | 0.52 | 0.34 | 0.30 | - |
| C17:1 | 0.089 | 0.075 | 0.035 | 0.065 | 0.025 | 0.21 | 0.75 | 0.39 | - |
| C18:0 | 4.162 | 3.314 | 3.999 | 3.425 | 0.423 | 0.95 | 0.10 | 0.75 | - |
| C18:1 | 7.775 | 8.369 | 7.792 | 9.257 | 2.250 | 0.83 | 0.62 | 0.84 | - |
| C18:2 | 6.029 | 5.992 | 5.467 | 6.364 | 1.674 | 0.95 | 0.79 | 0.77 | - |
| C18:3 | 0.830 | 0.890 | 0.770 | 0.870 | 0.310 | 0.90 | 0.79 | 0.96 | - |
| Bile acids,μg/g | |||||||||
| LCA | 0.092 | 1.001 | 0.166 | 0.067 | 0.397 | 0.27 | 0.30 | 0.20 | −0.32 |
| CDCA | 7.67 | 16.47 | 14.50 | 2.85 | 6.980 | 0.63 | 0.84 | 0.15 | - |
| CA | 0.144 | 4.615 | 0.404 | 0.202 | 1.513 | 0.17 | 0.15 | 0.12 | - |
| GDCA | 0.047 | 3.809 | 0.088 | 0.309 | 1.835 | 0.35 | 0.29 | 0.34 | - |
| TCDCA | 4.13 | 11.05 | 5.72 | 2.78 | 4.278 | 0.44 | 0.64 | 0.26 | - |
| DCA | 1.33 | 1.57 | 2.45 | 1.21 | 0.804 | 0.64 | 0.54 | 0.36 | - |
| GCA | 0.242 | 10.031 | 0.584 | 0.514 | 3.038 | 0.14 | 0.12 | 0.11 | - |
| GCDCA | 14.31 | 26.85 | 13.87 | 17.42 | 12.86 | 0.70 | 0.54 | 0.73 | - |
| TDCA | 1.991 | 1.202 | 1.830 | 2.496 | 0.954 | 0.56 | 0.95 | 0.45 | - |
| TCA | 1.499 | 3.347 | 0.500 | 0.210 | 1.597 | 0.20 | 0.63 | 0.50 | - |
| HDCA | 25.24 | 6.33 | 38.67 | 22.72 | 14.84 | 0.25 | 0.17 | 0.91 | - |
| Zn | 150 | 2500 | SE 1 | p-values | ADG (r Value) 4 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Carbadox | No | Yes | No | Yes | Zn | AB 2 | Zn × AB 3 | |||
| Amino acids, mg/g | ||||||||||
| Alanine | 0.056 | 0.081 | 0.063 | 0.069 | 0.007 | 0.77 | 0.01 | 0.13 | 0.40 | |
| Arginine | 0.006 | 0.010 | 0.011 | 0.008 | 0.002 | 0.60 | 0.69 | 0.10 | - | |
| Asparagine | 0.001 | 0.002 | 0.001 | 0.001 | 0.001 | 0.60 | 0.38 | 0.32 | - | |
| Aspartic acid | 0.050 | 0.054 | 0.052 | 0.059 | 0.006 | 0.55 | 0.31 | 0.79 | 0.32 | |
| Glutamic acid | 0.357 | 0.493 | 0.387 | 0.490 | 0.046 | 0.73 | <0.01 | 0.68 | 0.36 | |
| Glutamine | 0.007 | 0.009 | 0.007 | 0.009 | 0.002 | 0.94 | 0.20 | 0.90 | - | |
| Glycine | 0.048 | 0.076 | 0.049 | 0.055 | 0.009 | 0.29 | 0.07 | 0.22 | 0.34 | |
| Histidine | 0.011 | 0.047 | 0.010 | 0.013 | 0.009 | 0.05 | 0.03 | 0.07 | - | |
| Leucine/Isoleucine | 0.148 | 0.179 | 0.148 | 0.172 | 0.017 | 0.83 | 0.10 | 0.85 | - | |
| Lysine | 0.315 | 0.426 | 0.351 | 0.386 | 0.033 | 0.94 | 0.03 | 0.25 | - | |
| Methionine | 0.026 | 0.030 | 0.025 | 0.029 | 0.003 | 0.85 | 0.21 | 0.94 | - | |
| Phenylalanine | 0.128 | 0.156 | 0.121 | 0.132 | 0.020 | 0.42 | 0.32 | 0.68 | - | |
| Proline | 0.071 | 0.073 | 0.083 | 0.074 | 0.009 | 0.47 | 0.71 | 0.58 | - | |
| Serine | 0.050 | 0.060 | 0.052 | 0.059 | 0.005 | 0.88 | 0.04 | 0.72 | - | |
| Threonine | 0.028 | 0.034 | 0.029 | 0.035 | 0.003 | 0.85 | 0.07 | 0.93 | - | |
| Tryptophan | 0.022 | 0.018 | 0.014 | 0.016 | 0.005 | 0.28 | 0.77 | 0.55 | - | |
| Tyrosine | 0.175 | 0.178 | 0.158 | 0.180 | 0.024 | 0.76 | 0.61 | 0.70 | - | |
| Valine | 0.089 | 0.109 | 0.090 | 0.104 | 0.009 | 0.83 | 0.06 | 0.77 | - | |
| Fatty acids, mg/g | ||||||||||
| Acetic acid | 3.549 | 3.310 | 3.593 | 3.721 | 0.270 | 0.39 | 0.85 | 0.49 | 0.31 | |
| Propionic acid | 2.020 | 2.028 | 2.286 | 2.399 | 0.250 | 0.14 | 0.78 | 0.81 | - | |
| Butyric acid | 4.482 | 3.856 | 4.225 | 4.844 | 0.474 | 0.44 | 0.99 | 0.19 | - | |
| C6:0 | 0.753 | 0.828 | 0.631 | 1.313 | 0.163 | 0.22 | 0.01 | 0.04 | - | |
| C8:0 | 0.016 | 0.016 | 0.015 | 0.017 | 0.001 | 0.91 | 0.45 | 0.42 | - | |
| C10:0 | 0.009 | 0.011 | 0.011 | 0.013 | 0.001 | 0.11 | 0.14 | 0.76 | - | |
| C12:0 | 0.214 | 0.182 | 0.197 | 0.164 | 0.023 | 0.43 | 0.15 | 0.97 | - | |
| C14:0 | 0.350 | 0.351 | 0.443 | 0.442 | 0.049 | 0.06 | 1.00 | 0.98 | - | |
| C14:1 | 0.020 | 0.031 | 0.023 | 0.035 | 0.005 | 0.30 | < 0.01 | 0.89 | 0.50 | |
| C15:0 | 2.149 | 1.967 | 2.931 | 2.724 | 0.326 | 0.02 | 0.55 | 0.97 | - | |
| C16:0 | 2.931 | 2.592 | 3.317 | 2.975 | 0.205 | 0.06 | 0.10 | 0.99 | - | |
| C16:1 | 1.705 | 2.107 | 1.536 | 2.065 | 0.216 | 0.60 | 0.02 | 0.75 | - | |
| C17:0 | 1.430 | 0.838 | 2.000 | 1.591 | 0.341 | 0.05 | 0.14 | 0.79 | - | |
| C17:1 | 2.465 | 3.213 | 3.370 | 3.805 | 0.876 | 0.35 | 0.46 | 0.84 | - | |
| C18:0 | 4.603 | 3.862 | 4.658 | 4.034 | 0.347 | 0.74 | 0.05 | 0.86 | −0.23 | |
| C18:1 | 15.629 | 17.527 | 17.140 | 19.417 | 1.396 | 0.20 | 0.11 | 0.89 | 0.18 | |
| C18:2 | 5.462 | 5.976 | 6.034 | 7.150 | 0.567 | 0.10 | 0.13 | 0.57 | 0.32 | |
| C18:3 | 0.756 | 0.907 | 0.982 | 1.248 | 0.114 | 0.01 | 0.04 | 0.59 | 0.34 | |
| Bile acids, μg/g | ||||||||||
| LCA | 185.07 | 182.63 | 160.94 | 233.36 | 25.754 | 0.61 | 0.18 | 0.15 | - | |
| CDCA | 8.310 | 11.720 | 12.719 | 10.573 | 3.952 | 0.68 | 0.87 | 0.48 | - | |
| CA | 4.070 | 6.159 | 7.064 | 4.808 | 1.049 | 0.44 | 0.94 | 0.04 | - | |
| GDCA | 2.327 | 2.720 | 2.962 | 3.199 | 0.398 | 0.16 | 0.42 | 0.84 | - | |
| TCDCA | 1.152 | 1.754 | 1.207 | 1.534 | 0.331 | 0.80 | 0.16 | 0.68 | - | |
| DCA | 6.933 | 10.936 | 10.364 | 8.194 | 2.903 | 0.91 | 0.75 | 0.29 | - | |
| GCA | 0.516 | 0.651 | 0.588 | 0.516 | 0.100 | 0.72 | 0.72 | 0.24 | - | |
| GCDCA | 0.781 | 0.972 | 0.965 | 1.014 | 0.138 | 0.39 | 0.36 | 0.59 | - | |
| TDCA | 0.308 | 0.424 | 0.288 | 0.719 | 0.196 | 0.48 | 0.16 | 0.42 | - | |
| TCA | 0.048 | 0.080 | 0.078 | 0.069 | 0.019 | 0.61 | 0.54 | 0.26 | - | |
| HDCA | 129.23 | 120.74 | 140.32 | 131.94 | 17.58 | 0.53 | 0.63 | 1.00 | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, Y.-T.; Hu, Q.; Faris, R.J.; Guo, J.; Urriola, P.E.; Shurson, G.C.; Chen, C.; Saqui-Salces, M. Analysis of Gastrointestinal Responses Revealed Both Shared and Specific Targets of Zinc Oxide and Carbadox in Weaned Pigs. Antibiotics 2020, 9, 463. https://doi.org/10.3390/antibiotics9080463
Hung Y-T, Hu Q, Faris RJ, Guo J, Urriola PE, Shurson GC, Chen C, Saqui-Salces M. Analysis of Gastrointestinal Responses Revealed Both Shared and Specific Targets of Zinc Oxide and Carbadox in Weaned Pigs. Antibiotics. 2020; 9(8):463. https://doi.org/10.3390/antibiotics9080463
Chicago/Turabian StyleHung, Yuan-Tai, Qiong Hu, Richard J. Faris, Juanjuan Guo, Pedro E. Urriola, Gerald C. Shurson, Chi Chen, and Milena Saqui-Salces. 2020. "Analysis of Gastrointestinal Responses Revealed Both Shared and Specific Targets of Zinc Oxide and Carbadox in Weaned Pigs" Antibiotics 9, no. 8: 463. https://doi.org/10.3390/antibiotics9080463
APA StyleHung, Y. -T., Hu, Q., Faris, R. J., Guo, J., Urriola, P. E., Shurson, G. C., Chen, C., & Saqui-Salces, M. (2020). Analysis of Gastrointestinal Responses Revealed Both Shared and Specific Targets of Zinc Oxide and Carbadox in Weaned Pigs. Antibiotics, 9(8), 463. https://doi.org/10.3390/antibiotics9080463