Routes towards Novel Collagen-Like Biomaterials


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
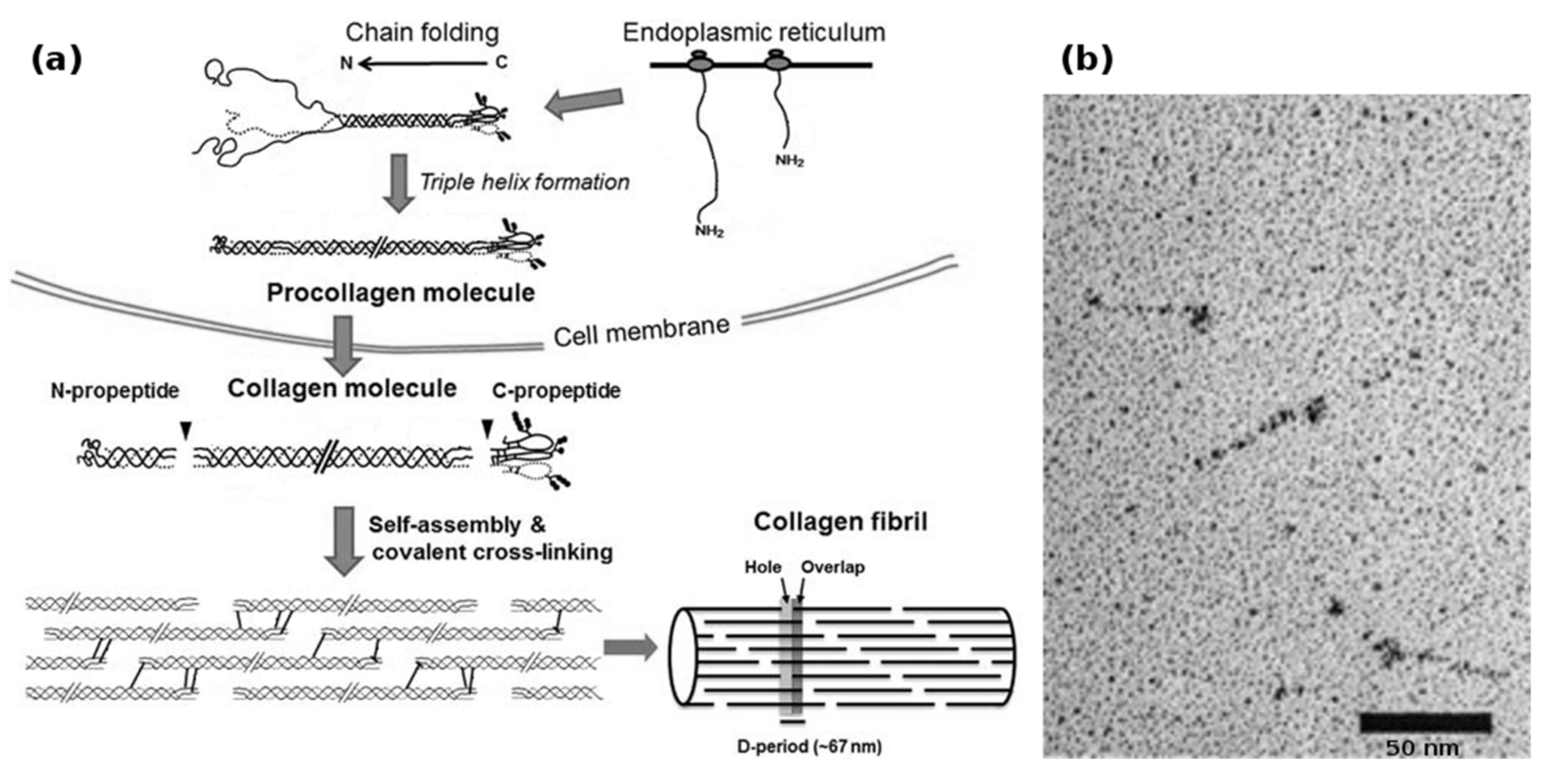
1.1. Molecular Structure of Collagen
1.2. Motivation
2. Collagen as a Biomaterial
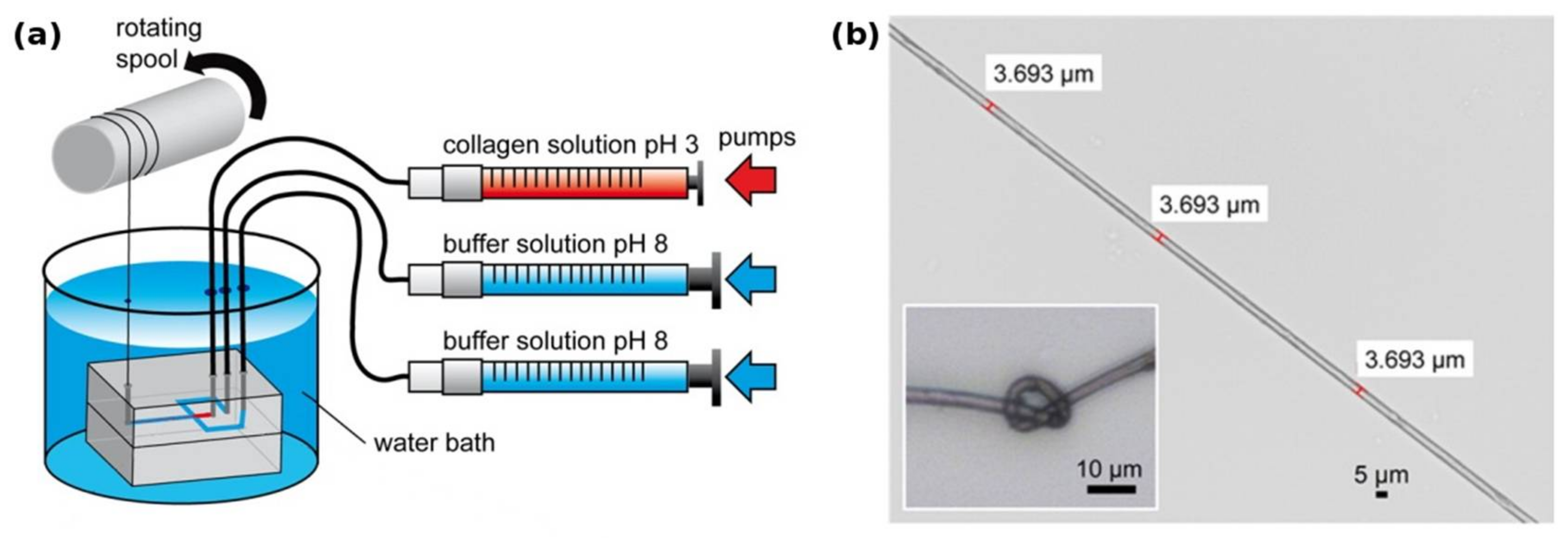
2.1. Towards Spun Collagen Fibers
2.2. Collagen-Analogues as Basis for Future Biomaterials
3. Mussel Byssus‒Silk, Collagen or Both?
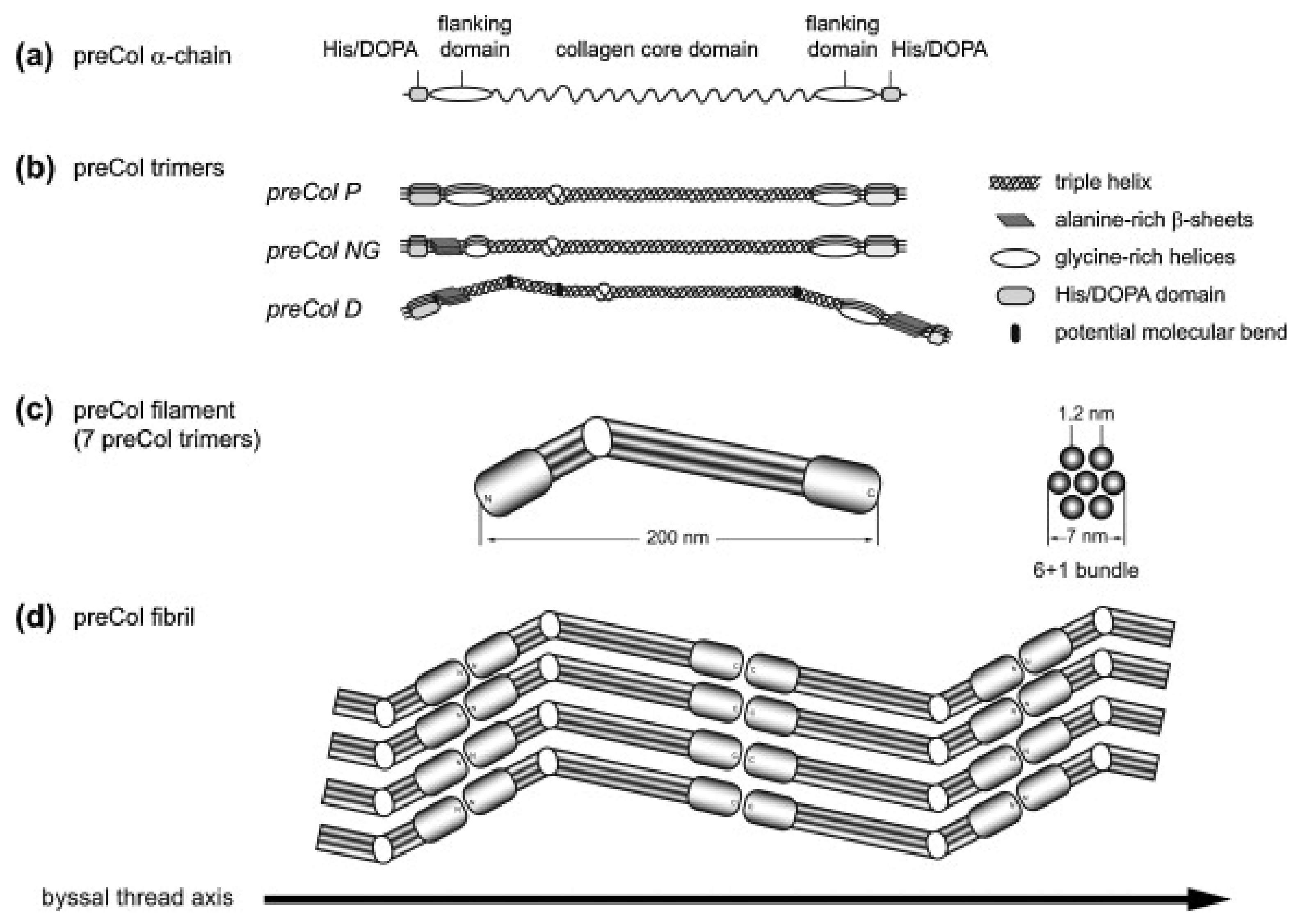
3.1. Hierarchical Assembly and Structure of preCols in the Byssus
3.2. Chemical Modifications of Byssus Proteins
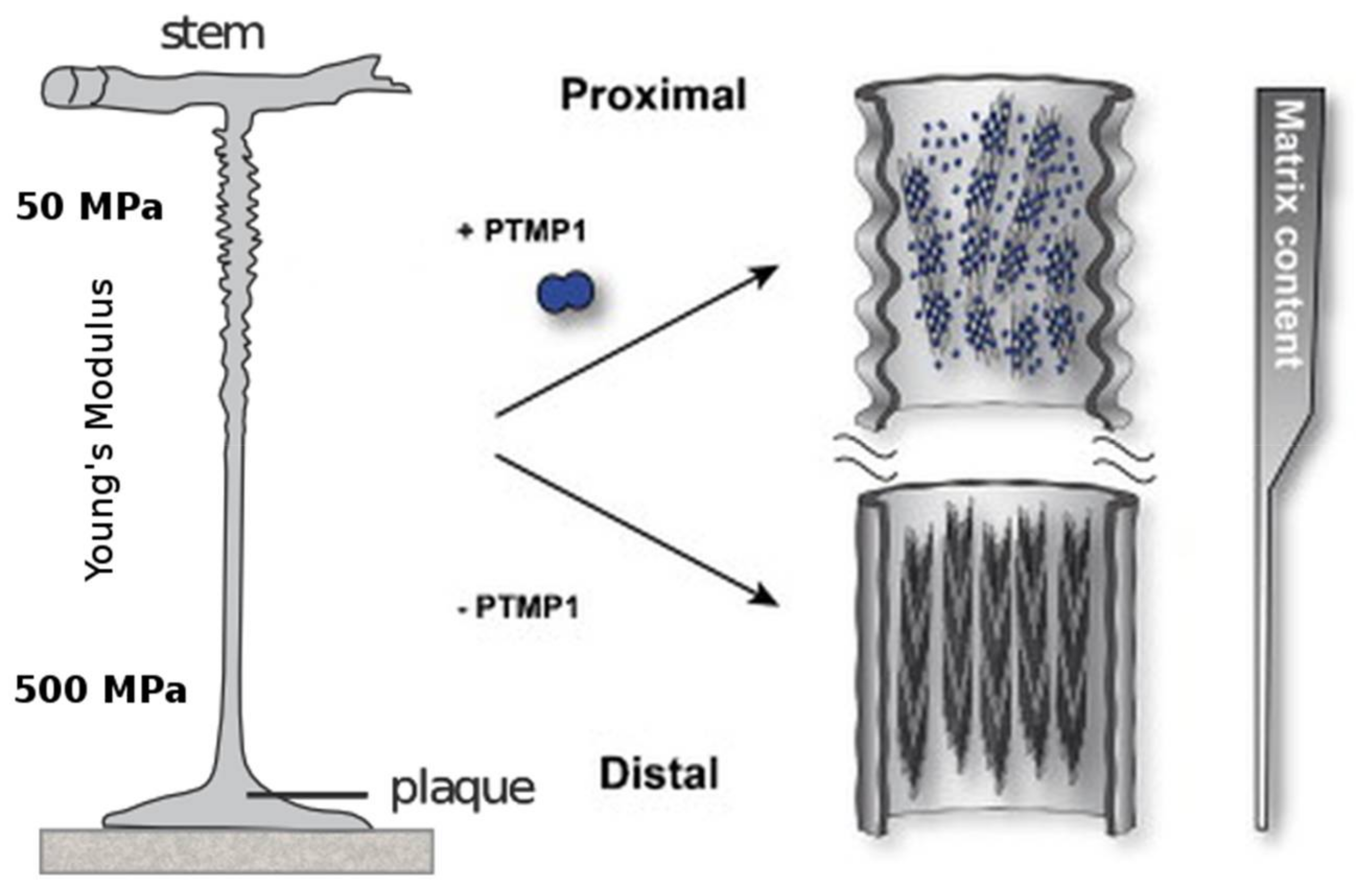
3.3. preCol-Based Biomaterials
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bouhlel, Z.; Genard, B.; Ibrahim, N.; Carrington, E.; Babarro, J.M.F.; Lok, A.; Flores, A.A.V.; Pellerin, C.; Tremblay, R.; Marcotte, I. Interspecies comparison of the mechanical properties and biochemical composition of byssal threads. J. Exp. Biol. 2017, 220, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, T.D.; Peng, Y.Y.; Trueman, H.E.; Weisman, S.; Okada, S.; Walker, A.A.; Sriskantha, A.; White, J.F.; Huson, M.G.; Werkmeister, J.A.; et al. A new class of animal collagen masquerading as an insect silk. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Fidler, A.L.; Vanacore, R.M.; Chetyrkin, S.V.; Pedchenko, V.K.; Bhave, G.; Yin, V.P.; Stothers, C.L.; Rose, K.L.; McDonald, W.H.; Clark, T.A.; et al. A unique covalent bond in basement membrane is a primordial innovation for tissue evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.W.; Okamura, B.; Hartikainen, H.; Secombes, C.J. A novel minicollagen gene links cnidarians and myxozoans. Proc. R. Soc. B 2011, 278, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Exposito, J.-Y.; Cluzel, C.; Garrone, R.; Lethias, C. Evolution of collagens. Anat. Rec. 2002, 268, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Lukomski, S.; Bachert, B.A.; Squeglia, F.; Berisio, R. Collagen-like proteins of pathogenic streptococci: Streptococcal collagen-like proteins. Mol. Microbiol. 2017, 103, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Bella, J.; Hulmes, D.J.S. Fibrillar Collagens. In Fibrous Proteins: Structures and Mechanisms; Parry, D.A.D., Squire, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Pacak, C.A.; MacKay, A.A.; Cowan, D.B. An improved method for the preparation of type I collagen from skin. J. Vis. Exp. JoVE 2014. [Google Scholar] [CrossRef] [PubMed]
- Mocan, E.; Tagadiuc, O.; Nacu, V. Aspects of collagen isolation procedure. Curiecul Med. 2011, 2, 320. [Google Scholar]
- Yaari, A.; Schilt, Y.; Tamburu, C.; Raviv, U.; Shoseyov, O. Wet spinning and drawing of human recombinant collagen. ACS Biomater. Sci. Eng. 2016, 2, 349–360. [Google Scholar] [CrossRef]
- Haynl, C.; Hofmann, E.; Pawar, K.; Förster, S.; Scheibel, T. Microfluidics-produced collagen fibers show extraordinary mechanical properties. Nano Lett. 2016, 16, 5917–5922. [Google Scholar] [CrossRef] [PubMed]
- Zeugolis, D.I.; Paul, R.G.; Attenburrow, G. Post-self-assembly experimentation on extruded collagen fibres for tissue engineering applications. Acta Biomater. 2008, 4, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Vesentini, S.; Redaelli, A.; Buehler, M.J. Hierarchical structure and nanomechanics of collagen microfibrils from the atomistic scale up. Nano Lett. 2011, 11, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.X.; Coyne, K.J.; Waite, J.H. Tough tendons. Mussel byssus has collagen with silk-like domains. J. Biol. Chem. 1997, 272, 32623–32627. [Google Scholar] [CrossRef]
- Tromp, G.; Kuivaniemi, H.; Stacey, A.; Shikata, H.; Baldwin, C.T.; Jaenisch, R.; Prockop, D.J. Structure of a full-length cDNA clone for the prepro alpha 1(I) chain of human type I procollagen. Biochem. J. 1988, 253, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Bansal, M. Collagen structure: The madras triple helix and the current scenario. IUBMB Life 2005, 57, 161–172. [Google Scholar] [CrossRef]
- Long, C.G.; Braswell, E.; Zhu, D.; Apigo, J.; Baum, J.; Brodsky, B. Characterization of collagen-like peptides containing interruptions in the repeating Gly-X-Y sequence. Biochemistry 1993, 32, 11688–11695. [Google Scholar] [CrossRef] [PubMed]
- Beck, K.; Chan, V.C.; Shenoy, N.; Kirkpatrick, A.; Ramshaw, J.A.M.; Brodsky, B. Destabilization of osteogenesis imperfecta collagen-like model peptides correlates with the identity of the residue replacing glycine. Proc. Natl. Acad. Sci. USA 2000, 97, 4273–4278. [Google Scholar] [CrossRef] [PubMed]
- Van Robertson, W.B. The effect of ascorbic acid deficiency on the collagen concentration of newly induced fibrous tissue. J. Biol. Chem. 1952, 196, 403–408. [Google Scholar] [PubMed]
- Yu, Z.; Mirochnitchenko, O.; Xu, C.; Yoshizumi, A.; Brodsky, B.; Inouye, M. Noncollagenous region of the streptococcal collagen-like protein is a trimerization domain that supports refolding of adjacent homologous and heterologous collagenous domains. Protein Sci. 2010, 19, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Keene, D.R.; Bujnicki, J.M.; Höök, M.; Lukomski, S. Streptococcal Scl1 and Scl2 proteins form collagen-like triple helices. J. Biol. Chem. 2002, 277, 27312–27318. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Zwiefka, A.; Caswell, C.C.; Xu, Y.; Keene, D.R.; Lukomska, E.; Zhao, Z.; Höök, M.; Lukomski, S. Assessment of prokaryotic collagen-like sequences derived from streptococcal Scl1 and Scl2 proteins as a source of recombinant GXY polymers. Appl. Microbiol. Biotechnol. 2006, 72, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Lukomski, S.; Nakashima, K.; Abdi, I.; Cipriano, V.J.; Shelvin, B.J.; Graviss, E.A.; Musser, J.M. Identification and characterization of a second extracellular collagen-like protein made by group A Streptococcus: Control of production at the level of translation. Infect. Immun. 2001, 69, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Harrington, M.J.; Waite, J.H. PH-dependent locking of giant mesogens in fibers drawn from mussel byssal collagens. Biomacromolecules 2008, 9, 1480–1486. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, D.; Oliveira, P.F.; Prosdocimi, F.; Rech, E.L. Review protein families, natural history and biotechnological aspects of spider silk. Genet. Mol. Res. 2012, 11, 2360–2380. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F.; Knight, D.P. Liquid crystalline spinning of spider silk. Nature 2001, 410, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Singla, A.; Lee, Y. Biomedical applications of collagen. Int. J. Pharm. 2001, 221, 1–22. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Raines, R.T. Review collagen-based biomaterials for wound healing: Collagen-based biomaterials. Biopolymers 2014, 101, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T. Evolution of catgut ligatures: The endeavours and success of Joseph Lister and William Macewen. Br. J. Surg. 1990, 77, 824–825. [Google Scholar] [CrossRef] [PubMed]
- Fung, Y.-C. Biomechanics; Springer: New York, NY, USA, 1993. [Google Scholar]
- Lee, S.; Hailey, D.M.; Lea, A.R. Tensile strength requirements for sutures. J. Pharm. Pharmacol. 1983, 35, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Siriwardane, M.L.; DeRosa, K.; Collins, G.; Pfister, B.J. Controlled formation of cross-linked collagen fibers for neural tissue engineering applications. Biofabrication 2014, 6, 015012. [Google Scholar] [CrossRef] [PubMed]
- Zeide, D.A. Adverse reactions to collagen implants. Clin. Dermatol. 1986, 4, 176–182. [Google Scholar] [CrossRef]
- Liscic, R.; Brinar, V.; Miklic, P.; Barsić, B.; Himbele, J. Creutzfeldt-Jakob disease in a patient with a lyophilized dura mater graft. Acta Med. Croatica 1999, 53, 93–96. [Google Scholar] [PubMed]
- Sabeh, F.; Shimizu-Hirota, R.; Weiss, S.J. Protease-dependent versus -independent cancer cell invasion programs: Three-dimensional amoeboid movement revisited. J. Cell Biol. 2009, 185, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Piez, K.A.; Reddi, A.H. Extracellular Matrix Biochemistry; Elsevier Science Ltd.: New York, NY, USA, 1984. [Google Scholar]
- Wu, X.; Black, L.; Santacana-Laffitte, G.; Patrick, C.W. Preparation and assessment of glutaraldehyde-crosslinked collagen–chitosan hydrogels for adipose tissue engineering. J. Biomed. Mater. Res. A 2007, 81A, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bauman, L. CosmoDerm/CosmoPlast (human bioengineered collagen) for the aging face. Facial Plast. Surg. 2004, 20, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.G.; Lovdahl, C.M.; Miles, A.J.; Hagen, V.L.; Fields, G.B. Solid-phase synthesis and stability of triple-helical peptides incorporating native collagen sequences. Biopolymers 1993, 33, 1695–1707. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Fan, P.; Brodsky, B.; Baum, J. Two-dimensional NMR assignments and conformation of (Pro-Hyp-Gly)10 and a designed collagen triple-helical peptide. Biochemistry 1993, 32, 7377–7387. [Google Scholar] [CrossRef] [PubMed]
- Cejas, M.A.; Kinney, W.A.; Chen, C.; Leo, G.C.; Tounge, B.A.; Vinter, J.G.; Joshi, P.P.; Maryanoff, B.E. Collagen-related peptides: Self-assembly of short, single strands into a functional biomaterial of micrometer scale. J. Am. Chem. Soc. 2007, 129, 2202–2203. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, S.K.; Taylor, K.M.; Bretscher, L.E.; Raines, R.T. Code for collagen’s stability deciphered. Nature 1998, 392, 666–667. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.; Bhumralkar, M.; Jefferson, E.A.; Kwak, J.; Locardi, E. Collagen mimetics. Biopolymers 1998, 47, 127–142. [Google Scholar] [CrossRef]
- Bachmann, A.; Kiefhaber, T.; Boudko, S.; Engel, J.; Bächinger, H.P. Collagen triple-helix formation in all-trans chains proceeds by a nucleation/growth mechanism with a purely entropic barrier. Proc. Natl. Acad. Sci. USA 2005, 102, 13897–13902. [Google Scholar] [CrossRef] [PubMed]
- Boudko, S.; Frank, S.; Kammerer, R.A.; Stetefeld, J.; Schulthess, T.; Landwehr, R.; Lustig, A.; Bächinger, H.P.; Engel, J. Nucleation and propagation of the collagen triple helix in single-chain and trimerized peptides: Transition from third to first order kinetics. J. Mol. Biol. 2002, 317, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Boudko, S.; Mizuno, K.; Schulthess, T.; Engel, J.; Bächinger, H.P. Collagen triple helix formation can be nucleated at either end. J. Biol. Chem. 2003, 278, 7747–7750. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Wong, D.; Kinberger, G.A.; Kwok, S.W.; Taulane, J.P.; Goodman, M. Facile and efficient assembly of collagen-like triple helices on a TRIS scaffold. Bioorg. Chem. 2007, 35, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.; De Capua, A.; Locardi, E.; Goodman, M. TREN (Tris(2-aminoethyl)amine): An effective scaffold for the assembly of triple helical collagen mimetic structures. J. Am. Chem. Soc. 2002, 124, 14085–14091. [Google Scholar] [CrossRef] [PubMed]
- Greiche, Y.; Heidemann, E. Collagen model peptides with antiparallel structure. Biopolymers 1979, 18, 2359–2361. [Google Scholar] [CrossRef]
- Barth, D.; Kyrieleis, O.; Frank, S.; Renner, C.; Moroder, L. The role of cystine knots in collagen folding and stability, Part II. conformational properties of (pro-hyp-gly)n model trimers with N- and C-terminal collagen type III cystine knots. Chem. Eur. J. 2003, 9, 3703–3714. [Google Scholar] [CrossRef] [PubMed]
- Boudko, S.P.; Engel, J.; Bächinger, H.P. The crucial role of trimerization domains in collagen folding. Int. J. Biochem. Cell Biol. 2012, 44, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Paramonov, S.E.; Gauba, V.; Hartgerink, J.D. Synthesis of collagen-like peptide polymers by native chemical ligation. Macromolecules 2005, 38, 7555–7561. [Google Scholar] [CrossRef]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Hagenau, A.; Scheibel, T. Towards the recombinant production of mussel byssal collagens. J. Adhes. 2010, 86, 10–24. [Google Scholar] [CrossRef]
- Fan, D. Proteine du Type Collagene Humain et Son Procede de Production. Patent WO2003106494A1, 24 December 2003. [Google Scholar]
- Schnicker, N.J.; Dey, M. Bacillus anthracis prolyl 4-Hydroxylase modifies collagen-like substrates in asymmetric patterns. J. Biol. Chem. 2016, 291, 13360–13374. [Google Scholar] [CrossRef] [PubMed]
- Schnicker, N.J.; Razzaghi, M.; Thakurta, S.G.; Chakravarthy, S.; Dey, M. Bacillus anthracis prolyl 4-hydroxylase interacts with and modifies elongation factor Tu. Biochemistry 2017, 56, 5771–5785. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yang, X.; Hang, B.; Li, J.; Huang, L.; Huang, F.; Xu, Z. efficient production of hydroxylated human-like collagen via the co-expression of three key genes in escherichia coli origami (DE3). Appl. Biochem. Biotechnol. 2016, 178, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Pozzolini, M.; Scarfì, S.; Mussino, F.; Salis, A.; Damonte, G.; Benatti, U.; Giovine, M. Pichia pastoris production of a prolyl 4-hydroxylase derived from Chondrosia reniformis sponge: A new biotechnological tool for the recombinant production of marine collagen. J. Biotechnol. 2015, 208, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Vuorela, A.; Myllyharju, J.; Nissi, R.; Pihlajaniemi, T.; Kivirikko, K.I. Assembly of human prolyl 4-hydroxylase and type III collagen in the yeast pichia pastoris: Formation of a stable enzyme tetramer requires coexpression with collagen and assembly of a stable collagen requires coexpression with prolyl 4-hydroxylase. EMBO J. 1997, 16, 6702–6712. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, L.N.; Zhu, C.H.; Fan, D.D.; Ma, X.X.; Mi, Y.; Xing, J.Y. Co-expression of recombinant human prolyl with human collagen α1 (III) chains in two yeast systems. Lett. Appl. Microbiol. 2015, 61, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Báez, J.; Olsen, D.; Polarek, J.W. Recombinant microbial systems for the production of human collagen and gelatin. Appl. Microbiol. Biotechnol. 2005, 69, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Yoshizumi, A.; Danon, S.J.; Glattauer, V.; Prokopenko, O.; Mirochnitchenko, O.; Yu, Z.; Inouye, M.; Werkmeister, J.A.; Brodsky, B.; et al. A Streptococcus pyogenes derived collagen-like protein as a non-cytotoxic and non-immunogenic cross-linkable biomaterial. Biomaterials 2010, 31, 2755–2761. [Google Scholar] [CrossRef] [PubMed]
- Nowack, H.; Hahn, E.; Timpl, R. Specificity of the antibody response in inbred mice to bovine type I and type II collagen. Immunology 1975, 29, 621–628. [Google Scholar] [PubMed]
- Kim, H.Y.; Kim, W.U.; Cho, M.L.; Lee, S.K.; Youn, J.; Kim, S.I.; Yoo, W.H.; Park, J.H.; Min, J.K.; Lee, S.H.; et al. Enhanced T cell proliferative response to type II collagen and synthetic peptide CII (255–274) in patients with rheumatoid arthritis. Arthritis Rheum. 1999, 42, 2085–2093. [Google Scholar] [CrossRef]
- Davison, P.F.; Levine, L.; Drake, M.P.; Rubin, A.; Bump, S. The serologic specificity of tropocollagen telopeptides. J. Exp. Med. 1967, 126, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Bentkover, S. The biology of facial fillers. Facial Plast. Surg. 2009, 25, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Stoichevska, V.; Howell, L.; Madsen, S.; Werkmeister, J.A.; Dumsday, G.J.; Ramshaw, J.A.M. Preparation and characterization of monomers to tetramers of a collagen-like domain from Streptococcus pyogenes. Bioengineered 2014, 5, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Parmar, P.A.; St-Pierre, J.-P.; Chow, L.W.; Puetzer, J.L.; Stoichevska, V.; Peng, Y.Y.; Werkmeister, J.A.; Ramshaw, J.A.M.; Stevens, M.M. Harnessing the versatility of bacterial collagen to improve the chondrogenic potential of porous collagen scaffolds. Adv. Healthc. Mater. 2016, 5, 1656–1666. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Stoichevska, V.; Vashi, A.; Howell, L.; Fehr, F.; Dumsday, G.J.; Werkmeister, J.A.; Ramshaw, J.A.M. Non-animal collagens as new options for cosmetic formulation. Int. J. Cosmet. Sci. 2015, 37, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Glattauer, V.; Ramshaw, J.A.M. Stabilisation of collagen sponges by glutaraldehyde vapour crosslinking. Int. J. Biomater. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Golser, A.; Roeber, M.; Boerner, H.; Scheibel, T. Engineered collagen—A redox switchable framework for tunable assembly and fabrication of biocompatible surfaces. ACS Biomater. Sci. Eng. 2017. [Google Scholar] [CrossRef]
- Hamaia, S.; Farndale, R.W. Integrin recognition motifs in the human collagens. Adv. Exp. Med. Biol. 2014, 819, 127–142. [Google Scholar] [PubMed]
- Woltersdorf, C.; Bonk, M.; Leitinger, B.; Huhtala, M.; Käpylä, J.; Heino, J.; Gil Girol, C.; Niland, S.; Eble, J.A.; Bruckner, P.; et al. The binding capacity of α1β1-, α2β1- and α10β1-integrins depends on non-collagenous surface macromolecules rather than the collagens in cartilage fibrils. Matrix Biol. J. Int. Soc. Matrix Biol. 2017, 63, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Zeltz, C.; Orgel, J.; Gullberg, D. Molecular composition and function of integrin-based collagen glues-introducing COLINBRIs. Biochim. Biophys. Acta 2014, 1840, 2533–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeltz, C.; Gullberg, D. The integrin-collagen connection—A glue for tissue repair? J. Cell Sci. 2016, 129, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Werten, M.W.T.; Teles, H.; Moers, A.P.H.A.; Wolbert, E.J.H.; Sprakel, J.; Eggink, G.; de Wolf, F.A. Precision gels from collagen-inspired triblock copolymers. Biomacromolecules 2009, 10, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.I.F.; Teles, H.; Moers, A.P.H.A.; Eggink, G.; de Wolf, F.A.; Werten, M.W.T. Secreted production of collagen-inspired gel-forming polymers with high thermal stability in Pichia pastoris. Biotechnol. Bioeng. 2011, 108, 2517–2525. [Google Scholar] [CrossRef] [PubMed]
- Beun, L.H.; Storm, I.M.; Werten, M.W.T.; de Wolf, F.A.; Cohen Stuart, M.A.; de Vries, R. From micelles to fibers: balancing self-assembling and random coiling domains in ph-responsive silk-collagen-like protein-based polymers. Biomacromolecules 2014, 15, 3349–3357. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.H.; Qin, X.X.; Coyne, K.J. The peculiar collagens of mussel byssus. Matrix Biol. J. Int. Soc. Matrix Biol. 1998, 17, 93–106. [Google Scholar] [CrossRef]
- Qin, X.-X.; Waite, J.H. A potential mediator of collagenous block copolymer gradients in mussel byssal threads. Proc. Natl. Acad. Sci. USA 1998, 95, 10517–10522. [Google Scholar] [CrossRef] [PubMed]
- Coyne, K.J.; Qin, X.X.; Waite, J.H. Extensible collagen in mussel byssus: A natural block copolymer. Science 1997, 277, 1830–1832. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.M.; Vaccaro, E.; Waite, J.H. A molecular, morphometric and mechanical comparison of the structural elements of byssus from Mytilus edulis and Mytilus galloprovincialis. J. Exp. Biol. 2002, 205, 1807–1817. [Google Scholar] [PubMed]
- Schmidt, S.; Reinecke, A.; Wojcik, F.; Pussak, D.; Hartmann, L.; Harrington, M.J. Metal-mediated molecular self-healing in histidine-rich mussel peptides. Biomacromolecules 2014, 15, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Ramalingam, K. Histochemical identification of dopa, dopamine and catechol in the phenol gland and the mode of tanning of byssus threads of Mytilus edulis. Acta Histochem. 1972, 42, 87–94. [Google Scholar] [PubMed]
- Waite, J.H. The phylogeny and chemical diversity of quinone-tanned glues and varnishes. Comp. Biochem. Physiol. Part B 1990, 97, 19–29. [Google Scholar] [CrossRef]
- Golser, A.V.; Scheibel, T. Biotechnological production of the mussel byssus derived collagen preColD. RSC Adv. 2017, 7, 38273–38278. [Google Scholar] [CrossRef]
- Heim, M.; Elsner, M.B.; Scheibel, T. Lipid-specific β-sheet formation in a mussel byssus protein domain. Biomacromolecules 2013, 14, 3238–3245. [Google Scholar] [CrossRef] [PubMed]
- Hagenau, A.; Suhre, M.H.; Scheibel, T.R. Nature as a blueprint for polymer material concepts: Protein fiber-reinforced composites as holdfasts of mussels. Prog. Polym. Sci. 2014, 39, 1564–1583. [Google Scholar] [CrossRef]
- Bairati, A.; Vitellaro Zuccarello, L. The ultrastructure of the byssal apparatus of Mytilus galloprovincialis: IV. Observations by transmission electron microscopy. Cell Tissue Res. 1976, 166. [Google Scholar] [CrossRef]
- Zuccarello, L.V. The collagen gland of mytilus galloprovincialis: An ultrastructural and cytochemical study on secretory granules. J. Ultrastruct. Res. 1980, 73, 135–147. [Google Scholar] [CrossRef]
- Suhre, M.H.; Scheibel, T. Structural diversity of a collagen-binding matrix protein from the byssus of blue mussels upon refolding. J. Struct. Biol. 2014, 186, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Suhre, M.H.; Gertz, M.; Steegborn, C.; Scheibel, T. Structural and functional features of a collagen-binding matrix protein from the mussel byssus. Nat. Commun. 2014, 5, 3392. [Google Scholar] [CrossRef] [PubMed]
- Hagenau, A.; Papadopoulos, P.; Kremer, F.; Scheibel, T. Mussel collagen molecules with silk-like domains as load-bearing elements in distal byssal threads. J. Struct. Biol. 2011, 175, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Koski, M.K.; Anantharajan, J.; Kursula, P.; Dhavala, P.; Murthy, A.V.; Bergmann, U.; Myllyharju, J.; Wierenga, R.K. Assembly of the elongated collagen prolyl 4-hydroxylase α2β2 heterotetramer around a central α2 dimer. Biochem. J. 2017, 474, 751–769. [Google Scholar] [CrossRef] [PubMed]
- Marumo, K.; Waite, J.H. Prolyl 4-hydroxylase in the foot of the marine mussel Mytilus edulis L.: purification and characterization. J. Exp. Zool. 1987, 244, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Yu, J.; Broomell, C.; Israelachvili, J.N.; Waite, J.H. Hydrophobic enhancement of dopa-mediated adhesion in a mussel foot protein. J. Am. Chem. Soc. 2013, 135, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Montroni, D.; Valle, F.; Rapino, S.; Fermani, S.; Calvaresi, M.; Harrington, M.J.; Falini, G. Functional biocompatible matrices from mussel byssus waste. ACS Biomater. Sci. Eng. 2017. [Google Scholar] [CrossRef]
- Butt, T.R.; Edavettal, S.C.; Hall, J.P.; Mattern, M.R. SUMO fusion technology for difficult-to-express proteins. Protein Exp. Purif. 2005, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golser, A.V.; Scheibel, T. Routes towards Novel Collagen-Like Biomaterials. Fibers 2018, 6, 21. https://doi.org/10.3390/fib6020021
Golser AV, Scheibel T. Routes towards Novel Collagen-Like Biomaterials. Fibers. 2018; 6(2):21. https://doi.org/10.3390/fib6020021
Chicago/Turabian StyleGolser, Adrian V., and Thomas Scheibel. 2018. "Routes towards Novel Collagen-Like Biomaterials" Fibers 6, no. 2: 21. https://doi.org/10.3390/fib6020021
APA StyleGolser, A. V., & Scheibel, T. (2018). Routes towards Novel Collagen-Like Biomaterials. Fibers, 6(2), 21. https://doi.org/10.3390/fib6020021