
Effect of Copper Nanoparticles and Ions on Epididymis and Spermatozoa Viability of Chinese Soft-Shelled Turtles Pelodiscus sinensis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and the Test Organism
2.2. Light Microscopy
2.3. Epididymal Sperm Preparation and Viability Test
2.4. Biochemical Assay
2.5. Analysis of mRNA Expression Levels
2.6. Statistical Analysis
3. Results and Discussion
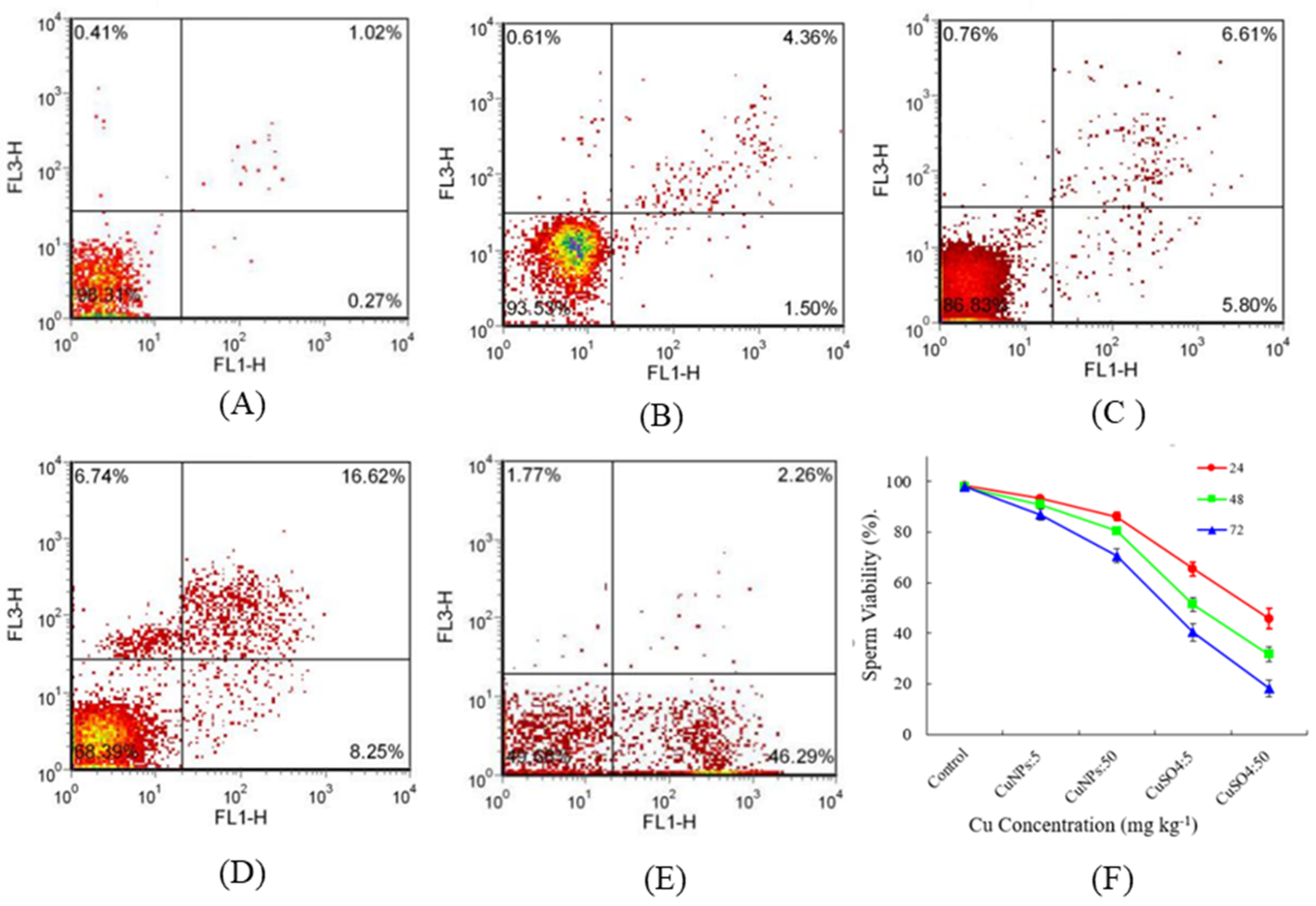
3.1. Effect of CuNPs and Cu Ions on Sperm Viability
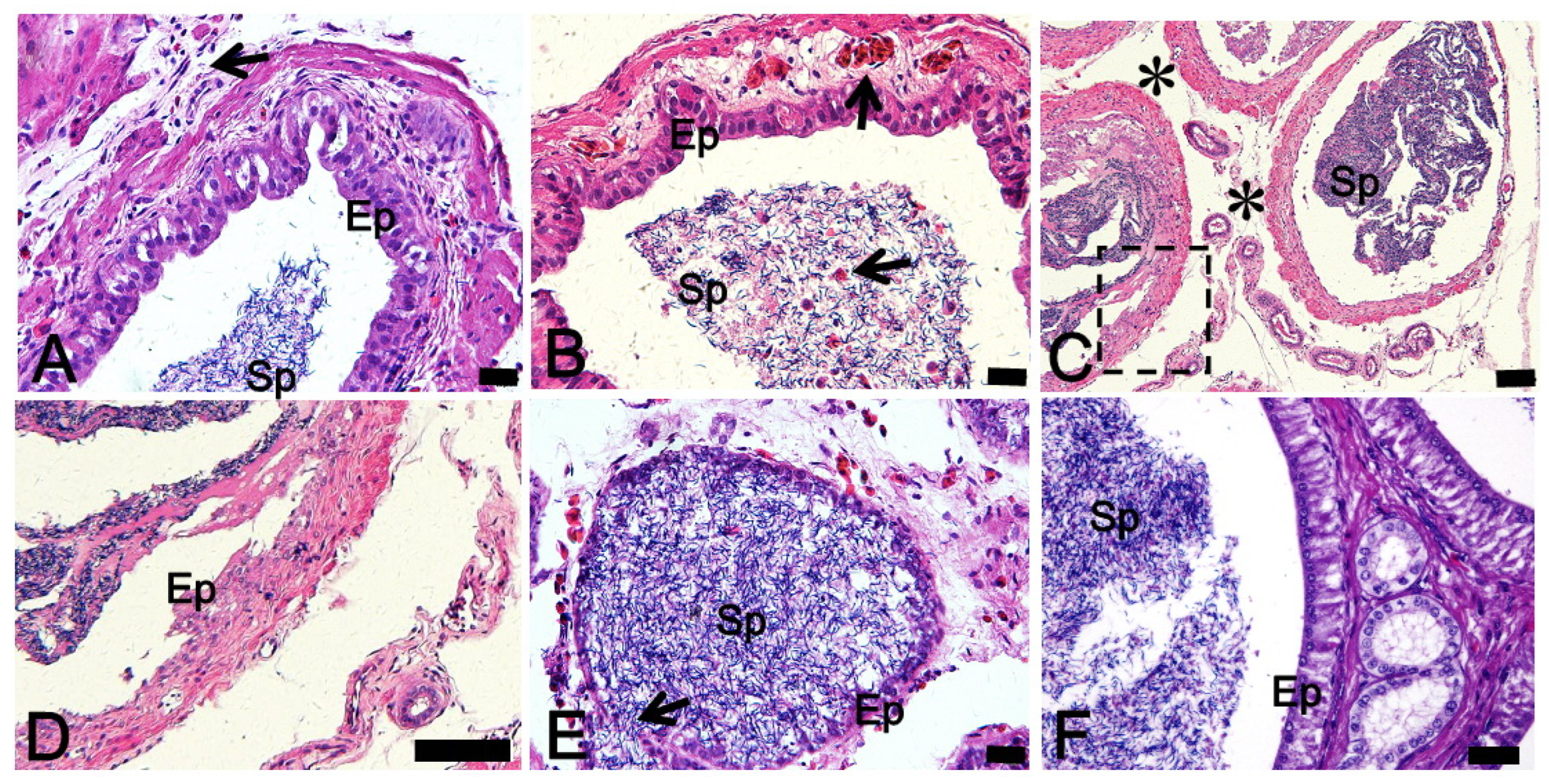
3.2. Effect of CuNPs and Cu Ions on the Histological Analysis
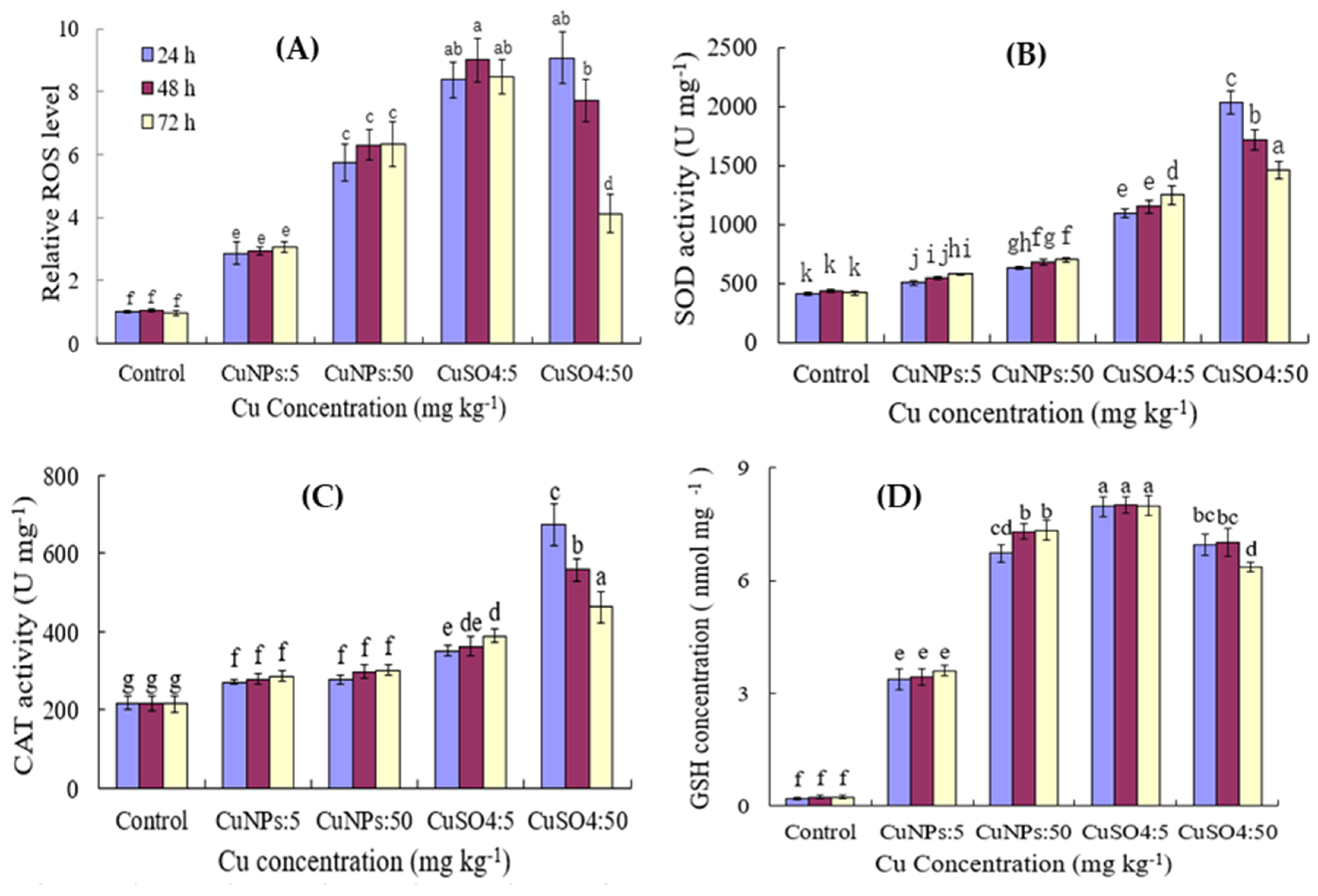
3.3. Effect of CuNPs and Cu Ions on the Antioxidant Enzyme Activity
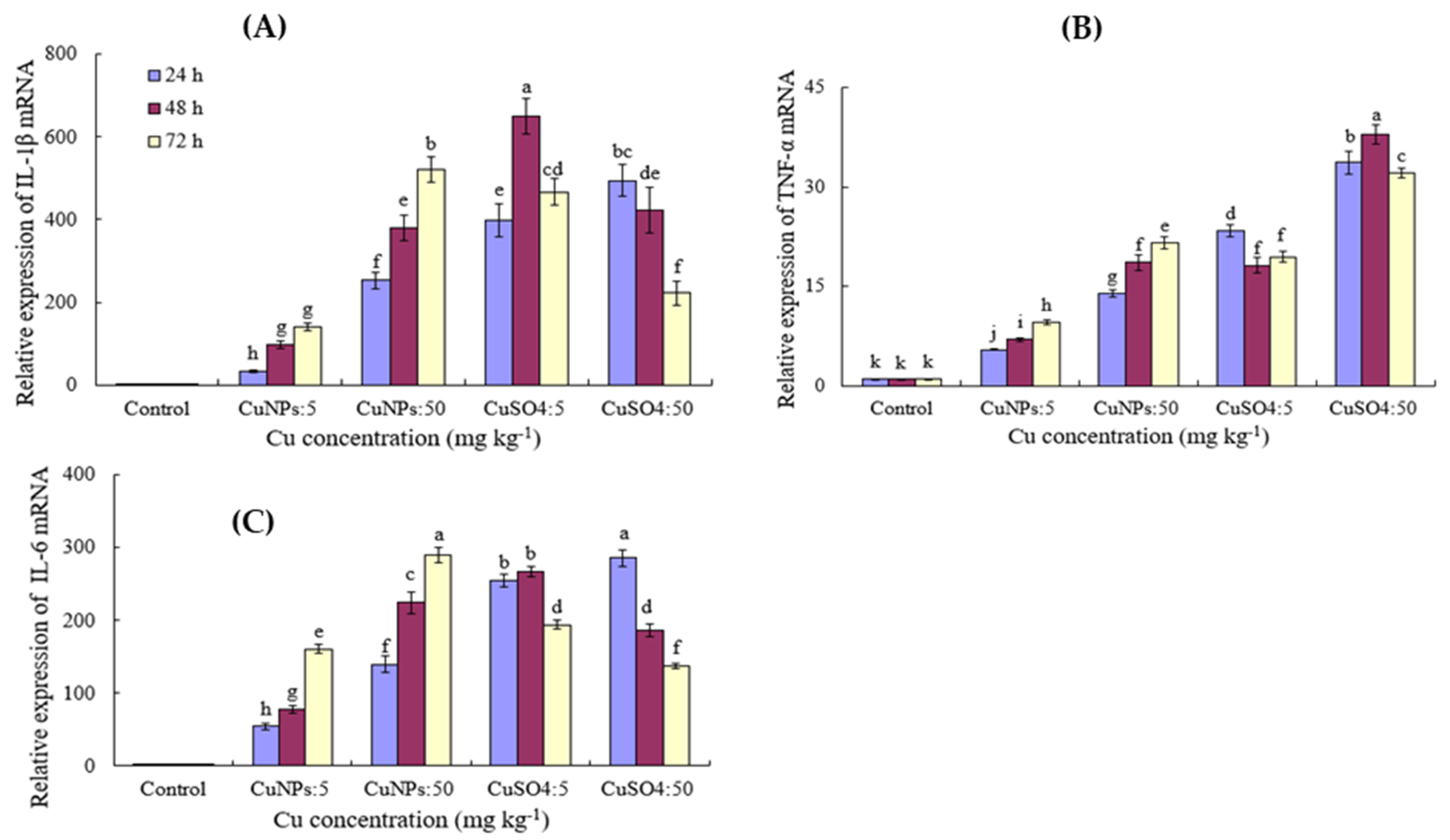
3.4. Effect of CuNPs and Cu Ions on the Relative mRNA Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, J.; Wang, X.X.; Hayat, T.; Wang, X.K. Ecotoxicological effects and mechanism of CuO nanoparticles to individual organisms. Environ. Pollut. 2017, 221, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, W.X. Comparative contributions of copper nanoparticles and ions to copper bioaccumulation and toxicity in barnacle larvae. Environ. Pollut. 2019, 249, 116–124. [Google Scholar] [CrossRef]
- Adeleye, A.S.; Oranu, E.A.; Tao, M.; Keller, A.A. Release and detection of nanosized copper from a commercial antifouling paint. Water Res. 2016, 102, 374–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller-Karanassos, C.; Arundel, W.; Lindeque, P.K.; Vance, T.; Turner, A.; Cole, M. Environmental concentrations of antifouling paint particles are toxic to sediment dwelling invertebrates. Environ. Pollut. 2020, 268, 115754. [Google Scholar] [CrossRef]
- Wu, F.; Harper, B.J.; Crandon, L.E.; Harper, S.L. Assessment of cu and CuO nanoparticle ecological responses using laboratory small-scale microcosms. Environ. Sci. Nano 2020, 7, 105–115. [Google Scholar] [CrossRef]
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and threats of contamination on aquatic ecosystems. In Bioremediation and Biotechnology; Springer: Cham, Switzerland, 2020; pp. 1–26. [Google Scholar]
- Zhu, Y.C.; Xu, J.H.; Lu, T.; Zhang, M.; Ke, M.J.; Fu, Z.W.; Pan, X.L.; Qian, H.F. A comparison of the effects of Cu nanoparticles and Cu sulfate on Phaeodactylum tricornutum physiology and transcription. Environ. Toxicol. Pharmacol. 2017, 56, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Gong, J.L.; Cao, W.C.; Chen, Z.P.; Huang, D.L.; Ye, J.; Cai, Z. The combined toxicity and mechanism of multi-walled carbon nanotubes and nano copper oxide toward freshwater algae: Tetradesmus obliquus. J. Environ. Sci. 2022, 112, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, F.; Xiang, J. Acute effects of cadmium and copper on survival, oxygen consumption, ammonia-N excretion, and metal accumulation in juvenile Exopalaemon carinicauda. Ecotoxicol. Environ. Saf. 2014, 104, 209–214. [Google Scholar] [CrossRef]
- Bat, L.; Gündoğdu, A.; Sezgin, M.; Çulha, M.; Gönlügür, G.; Akbulut, M. Acute toxicity of zinc, copper and lead to three species of marine organisms from the Sinop Peninsula, Black Sea. Turk. J. Biol. 1999, 23, 537–544. [Google Scholar]
- Vardhanan, Y.S.; Radhakrishnan, T. Acute toxicity evaluation of copper, arsenic and HCH to paddy field crab, Paratelphusa hydrodromus (Herb). J. Environ. Biol. 2002, 23, 387–392. [Google Scholar] [PubMed]
- Chourpagar, A.R.; Kulkarni, G.K. Heavy metal toxicity to a freshwater crab, Barytelphusa cunicularis (westwood) from aurangabad region. Recent Res. Sci. Technol. 2011, 3, 1–5. [Google Scholar]
- Bini, G.; Chelazzi, G. Acclimatable cardiac and ventilatory responses to copper in the freshwater crayfish Procambarus clarkii. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 235–241. [Google Scholar] [CrossRef]
- Malhotra, N.; Ger, T.R.; Uapipatanakul, B.; Huang, J.C.; Chen, K.H.; Hsiao, C.D. Review of Copper and Copper Nanoparticle Toxicity in Fish. Nanomaterials 2020, 10, 1126. [Google Scholar] [CrossRef] [PubMed]
- Boyle, D.; Clark, N.J.; Handy, R.D. Toxicities of copper oxide nanomaterial and copper sulphate in early life stage zebrafish: Effects of pH and intermittent pulse exposure. Ecotoxicol. Environ. Saf. 2020, 190, 109985. [Google Scholar] [CrossRef]
- Adam, N.; Schmitt, C.; De Bruyn, L.; Knapen, D.; Blust, R. Aquatic acute species sensitivity distributions of ZnO and CuO nanoparticles. Sci. Total Environ. 2015, 526, 233–242. [Google Scholar] [CrossRef]
- Jiang, C.J.; Castellon, B.T.; Matson, C.W.; Aiken, G.R.; Hsu-Kim, H. Relative contributions of Cu oxide nanoparticles and dissolved Cu to Cu uptake kinetics of gulf killifish (Fundulus grandis) embryos. Environ. Sci. Technol. 2017, 51, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Vijver, M.G.; Richardson, M.K.; Ahmad, F.; Peijnenburg, W.J. Particlespecific toxic effects of differently shaped zinc oxide nanoparticles to zebrafish embryos (Danio rerio). Environ. Toxicol. Chem. 2014, 33, 2859–2868. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, D.; Xie, M.W.; Li, Q.B.; Liu, Q.M. Behavior and toxicity of zinc oxide nanoparticles in aquatic environment. Environ. Chem. 2016, 35, 2528–2534. [Google Scholar]
- Volland, M.; Hampel, M.; Katsumiti, A.; Yeste, M.P.; Gatica, J.M.; Cajaraville, M.; Blasco, J. Synthesis methods influence characteristics, behaviour and toxicity of bare CuO NPs compared to bulk CuO and ionic Cu after in vitro exposure of Ruditapes philippinarum hemocytes. Aquat. Toxicol. 2018, 199, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Ding, Z.C.; Zhao, G.; Zhang, T.; Xu, Q.H.; Cui, B.; Liu, J.X. Transcriptional responses and mechanisms of Cu nanoparticle toxicology on zebrafish embryos. J. Hazard Mater. 2018, 344, 1057–1068. [Google Scholar] [CrossRef]
- Lopes, F.M.; Varela Junior, A.S.; Corcini, C.D.; Da Silva, A.C.; Guazzelli, V.G.; Tavares, G.; Da Rosa, C.E. Effect of glyphosate on the sperm quality of zebrafish Danio rerio. Aquat. Toxicol. 2014, 155, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Devaux, A.; Bony, S.; Plenet, S.; Sagnes, P.; Segura, S.; Suaire, R.; Novack, M.; Gilles, A.; Olivier, J.M. Field evidence of reproduction impairment through sperm DNA damage in the fish nase (Chondrostoma nasus) in anthropized hydrosystems. Aquat. Toxicol. 2015, 169, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Góralska, M.; Dziewulsk, K.; Kulasz, M. Effect of copper nanoparticles and ions on spermatozoa motility of sea trout (Salmo trutta m. Trutta L.). Aquat. Toxicol. 2019, 211, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, B.; Maleki, A.; Johari, S.A.; Shahmoradi, B.; Mohammadi, E.; Davari, B. Histopathological effects of copper oxide nanoparticles on the gill and intestine of common carp (Cyprinus carpio) in the presence of titanium dioxide nanoparticles. Chem. Ecol. 2017, 33, 295–308. [Google Scholar] [CrossRef]
- Gürkan, M.; Gürkan, S.E.; Yılmaz, S.; Ate, M. Comparative toxicity of Alpha and Gamma iron oxide nanoparticles in Rainbow Trout: Histopathology, hematology, accumulation, and oxidative stress. Water Air Soil Pollut. 2021, 232, 37–52. [Google Scholar] [CrossRef]
- Kim, E.H.; Jeong, J.A.; Choi, E.K.; Jeong, T.Y. Antioxidant enzyme activity in Daphnia magna under microscopic observation and shed carapace length as an alternative growth endpoint. Sci. Total Environ. 2021, 794, 148771. [Google Scholar] [CrossRef]
- Chen, B.J.; Zhang, W.Y.; Niu, C.J.; Li, W.J.; Jia, H.; Storey, K.B. Antioxidant response to acute cold exposure and during recovery in juvenile Chinese soft-shelled turtles (Pelodiscus sinensis). J. Exp. Biol. 2019, 222, 197863. [Google Scholar] [CrossRef] [Green Version]
- Sanpradit, P.; Buapet, P.; Kongseng, S.; Peerakietkhajorn, S. Temperature and concentration of ZnO particles affect life history traits and oxidative stress in Daphnia magna. Aquat. Toxicol. 2020, 224, 105517. [Google Scholar] [CrossRef]
- Zhang, W.J.; Gao, J.L.; Lu, L.; Bold, T.; Li, X.; Wang, S.; Chang, Z.S.; Chen, J.; Kong, X.; Zheng, Y.X.; et al. Intracellular GSH/GST antioxidants system change as an earlier biomarker for toxicity evaluation of iron oxide nanoparticles. NanoImpact 2021, 23, 100338. [Google Scholar] [CrossRef]
- Zarski, D.; Cejko, B.I.; Krejszeff, S.; Palinska-Zarska, K.; Horvath, A.; Sarosiek, B.; Judycka, S.; Kowalski, R.K.; Laczynska, B.; Kucharczyk, D. The effect of osmolality on egg fertilization in common carp, Cyprinus carpio Linnaeus, 1758. J. Appl. Ichthyol. 2015, 31, 159–163. [Google Scholar] [CrossRef]
- Bian, X.G.; Gandahi, J.A.; Liu, Y.; Yang, P.; Liu, Y.; Zhang, L.L.; Zhang, Q.; Chen, Q.S. The ultrastructural characteristics of the spermatozoa stored in the cauda epididymidis in Chinese soft-shelled turtle Pelodiscus sinensis during the breeding season. Micron 2013, 44, 202–209. [Google Scholar] [CrossRef]
- Zebral, Y.D.; Anni, I.S.A.; Junior, A.S.V.; Corcini, C.D.; Silva, J.C.; Caldas, J.S.; Acosta, I.B.; Afonso, S.B.; Bianchini, A. Life-time exposure to waterborne copper IV: Sperm quality parameters are negatively affected in the killifish Poecilia vivipara. Chemosphere 2019, 236, 124332. [Google Scholar] [CrossRef] [PubMed]
- Sakhaee, E.; Emadi, L.; Abshenas, J.; Kheirandish, R.; Azari, O.; Amiri, E. Evaluation of epididymal sperm quality following experimentally induced copper poisoning in male rats. Andrologia 2012, 44, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, S.; Massanyi, P. In vitro copper inhibition of the rabbit spermatozoa motility. J. Environ. Sci. Health Part A 2008, 43, 651–656. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma membrane changes during programmed cell deaths. Cell Res. 2018, 28, 9. [Google Scholar] [CrossRef]
- Yuan, J.L.; Gu, Z.M.; Zheng, Y.; Zhang, Y.Y.; Gao, J.G.; Chen, S.; Wang, Z.Z. Accumulation and detoxification dynamics of microcystin-LR and antioxidant responses in male red swamp crayfish Procambarus clarkii. Aquat. Toxicol. 2016, 177, 8–18. [Google Scholar] [CrossRef]
- Mahmoud, A.E.; Mahmoud, M.A.D.; Amaal, M.; Radwa, M.S. Effects of titanium dioxide nanoparticles on red swamp crayfish, Procambarus clarkii: Bioaccumulation, oxidative stress and histopathological biomarkers. Egypt. J. Aquat. Res. 2019, 45, 11–18. [Google Scholar]
- Bagaitkar, J.; Huang, J.; Zeng, M.Y.; Pech, N.K.; Monlish, D.A.; Perez-Zapata, L.J.; Miralda, I.; Schuettpelz, L.G.; Dinauer, M.C. NADPH oxidase activation regulates apoptotic neutrophil clearance by murine macrophages. Blood 2018, 131, 2367–2378. [Google Scholar] [CrossRef]
- Mostek, A.; Słowińska, M.; Judycka, S.; Karol, H.; Ciereszko, A.; Dietrich, M.A. Identification of oxidatively modified proteins due to cryopreservation of carp semen. J. Anim. Sci. 2018, 96, 1453–1465. [Google Scholar] [CrossRef] [Green Version]
- Varfolomeev, E.E.; Ashkenazi, A. Tumor Necrosis Factor. Cell 2004, 116, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Reis, M.I.R.; Vale, A.D.; Pereira, P.J.B.; Azevedo, J.E.; dos Santo, N.M.S. Caspase-1 and IL-1β Processing in a Teleost Fish. PLoS ONE 2012, 7, 50450. [Google Scholar]
- Zhang, A.; Chen, D.; Wei, H.; Du, L.; Zhao, T.; Wang, X.; Hong, Z. Functional characterization of TNF-α in grass carp head kidney leukocytes: Induction and involvement in the regulation of NF-κB signaling. Fish Shellfish Immun. 2012, 33, 1123–1132. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes Primer Sequence (5′-3′) | Annealing (°C) | Length (bp) | GenBank Accession Number |
|---|---|---|---|
| psIL-1β Fwd GACAATGACTTGAGCAGCA psIL-1β Rev AGCCATGTTCAGCCTCACTT | 60 | 226 | JX846915 |
| psTNF-α Fwd CAGAGAGCAGGCCCTATGAC psTNF-α Rev TTGTTCCCCTTGAAGACCAC | 60 | 218 | XM_006127997 |
| psIL-6 Fwd GGAGATGCTTGCCCAAAAT psIL-6 Rev TGCACCAGGATTTTTCACC | 60 | 244 | XM_006138351 |
| β-actin-Fwd TGTTACCCATACTGTGCCCATC β-actin-Rev TAGCCATCTCCTGTTCAAAATCC | 60 | 212 | EU727174 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wei, Y.; Gao, S.; Wang, Q.; Chen, J.; Tang, B.; Bian, X. Effect of Copper Nanoparticles and Ions on Epididymis and Spermatozoa Viability of Chinese Soft-Shelled Turtles Pelodiscus sinensis. Coatings 2022, 12, 110. https://doi.org/10.3390/coatings12020110
Yang L, Wei Y, Gao S, Wang Q, Chen J, Tang B, Bian X. Effect of Copper Nanoparticles and Ions on Epididymis and Spermatozoa Viability of Chinese Soft-Shelled Turtles Pelodiscus sinensis. Coatings. 2022; 12(2):110. https://doi.org/10.3390/coatings12020110
Chicago/Turabian StyleYang, Li, Yating Wei, Shuai Gao, Qifei Wang, Jiaqi Chen, Boping Tang, and Xunguang Bian. 2022. "Effect of Copper Nanoparticles and Ions on Epididymis and Spermatozoa Viability of Chinese Soft-Shelled Turtles Pelodiscus sinensis" Coatings 12, no. 2: 110. https://doi.org/10.3390/coatings12020110
APA StyleYang, L., Wei, Y., Gao, S., Wang, Q., Chen, J., Tang, B., & Bian, X. (2022). Effect of Copper Nanoparticles and Ions on Epididymis and Spermatozoa Viability of Chinese Soft-Shelled Turtles Pelodiscus sinensis. Coatings, 12(2), 110. https://doi.org/10.3390/coatings12020110