The Vascular Effects of Isolated Isoflavones—A Focus on the Determinants of Blood Pressure Regulation


Abstract
:Simple Summary
Abstract
1. Introduction
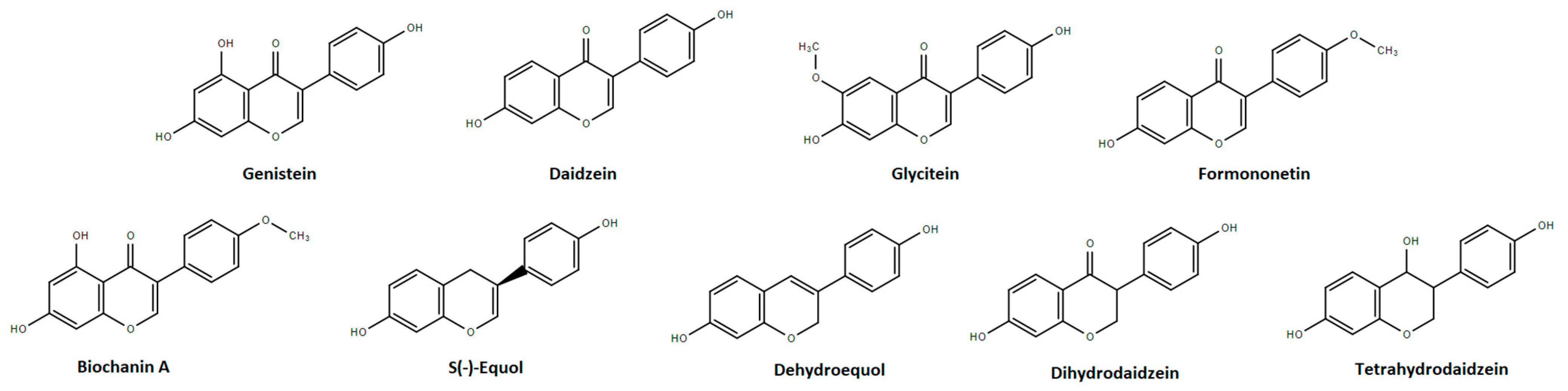
2. Chemical Characterization, Metabolism, and Safety
3. Vasorelaxant Effects of Isoflavones In Vitro
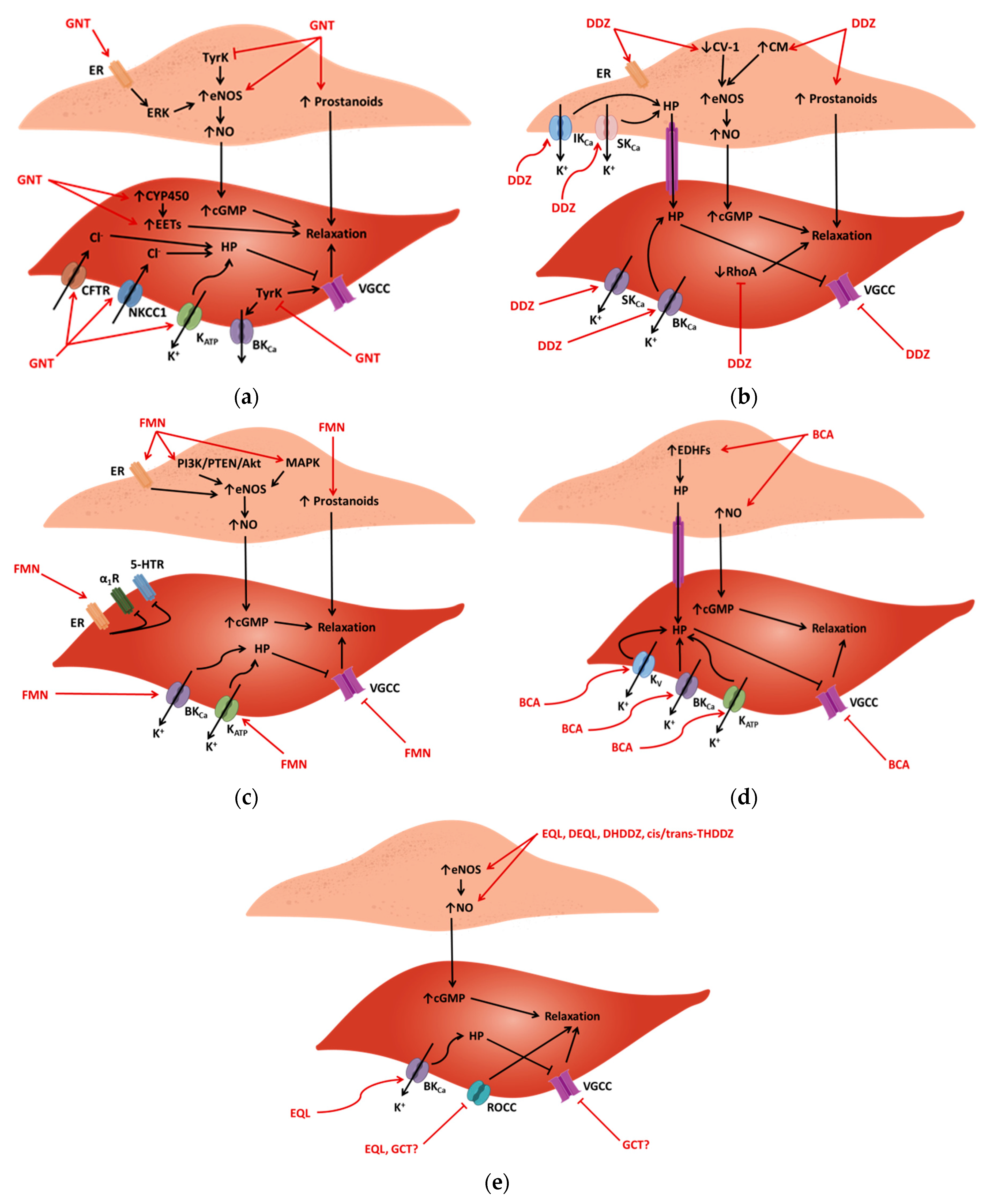
3.1. Endothelium-Dependent Vasorelaxation
3.2. Endothelium-Independent Vasorelaxation—The Role of Calcium Channels
3.3. Endothelium-Independent Vasorelaxation—The Role of Potassium Channels
3.4. Endothelium-Independent Vasorelaxation—The Role of Chloride Channels
3.5. Potentiation of the Protein Kinase A Pathway
3.6. Role of Tyrosine Kinase Inhibition
3.7. Inhibition of Rho-Kinase
3.8. Activity on Estrogen and Epidermal Growth Factor Receptors
3.9. Effect of Adrenergic Receptors

{kind=link}
{kind=link}
| Authors | Compound Concentration | Species and Strain | Type of Vessel | Main Results |
|---|---|---|---|---|
| Laniyonu et al. (1994) [228] | 1–15 μM | Male Sprague-Dawley rats | Aorta | Relaxation of pervanadate-induced but not of KCl-induced contraction |
| Moritoki et al. (1995) [211] | 100 μM | Rat (undisclosed strain) | Aorta | Prevention of LPS-primed, L-arginine-mediated vasorelaxation |
| Filipeanu et al. (1995) [153] | 10−6–10−3.5 M | Male Wistar rats | Aorta | Relaxation of PE-preconstricted vessels |
| Herrera et al. (1996) [229] | 10−6–10−3 M | Wistar rats (both genders) | Aorta | Relaxation of NE-, KCl-, phorbol 12-myristate-13-acetate-preconstricted vessels, responses found to be independent of tyrosine kinase inhibition |
| Watts et al. (1996) [147] | 5 × 10−6 M | Male Sprague-Dawley rats | Carotid artery and aorta (denuded) | Relaxation of 5-HT-preconstricted denuded carotid artery but absence of relaxation of phorbol-12,13-dibutyrate or KCl-preconstricted vessels |
| Duarte et al. (1997) [209] | Genistein (10 mg/kg i.p.) | Male Wistar rats | Aorta | Inhibition of LPS-mediated hyporresponsiveness to NE and inhibition of nitrite accumulation without affecting NOS. |
| Satake et al. (1999) [198] | 3 × 10−7, 10−6, 10−5 M | Male Wistar rats | Aorta | Relaxation of PE-preconstricted vessels. Potentiation of isoproterenol, forskolin and dibutyryl cAMP-induced vasorelaxation Isoproterenol-mediated vasorelaxation was inhibited by α-naphthoflavone and by 8-methoxypsoralen; the potentiation of forskolin-mediated vasorelaxation was inhibited by theophylline, iberiotoxin and quinacrine |
| Squadrito et al. (2000) [156] | 0.2 mg/kg/day administered subcutaneously for 4 weeks | Ovariectomized mature female Sprague-Dawley rats | Aorta | Ovariectomy decreased Ach-induced vasorelaxation but did not affect SNP response. Genistein restored endothelial-dependent vasorelaxation as well as Nω-L-arginine (L-NMA)-induced contraction. |
| Mishra et al. (2000) [149] | 1–100 μM | Male Wistar rats | Aorta and pulmonary arteries | Relaxation of PE and KCl-preconstricted vessels, attenuated by endothelium denudation and L-NAME but not by indomethacin. |
| Suenaga et al. (2002) [217] | 3 × 10−6, 10−5, 3 × 10−5 M | Male Wistar rats | Aorta | Prevention of lysophosphatidylcholine-induced contraction via tyrosine kinase activation. |
| Valero et al. (2006) [197] | 1–100 μM | Male Wistar rats | Aorta (denuded) | Relaxation, attenuated by diphenylamine-2-carboxylic acid, glibenclamide, H-89 and bumetanide. |
| Vera et al. (2007) [230] | 10 mg/kg/day administered by gavage for 5 weeks | Female SHRs | Aorta | Potentiation of Ach-mediated vasorelaxation. Attenuation of angiotensin II-mediated vasoconstriction. No effect on NE-mediated vasoconstriction. |
| Baluchnejadmojarad et al. (2008) [231] | 1 mg/kg/day administered intraperitoneally for 4 weeks | Male albino Wistar rats with streptozotocin-induced diabetes and controls | Aorta | Potentiation of Ach-mediated vasorelaxation, partially attenuated by L-NAME and indomethacin. Attenuation of NE and KCl-mediated contraction of aortic rings, with the endothelium removal abolishing the difference between treated and untreated diabetic rats |
| Galan-Martinez (2008) [151] | 1 to 300 μM | Male adult Wistar rats | Aorta (denuded) | Vasorelaxation of KCl-preconstricted vessels. |
| Je et al. (2009) [152] | 0.03and 0.1 mM | Male Sprague-Dawley rats | Aorta | Relaxation of PE-, phorbol ester-, KCl-, fluoride-, and TXA2-preconstricted vessels, regardless of endothelial function. Vasorelaxation was not inhibited by iberiotoxin. |
| Liu et al. (2007) [214] | 10−6, 10−5, or 10−4 M | Cows | Pulmonary Arteries and Veins | Relaxation of KCl-, caffeine- and U46619-preconstricted vessels. Genistein prevented U46619-induced tyrosine phosphorylation of a Rho-GEF, a necessary component for Rho kinase action. This leads to the notion that genistein might have inhibited tyrosine kinase. |
| Nevala et al. (1998) [148] | 10−6–10−4 M | Female and male Wistar rats were used | Mesenteric arteries (intact and denuded) | Relaxation of NE-, KCl- and CaCl2-preconstricted vessels, which was unaffected by L-NAME or indomethacin |
| Honore et al. (1997) [232] | 140 mg administered intravenously | Young adult rhesus monkeys | Coronary arteries | Vasorelaxation in monkeys receiving an isoflavone-deprived diet. |
| Figtree et al. (2000) [38] | 10, 20 and 40 mM | Adult New Zealand white rabbits (both genders) | Coronary arteries | Relaxation of KCl-preconstricted vessels, either intact or denuded. The response was unaffected by L-NAME, indomethacin, glibenclamide, barium, methylene blue or ICI 182,780. |
| Lee et al. (2003) [206] | 0.1–100 μM | Pigs | Coronary arteries | Relaxation of U46619-preconstricted vessels, not affected by bradykinin or A23187 but potentiated by SNP and cromakalim. Endothelium denudation and tyrphostin 23 did not affect SNP-induced relaxation. |
| Kimoto et al. (2005) [207] | 10−6 M | Pigs | Coronary arteries (denuded) | Mexiletine and lidocaine partially abolished the vasorelaxant response to levcromakalim (i.e. KATP channel activator). Genistein restored the levcromakalin-induced vasorelaxation in the presence of mexiletine but not of lidocaine. |
| Ng et al. (2008) [183] | 10−6.5–10−4 M | Pigs | Coronary arteries | Increase in the activity of protein kinase A at high concentration (10−4.5 M). At a lower concentration (10−5.5 M) it failed to increase PKA activity, unless together with forskolin. SQ22536 (i.e., adenylyl cyclase inhibitor) blocked the genistein-mediated potentiation of PKA, unlike NF 449 (i.e. P2X1 receptor antagonist). |
| Pinna et al. (2019) [157] | 1 nM–0.1 mM | Human subjects | Umbilical veins | Relaxation of 5-HT-preconstricted intact vessels, attenuated by L-NAME, and completely abolished by L-NAME and glibenclamide. |
| Kitazono et al. (1998) [202] | 10−6 or 3×10−6 M | Male Sprague-Dawley rats | Basilar artery | Attenuation of Ach and bradykinin-mediated vasorelaxation, whereas SNP-mediated relaxation was not affected. |
| Kitayama et al. (2002) [218] | 1 mg/kg chow for 2 months | Male SHR | Basilar artery | Potentiation of Ach- and Y-27632-mediated vasorelaxation in SHR. No attenuation of the increased Rho kinase activity. |
| Authors | Compound Concentration | Species and Strain | Type of Vessel | Main Results |
|---|---|---|---|---|
| Mishra et al. (2000) [149] | 10 and 100 μM | Adult male Wistar rats | Aorta | Relaxation of PE-preconstricted vessels, attenuated by endothelium denudation and by L-NAME. |
| Woodman et al. (2004) [188] | 0.2 mg/kg/day administered subcutaneously for 7 days | Male Sprague–Dawley rats | Aorta | Potentiation of Ach-induced vasorelaxation, attenuated by L-NOLA, PPOH, barium, and oubain |
| Ajay et al. (2003) [150] | 0.3 mM | Male Wistar-Kyoto rats | Aorta (intact) | Relaxation of PE and KCl-preconstricted vessels, reduced by L-NAME and indomethacin |
| Je et al. (2009) [152] | 0.01, 0.03 and 0.1 mM | Male Sprague-Dawley rats | Aorta | Relaxation of PE-, KCl-, fluoride-, and phorbol ester-preconstricted vessels |
| Sharma et al. (2012) [162] | 0.2, 0.4 or 0.6 mg/kg/day administered subcutaneously for 7 days | Female Sprague-Dawley ovariectomized rats | Aorta | Potentiation of Ach-, but not of SNP-induced, vasorelaxation. Increased expression of eNOS, calmodulin, and decreased expression of caveolin-1, which prevented ovariectomy-induced vascular dysfunction. |
| Roghani et al. (2013) [158] | 5 or 10 mg/kg administrated by gavage for 7 weeks | Healthy and streptozotocin-induced diabetic Male Wistar rats | Aorta | Relaxation of PE-preconstricted vessels, abolished by endothelium denudation. In treated animals, Ach-induced vasorelaxation was also higher, but L-NAME and indomethacin attenuated this response. |
| Prawez et al. (2015) [159] | 0.5 mg/kg administered subcutaneously for 6 weeks | L-NAME hypertensive male Wistar-rats | Aorta | Relaxation of KCl-preconstricted vessels. Potentiation of Ach-induced vasorelaxation. Decreased potency of SNP-induced vasorelaxation. |
| Sobey et al. (2004) [161] | 0.2 mg/kg/day administered subcutaneously for 7 days | Male Sprague-Dawley rats | Carotid and basilar arteries | Potentiation of Ach-induced vasorelaxation. Potentiation of L-NNA-induced contraction. Expression of caveolin-1 decreased, expression of calmodulin increased and expression of eNOS was unaffected. |
| Jackman et al. (2007) [99] | 10−7–10−3 M | Adult Sprague–Dawley rats (both genders) | Carotid and basilar arteries | Relaxation of carotid arteries and vasodilation (in vivo) of basilar arteries. |
| Zhang et al. (2010) [189] | 10−7–10−4 M | Male Sprague–Dawley rats | Basilar artery | Vasorelaxation, inhibited by paxilline, but enhanced by NS1619 |
| Torregrosa et al. (2003) [155] | 10−7–10−4 M | New Zealand White male rabbits | Basilar arteries | Relaxation of KCl or UTP-precontracted vessels, unaffected by endothelial denudation or by L-NAME, ODQ, or NS2028 |
| Nevala et al. (1998) [148] | 10−6–10−4 M | Wistar rats (both genders) | Mesenteric arteries | Relaxation of NE, KCl or CaCl2 precontracted vessels, unaffected by endothelium denudation, indomethacin or L-NAME |
| Nevala et al. (2001) [190] | 10–100 mM | Female Wistar rats | Mesenteric arteries (denuded) | Relaxation of NE- and KCl-preconstricted vessels, attenuated by iberiotoxin, charybdotoxin, and apamin |
| Authors | Compound Concentration | Species and Strain | Type of Vessel | Main Results |
|---|---|---|---|---|
| Wu et al. (2010) [167] | 10 μM, 100 μM, and 1 mM | Male Sprague-Dawley rats | Aorta | Relaxation of intact vessels, suppressed by L-NAME and methylene blue but not by 1400 W or propranolol. In endothelium-denuded vessels, vasorelaxation was attenuated by methylene blue, glibenclamide, or iberiotoxin. Enhancement of eNOS expression and activity. |
| Zhao et al. (2012) [184] | 10–100 μM | Male Sprague-Dawley rats | Aorta | Relaxation of KCl- and NE-preconstricted vessels |
| Li et al. (2018) [169] | 10−8–10−3 M | Sprague-Dawley rats | Aorta (intact and denuded) | Relaxation of KCl-preconstricted vessels, suppressed by endothelium denudation and by L-NAME |
| Sun et al. (2011) [154] | 1–300 μM | Sprague-Dawley rats | Mesenteric, renal, basilar, coronary and aortic arteries (intact and denuded) | Relaxation of PE- or U46619-preconstricted vessels, attenuated by endothelium denudation |
| Male SHRs | Mesenteric arteries | Relaxation of PE- or U46619-preconstricted arteries, attenuated by endothelium denudation and by L-NAME but unaffected by indomethacin or glibenclamide | ||
| Sun et al. (2016) [170] | 1–10 μM | Male Sprague-Dawley rats | Mesenteric arteries | Vasorelaxation, attenuated by ICI 182780. Expression of eNOS increased via ERK and JNK activation. |
| Tseng et al. (2016) [168] | 1 nM–100 μM | Male Sprague-Dawleyrats | Mesenteric arteries | Relaxation of intact vessels, suppressed by L-NAME and glibenclamide |
| Sun et al. (2013) [225] | 50 mg/kg administered orally for 8 weeks | Male SHRs | Mesenteric arteries | Attenuation of NE or 5-HT-induced vasoconstriction. Potentiation of Ach-induced vasodilation. Expression of alpha-1 adrenergic and 5-HT2A/1B receptors in VSM cells decreased. |
| Authors | Compound Concentration | Species and Strain | Type of Vessel | Main Results |
|---|---|---|---|---|
| Choi et al. (2014) [176] | 10−7–10−4 M | Male Sprague-Dawley rats subjected to 2K1C-induced renovascular hypertension | Aorta | Endothelium-dependent vasorelaxation was attenuated by endothelium denudation, by glibenclamide, TEA and by 4-aminopyridine |
| Wang et al. (2005) [185] | 10−9–10−4 M | Sprague-Dawley rats | Aorta | Relaxation of PE-, KCl-, and CaCl2-preconstricted vessels, intact and endothelium-denuded, attenuated by TEA and glibenclamide but not by 4-aminopyridine |
| Wang et al. (2006) [175] | 10−7–3×10−4 M | SHRs | Aorta | Vasorelaxation, attenuated by endothelium denudation, by glibenclamide and 4-aminopyridine but not by L-NAME or indomethacin |
| Migko et al. (2020) [177] | 10−7–10−3 M | Wistar Han rats | Aorta | Relaxation of NE-preconstricted vessels. |
| 3×10−6–3×10−5 M | Pigs | Coronary arteries | Relaxation of KCl-, CaCl2-, 5-HT-, and U46619-preconstricted vessels. Endothelium denudation did not affect relaxation of U46619-preconstricted vessels. Potentiation of vasorelaxation by SNP but not by forskolin. | |
| Torregrosa et al. (2003) [155] | 10−6–10−4 M | New Zealand White male rabbits | Basilar artery | Vasorelaxation, unaffected by endothelium denudation, by L-NAME, indomethacin, ODQ, or NS2028 |
| Figtree et al. (2000) [38] | 3, 10 and 30 μM | Adult male or non-pregnant female New Zealand white rabbits | Coronary arteries | Relaxation, independent of gender, of intact and denuded vessels |
| Authors | Compound Concentration | Species and Strain | Type of Vessel | Main Results |
|---|---|---|---|---|
| Chin-Dusting et al. (2001) [178] | Dihydrodaidzein, cis-, and trans-tetrahydrodaidzein, dehydroequol (1 μg/mL) | Male Sprague-Dawley rats | Aorta | Vasorelaxation, inhibited by endothelium denudation, by L-NOLA, ODQ, or KCl |
| Joy et al. (2006) [179] | Equol (0.03–15 μM) | Sprague-Dawley rats | Aorta | Vasorelaxation by stimulation of NO release, a response that is unaffected by ICI 182,780 or pertussis toxin |
| Ohkura et al. (2015) [180] | Equol (200 mg/day for 5 weeks via osmotic pump) | Isoflavone-deficient ovariectomized Sprague-Dawley rats | Carotid arteries | Restoration of Ach-mediated vasorelaxation, which was suppressed by Nω-monomethyl-L-arginine acetate (L-NMMA) |
| Kim et al. (2015) [181] | Equol (10−11–10−6 M) | Human subjects | Uterine arteries | Relaxation of PE-, but not of KCl-preconstricted vessels |
| Cheng et al. (2010) [182] | Equol (0.1, 1, and 10 μM) | Pigs | Pulmonary arteries | Restoration of ritonavir-induced reduction in eNOS expression. Restoration of bradykinin-mediated vasorelaxation. |
4. Effect on Blood Pressure and Blood Flow
4.1. Effect on the Renin–Angiotensin–Aldosterone Axis
4.2. Effect on Baroreceptor Sensitivity
4.3. Effect on Catecholamine Synthesis
4.4. Possible Connection between Effects on Immunity and Blood Pressure
| Authors | Compound Concentration/Dosage and Duration of Treatment | Animal Species and Strain | Blood Pressure Measurement Technique | Main Results |
|---|---|---|---|---|
| Vera et al. (2007) [230] | Genistein (10 mg/kg/day) by gavage for 5 weeks | Ovariectomized SHRs | Tail-cuff plethysmography | Reduction of SBP after 5 weeks |
| Prawez et al. (2015) [159] | Daidzein (0.5 mg/kg) administered subcutaneously for 6 weeks | L-NAME-induced hypertensive rats | Invasive pressure transducer | Reduction of MBP in comparison to untreated animals |
| Cao et al. (2006) [237] | Daidzein sulfates (20 mg/kg and 40 mg/kg) with a single or multiple (1/day for 2 weeks) oral administrations | SHRs | Tail-cuff plethysmography | Blood pressure reduction with single or multiple administrations. Daidzein decreases blood pressure in multidose (more slowly than for sulfates) but not in single dose administrations |
| Liu et al. (2015) [98] | Daidzein (63 mg) taken orally for 6 months | Pre-hypertensive and untreated hypertensive post-menopausal subjects | Portable device, calibrated by a mercury sphygmomanometer | No change in blood pressure. No improvement in flow-mediated dilation was observed. |
| Sachdeva et al. (2016) [238] | Biochanin A (10 mg/kg, p.o. for 1 week) | Hypertensive ovariectomized rats | Tail-cuff plethysmography | Reduction of SBP, DBP and MBP, blunted by L-NAME |
| Sun et al. (2013) [225] | Formononetin (50 mg/kg/day for 8 weeks) | SHRs | Tail-cuff plethysmography | Blood pressure reduction and endothelium-dependent vasodilation |
| Palanisamy and Venkataraman (2013) [253] | Genistein (1 mg/kg/day for 44 days) by oral administration | Fructose-fed hypertensive male Wistar rats | Tail-cuff plethysmography | No significant change in blood pressure |
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, X.; Duncan, A.M.; Merz, B.E.; Kurzer, M.S. Effects of soy isoflavones on estrogen and phytoestrogen metabolism in premenopausal women. Cancer Epidemiol. Biomarkers Prev. 1998, 7, 1101–1108. [Google Scholar]
- Pino, A.M.; Valladares, L.E.; Palma, M.A.; Mancilla, A.M.; Yáñez, M.; Albala, C. Dietary isoflavones affect sex hormone-binding globulin levels in postmenopausal women. J. Clin. Endocrinol. Metab. 2000, 85, 2797–2800. [Google Scholar] [CrossRef] [PubMed]
- Dakora, F.D.; Phillips, D.A. Diverse functions of isoflavonoids in legumes transcend anti-microbial definitions of phytoalexins. Physiol. Mol. Plant Pathol. 1996, 49, 1–20. [Google Scholar] [CrossRef]
- Bellou, S.; Karali, E.; Bagli, E.; Al-Maharik, N.; Morbidelli, L.; Ziche, M.; Adlercreutz, H.; Murphy, C.; Fotsis, T. The isoflavone metabolite 6-methoxyequol inhibits angiogenesis and suppresses tumor growth. Mol. Cancer 2012, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.C.; Chen, Y.M.; Ho, S.S.S.; Woo, J.L.F. Soy isoflavone supplementation and fasting serum glucose and lipid profile among postmenopausal Chinese women: A double-blind, randomized, placebo-controlled trial. Menopause 2007, 14, 905–912. [Google Scholar] [CrossRef]
- Wei, P.; Liu, M.; Chen, Y.; Chen, D.C. Systematic review of soy isoflavone supplements on osteoporosis in women. Asian Pac. J. Trop. Med. 2012, 5, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, F.H.; Li, Y. Soy Isoflavones and Cancer Prevention. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef]
- Clarkson, T.B. Soy, Soy Phytoestrogens and Cardiovascular Disease. J. Nutr. 2002, 132, 566–569. [Google Scholar] [CrossRef]
- Matthan, N.R.; Jalbert, S.M.; Ausman, L.M.; Kuvin, J.T.; Karas, R.H.; Lichtenstein, A.H. Effect of soy protein from differently processed products on cardiovascular disease risk factors and vascular endothelial function in hypercholesterolemic subjects. Am. J. Clin. Nutr. 2007, 85, 960–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teede, H.J.; McGrath, B.P.; DeSilva, L.; Cehun, M.; Fassoulakis, A.; Nestel, P.J. Isoflavones reduce arterial stiffness: A placebo-controlled study in men and postmenopausal women. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestel, P.J.; Pomeroy, S.; Sally, K.; Komesaroff, P.; Behrsing, J.; Cameron, J.D.; West, L. Isoflavones from red clover improve systemic arterial compliance but not plasma lipids in menopausal women. J. Clin. Endocrinol. Metab. 1999, 84, 895–898. [Google Scholar] [CrossRef]
- Nestel, P.J.; Yamashita, T.; Sasahara, T.; Pomeroy, S.; Dart, A.; Komesaroff, P.; Owen, A.; Abbey, M. Soy isoflavones improve systemic arterial compliance but not plasma lipids in menopausal and perimenopausal women. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 3392–3398. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Sumner, L.W. Legume natural products: Understanding and manipulating complex pathways for human and animal health. Plant Physiol. 2003, 131, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.A.; Frankel, F.; Takahashr, H.; Vance, N.; Stiegerwald, C.; Edelstein, S. Collected literature on isoflavones and chronic diseases. Cogent Food Agric. 2016, 2, 1135861. [Google Scholar] [CrossRef]
- Heinonen, S.M.; Wähälä, K.; Adlercreutz, H. Identification of urinary metabolites of the red clover isoflavones formononetin and biochanin A in human subjects. J. Agric. Food Chem. 2004, 52, 6802–6809. [Google Scholar] [CrossRef]
- Lee, S.J.; Yan, W.; Ahn, J.K.; Chung, I.M. Effects of year, site, genotype and their interactions on various soybean isoflavones. Field Crop. Res. 2003, 81, 181–192. [Google Scholar] [CrossRef]
- Cassidy, A.; Brown, J.E.; Hawdon, A.; Faughnan, M.S.; King, L.J.; Millward, J.; Zimmer-Nechemias, L.; Wolfe, B.; Setchelly, K.D.R. Factors affecting the bioavailability of soy isoflavones in humans after ingestion of physiologically relevant levels from different soy foods. J. Nutr. 2006, 136, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.A.; Custer, L.J.; Wang, W.; Shi, C.Y. HPLC analysis of isoflavonoids and other phenolic agents from foods and from human fluids. Proc. Soc. Exp. Biol. Med. 1998, 217, 263–273. [Google Scholar] [CrossRef]
- King, R.A.; Bursill, D.B. Plasma and urinary kinetics of the isoflavones daidzein and genistein after a single soy meal in humans. Am. J. Clin. Nutr. 1998, 67, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Yamaguchi, M.; Sobue, T.; Takahashi, T.; Miura, T.; Arai, Y.; Mazur, W.; Wähälä, K.; Adlercreutz, H. Pharmacokinetics of soybean isoflavones in plasma, urine and feces of men after ingestion of 60 g baked soybean powder (kinako). J. Nutr. 1998, 128, 1710–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Wang, H.J.; Murphy, P.A.; Cook, L.; Hendrich, S. Daidzein is a more bioavailable soymilk isoflavone than is genistein in adult women. J. Nutr. 1994, 124, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Chundi, V.; Challa, S.R.; Garikapati, D.R.; Juvva, G.; Jampani, A.; Pinnamaneni, S.H.; Venigalla, S. Biochanin-A attenuates neuropathic pain in diabetic rats. J. Ayurveda Integr. Med. 2016, 7, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harini, R.; Ezhumalai, M.; Pugalendi, K.V. Antihyperglycemic effect of biochanin A, a soy isoflavone, on streptozotocin-diabetic rats. Eur. J. Pharmacol. 2012, 676, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Chukwumah, Y.C.; Walker, L.T.; Verghese, M.; Ogutu, S. Effect of frequency and duration of ultrasonication on the extraction efficiency of selected isoflavones and trans-resveratrol from peanuts (Arachis hypogaea). Ultrason. Sonochem. 2009, 16, 293–299. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Yang, X.; Xia, Z. Effects of sodium selenite and germination on the sprouting of chickpeas (Cicer arietinum L.) and its content of selenium, formononetin and biochanin A in the sprouts. Biol. Trace Elem. Res. 2012, 146, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Khedgikar, V.; Gautam, J.; Kushwaha, P.; Kumar, A.; Nagar, G.K.; Dixit, P.; Chillara, R.; Voruganti, S.; Singh, S.P.; Uddin, W.; et al. A standardized phytopreparation from an Indian medicinal plant (Dalbergia sissoo) has antiresorptive and bone-forming effects on a postmenopausal osteoporosis model of rat. Menopause 2012, 19, 1336–1346. [Google Scholar] [CrossRef]
- Sartorelli, P.; Carvalho, C.S.; Reimão, J.Q.; Ferreira, M.J.P.; Tempone, A.G. Antiparasitic activity of biochanin A, an isolated isoflavone from fruits of Cassia fistula (Leguminosae). Parasitol. Res. 2009, 104, 311–314. [Google Scholar] [CrossRef]
- Ming, X.; Ding, M.; Zhai, B.; Xiao, L.; Piao, T.; Liu, M. Biochanin A inhibits lipopolysaccharide-induced inflammation in human umbilical vein endothelial cells. Life Sci. 2015, 136, 36–41. [Google Scholar] [CrossRef]
- Chen, J.; Ge, B.; Wang, Y.; Ye, Y.; Zeng, S.; Huang, Z. Biochanin a promotes proliferation that involves a feedback loop of microRNA-375 and estrogen receptor alpha in breast cancer cells. Cell. Physiol. Biochem. 2015, 35, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Howes, J.B.; Tran, D.; Brillante, D.; Howes, L.G. Effects of dietary supplementation with isoflavones from red clover on ambulatory blood pressure and endothelial function in postmenopausal type 2 diabetes. Diabetes Obes. Metab. 2003, 5, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Thorup, A.C.; Lambert, M.N.; Kahr, H.S.; Bjerre, M.; Jeppesen, P.B. Intake of novel red clover supplementation for 12 weeks improves bone status in healthy menopausal women. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, M.N.T.; Thorup, A.C.; Hansen, E.S.S.; Jeppesen, P.B. Combined Red Clover isoflavones and probiotics potently reduce menopausal vasomotor symptoms. PLoS ONE 2017, 12, e0176590. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.R.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol—A clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [CrossRef] [Green Version]
- Szeja, W.; Grynkiewicz, G.; Rusin, A. Isoflavones, their Glycosides and Glycoconjugates. Synthesis and Biological Activity. Curr. Org. Chem. 2016, 21, 218–235. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J. Nutr. 2001, 131, 1362–1375. [Google Scholar] [CrossRef] [Green Version]
- Figtree, G.A.; Griffiths, H.; Lu, Y.Q.; Webb, C.M.; MacLeod, K.; Collins, P. Plant-derived estrogens relax coronary arteries in vitro by a calcium antagonistic mechanism. J. Am. Coll. Cardiol. 2000, 35, 1977–1985. [Google Scholar] [CrossRef] [Green Version]
- Varinska, L.; Gal, P.; Mojzisova, G.; Mirossay, L.; Mojzis, J. Soy and breast cancer: Focus on angiogenesis. Int. J. Mol. Sci. 2015, 16, 11728–11749. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Cho, J.Y.; Kim, J.H.; Park, W.K.; Kim, D.K.; Rhyu, M.R. Extracts from Schizandra chinensis fruit activate estrogen receptors: A possible clue to its effects on nitric oxide-mediated vasorelaxation. Biol. Pharm. Bull. 2004, 27, 1066–1069. [Google Scholar] [CrossRef] [Green Version]
- Dan Ramdath, D.; Padhi, E.M.T.; Sarfaraz, S.; Renwick, S.; Duncan, A.M. Beyond the cholesterol-lowering effect of soy protein: A review of the effects of dietary soy and its constituents on risk factors for cardiovascular disease. Nutrients 2017, 9, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tham, D.M.; Gardner, C.D.; Haskell, W.L. Potential health benefits of dietary phytoestrogens: A review of the clinical, epidemiological, and mechanistic evidence. J. Clin. Endocrinol. Metab. 1998, 83, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Bingham, S.A.; Atkinson, C.; Liggins, J.; Bluck, L.; Coward, A.; August, R.I.; Revised, I.; Accepted, I.D.; December, I. Phyto-oestrogens: Where are we now? Clin. Nutr. 1998, 44, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adlercreutz, H.; Mousavi, Y.; Clark, J.; Höckerstedt, K.; Hämäläinen, E.; Wähälä, K.; Mäkelä, T.; Hase, T. Dietary phytoestrogens and cancer: In vitro and in vivo studies. J. Steroid Biochem. Mol. Biol. 1992, 41, 331–337. [Google Scholar] [CrossRef]
- Merritt, J.C. Therapeutic options: Hormone replacement therapy-soy therapy. J. Natl. Med. Assoc. 2001, 93, 288–292. [Google Scholar]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Cornu, K.A.L.; Ryder, J.J.; Hall, W.L.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials 1, 2. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef]
- Rosero Arenas, M.Á.; Rosero Arenas, E.; Portaceli Armiñana, M.A.; García García, M.Á. Utilidad de los fitoestrógenos en la reducción de la presión arterial. Revisión sistemática y metaanálisis. Aten. Primaria 2008, 40, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Taku, K.; Lin, N.; Cai, D.; Hu, J.; Zhao, X.; Zhang, Y.; Wang, P.; Melby, M.K.; Hooper, L.; Kurzer, M.S.; et al. Effects of soy isoflavone extract supplements on blood pressure in adult humans: Systematic review and meta-analysis of randomized placebo-controlled trials. J. Hypertens. 2010, 28, 1971–1982. [Google Scholar] [CrossRef]
- Dong, J.Y.; Tong, X.; Wu, Z.W.; Xun, P.C.; He, K.; Qin, L.Q. Effect of soya protein on blood pressure: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2011, 106, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.X.; Li, S.H.; Chen, J.Z.; Sun, K.; Wang, X.J.; Wang, X.G.; Hui, R.T. Effect of soy isoflavones on blood pressure: A meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 463–470. [Google Scholar] [CrossRef]
- Yan, Z.; Zhang, X.; Li, C.; Jiao, S.; Dong, W. Association between consumption of soy and risk of cardiovascular disease: A meta-analysis of observational studies. Eur. J. Prev. Cardiol. 2017, 24, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Namazi, N.; Saneei, P.; Larijani, B.; Esmaillzadeh, A. Soy product consumption and the risk of all-cause, cardiovascular and cancer mortality: A systematic review and meta-analysis of cohort studies. Food Funct. 2018, 9, 2576–2588. [Google Scholar] [CrossRef] [PubMed]
- Nachvak, S.M.; Moradi, S.; Anjom-shoae, J.; Rahmani, J.; Nasiri, M.; Maleki, V.; Sadeghi, O. Soy, Soy Isoflavones, and Protein Intake in Relation to Mortality from All Causes, Cancers, and Cardiovascular Diseases: A Systematic Review and Dose–Response Meta-Analysis of Prospective Cohort Studies. J. Acad. Nutr. Diet. 2019, 119, 1483–1500.e17. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wu, X.; Zhuang, W.; Xia, L.; Chen, Y.; Zhao, R.; Yi, M.; Wan, Q.; Du, L.; Zhou, Y. Soy and Isoflavone Consumption and Multiple Health Outcomes: Umbrella Review of Systematic Reviews and Meta-Analyses of Observational Studies and Randomized Trials in Humans. Mol. Nutr. Food Res. 2020, 64, 1–12. [Google Scholar] [CrossRef]
- Coward, L.; Smith, M.; Kirk, M.; Barnes, S. Chemical modification of isoflavones in soyfoods during cooking and processing. Am. J. Clin. Nutr. 1998, 68, 1486–1491. [Google Scholar] [CrossRef]
- Slavin, J.L.; Karr, S.C.; Hutchins, A.M.; Lampe, J.W. Influence of soybean processing, habitual diet, and soy dose on urinary isoflavonoid excretion. Am. J. Clin. Nutr. 1998, 68, 1492–1495. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tsuji, S.; Tonogai, Y. Determination of the levels of isoflavonoids in soybeans and soy-derived foods and estimation of isoflavonoids in the Japanese daily intake. J. AOAC Int. 2000, 83, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A bacterial metabolite from the daidzein. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Amaretti, A.; Roncaglia, L.; Leonardi, A.; Raimondi, S. Dietary isoflavones and intestinal microbiota: Metabolism and transformation into bioactive compounds. In Isoflavones: Biosynthesis, Occurrence and Health Effects; Thompson, M.J., Ed.; Nova Publisher: New York, NY, USA, 2010; pp. 137–161. [Google Scholar]
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; McLauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, S.I.; Suzuki, T. Isolation and characterization of a novel equol-producing bacterium from human feces. Biosci. Biotechnol. Biochem. 2008, 72, 2660–2666. [Google Scholar] [CrossRef] [Green Version]
- Rothwell, J.A.; Day, A.J.; Morgan, M.R.A. Experimental determination of octanol-water partition coefficients of quercetin and related flavonoids. J. Agric. Food Chem. 2005, 53, 4355–4360. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Ge, J.; Zhang, Y.P.; Yu, Y.; Zhang, X.Y. Solubility of genistein in water, methanol, ethanol, propan-2-ol, 1-butanol, and ethyl acetate from (280 to 333) K. J. Chem. Eng. Data 2010, 55, 5286–5288. [Google Scholar] [CrossRef]
- Kwon, S.H.; Kang, M.J.; Huh, J.S.; Ha, K.W.; Lee, J.R.; Lee, S.K.; Lee, B.S.; Han, I.H.; Lee, M.S.; Lee, M.W.; et al. Comparison of oral bioavailability of genistein and genistin in rats. Int. J. Pharm. 2007, 337, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Steensma, A.; Faassen-Peters, M.A.W.; Noteborn, H.P.J.M.; Rietjens, I.M.C.M. Bioavailability of genistein and its glycoside genistin as measured in the portal vein of freely moving unanesthetized rats. J. Agric. Food Chem. 2006, 54, 8006–8012. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, H.; Hu, M. Metabolism of flavonoids via enteric recycling: Role of intestinal disposition. J. Pharmacol. Exp. Ther. 2003, 304, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Enokizono, J.; Kusuhara, H.; Sugiyama, Y. Effect of breast cancer resistance protein (Bcrp/Abcg2) on the disposition of phytoestrogens. Mol. Pharmacol. 2007, 72, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, A.I.; Vallejo, F.; Barrera, B.; Merino, G.; Prieto, J.G.; Tomás-Barberán, F.; Espín, J.C. Bioavailability of the Glucuronide and Sulfate Conjugates of Genistein and Daidzein in Breast Cancer Resistance Protein 1 Knockout Mice Received. Drug Metab. Dispos. 2012, 39, 2008–2012. [Google Scholar] [CrossRef]
- Yang, Z.; Kulkarni, K.; Zhu, W.; Hu, M. Bioavailability and Pharmacokinetics of Genistein: Mechanistic Studies on its ADME. Anticancer Agents Med. Chem. 2012, 12, 1264–1280. [Google Scholar] [CrossRef]
- Coldham, N.G.; Sauer, M.J. Pharmacokinetics of [14C]genistein in the rat: Gender-related differences, potential mechanisms of biological action, and implications for human health. Toxicol. Appl. Pharmacol. 2000, 164, 206–215. [Google Scholar] [CrossRef]
- Heinonen, S.M.; Hoikkala, A.; Wähälä, K.; Adlercreutz, H. Metabolism of the soy isoflavones daidzein, genistein and glycitein in human subjects. Identification of new metabolites having an intact isoflavonoid skeleton. J. Steroid Biochem. Mol. Biol. 2003, 87, 285–299. [Google Scholar] [CrossRef]
- Liu, Y.A.N.; Hu, M. Absorption and metabolism of flavonoids in the caco-2 cell culture model and a perused rat intestinal model. Drug Metab. Dispos. 2002, 30, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coldham, N.G.; Darby, C.; Hows, M.; King, L.J.; Zhang, A.Q.; Sauer, M.J. Comparative metabolism of genistin by human and rat gut microflora: Detection and identification of the end-products of metabolism. Xenobiotica 2002, 32, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, K.; Furuta, T.; Yokokawa, A.; Ogura, K.; Hiratsuka, A.; Ishii, K. Plasma profiling of intact isoflavone metabolites by high-performance liquid chromatography and mass spectrometric identification of flavone glycosides daidzin and genistin in human plasma after administration of kinako. Drug Metab. Dispos. 2008, 36, 1485–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.J.W.; Anderson, K.E. Sex and long-term soy diets affect the metabolism and excretion of soy isoflavones in humans. Am. J. Clin. Nutr. 1998, 68, 1500–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Zhang, J.; Ye, J. Determination of puerarin, daidzein and rutin in Pueraria lobata (Wild.) Ohwi by capillary electrophoresis with electrochemical detection. J. Chromatogr. A 2001, 923, 255–262. [Google Scholar] [CrossRef]
- Murphy, P.A.; Song, T.; Buseman, G.; Barua, K.; Beecher, G.R.; Trainer, D.; Holden, J. Isoflavones in retail and institutional soy foods. J. Agric. Food Chem. 1999, 47, 2697–2704. [Google Scholar] [CrossRef]
- Barnes, S.; Kirk, M.; Coward, L. Isoflavones and Their Conjugates in Soy Foods: Extraction Conditions and Analysis by HPLC-Mass Spectrometry. J. Agric. Food Chem. 1994, 42, 2466–2474. [Google Scholar] [CrossRef]
- Totta, P.; Acconcia, F.; Virgili, F.; Cassidy, A.; Weinberg, P.D.; Rimbach, G.; Marino, M. Daidzein-sulfate metabolites affect transcriptional and antiproliferative activities of estrogen receptor-β in cultured human cancer cells. J. Nutr. 2005, 135, 2687–2693. [Google Scholar] [CrossRef] [Green Version]
- Rowland, I.R.; Wiseman, H.; Sanders, T.A.B.; Adlercreutz, H.; Bowey, E.A. Interindividual variation in metabolism of soy isoflavones and lignans: Influence of habitual diet on equol production by the gut microflora. Nutr. Cancer 2000, 36, 27–32. [Google Scholar] [CrossRef]
- Soidinsalo, O.; Wähälä, K. Synthesis of phytoestrogenic isoflavonoid disulfates. Steroids 2004, 69, 613–616. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Faughnan, M.S.; Avades, T.; Zimmer-Nechemias, L.; Brown, N.M.; Wolfe, B.E.; Brashear, W.T.; Desai, P.; Oldfield, M.F.; Botting, N.P.; et al. Comparing the pharmacokinetics of daidzein and genistein with the use of 13C-labeled tracers in premenopausal women. Am. J. Clin. Nutr. 2003, 77, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanti, P.; Sawaya, B.P.; Custer, L.J.; Franke, A.A. Serum levels and metabolic clearance of the isoflavones genistein and daidzein in hemodialysis patients. J. Am. Soc. Nephrol. 1999, 10, 864–871. [Google Scholar] [PubMed]
- Qu, Y.; Zhang, Z.; Lu, Y.; Zheng, D.; Wei, Y. Network Pharmacology Reveals the Molecular Mechanism of Cuyuxunxi Prescription in Promoting Wound Healing in Patients with Anal Fistula. Evid. Based Complement. Altern. Med. 2019, 2019, 3865121. [Google Scholar] [CrossRef]
- Luo, L.Y.; Fan, M.X.; Zhao, H.Y.; Li, M.X.; Wu, X.; Gao, W.Y. Pharmacokinetics and Bioavailability of the Isoflavones Formononetin and Ononin and Their in Vitro Absorption in Ussing Chamber and Caco-2 Cell Models. J. Agric. Food Chem. 2018, 66, 2917–2924. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Wahajuddin; Raju, K.S.R.; Ali, M.M.; Kohli, K.; Jain, G.K. Reduced bioavailability of tamoxifen and its metabolite 4-hydroxytamoxifen after oral administration with Biochanin A (an Isoflavone) in rats. Phyther. Res. 2012, 26, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Wahajuddin; Tewari, D.; Pradhan, T.; Jain, G.K. PAMPA permeability, plasma protein binding, blood partition, pharmacokinetics and metabolism of formononetin, a methoxylated isoflavone. Food Chem. Toxicol. 2011, 49, 1056–1062. [Google Scholar] [CrossRef]
- Han, H.K.; Lee, B.J.; Lee, H.K. Enhanced dissolution and bioavailability of biochanin A via the preparation of solid dispersion: In vitro and in vivo evaluation. Int. J. Pharm. 2011, 415, 89–94. [Google Scholar] [CrossRef]
- Moon, Y.J.; Sagawa, K.; Frederick, K.; Zhang, S.; Morris, M.E. Pharmacokinetics and bioavailability of the isoflavone biochanin A in rats. AAPS J. 2006, 8, E433–E442. [Google Scholar]
- Setchell, K.D.R.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D.; et al. S-equol, a potent ligand for estrogen receptor β, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.C.; Vom Saal, F.S.; Welshons, W.V. The Effective Free Fraction of Estradiol Measured by Whole Cell Uptake Assays: And Xenoestrogens in Human Serum Physiology of Delivery Modifies Estrogenic Activity. Exp. Biol. Med. 1998, 217, 300–309. [Google Scholar] [CrossRef]
- Bennetts, H.W.; Underwood, E.J.; Shier, F.L. A specific breeding problem of sheep on subterranean clover pastures in Western Australia. Br. Vet. J. 1946, 102, 348–352. [Google Scholar] [PubMed]
- Millington, A.J.; Francis, C.M.; Mckeown, N.R. Wether bioassay of annual pasture legumes. II. The oestrogenic activity of nine strains of Trifolium subterraneum L. Aust. J. Agric. Res. 1964, 15, 527–536. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Penalvo, J.L.; Gil, J.I.; Medina, S.; Horcajada, M.N.; Lafay, S.; Silberberg, M.; Llorach, R.; Zafrilla, P.; Garcia-Mora, P.; et al. Soy Isoflavones and Cardiovascular Disease Epidemiological, Clinical and -Omics Perspectives. Curr. Pharm. Biotechnol. 2012, 13, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Gardana, C.; Canzi, E.; Simonetti, P. The role of diet in the metabolism of daidzein by human faecal microbiota sampled from Italian volunteers. J. Nutr. Biochem. 2009, 20, 940–947. [Google Scholar] [CrossRef]
- Nagata, C.; Ueno, T.; Uchiyama, S.; Nagao, Y.; Yamamoto, S.; Shibuya, C.; Kashiki, Y.; Shimizu, H. Dietary and lifestyle correlates of urinary excretion status of equol in Japanese women. Nutr. Cancer 2008, 60, 49–54. [Google Scholar] [CrossRef]
- Tanaka, M.; Fujimoto, K.; Chihara, Y.; Torimoto, K.; Yoneda, T.; Tanaka, N.; Hirayama, A.; Miyanaga, N.; Akaza, H.; Hirao, Y. Isoflavone supplements stimulated the production of serum equol and decreased the serum dihydrotestosterone levels in healthy male volunteers. Prostate Cancer Prostatic Dis. 2009, 12, 247–252. [Google Scholar] [CrossRef]
- Liu, Z.M.; Ho, S.C.; Chen, Y.M.; Tomlinson, B.; Ho, S.; To, K.; Woo, J. Effect of whole soy and purified daidzein on ambulatory blood pressure and endothelial function-a 6-month double-blind, randomized controlled trial among Chinese postmenopausal women with prehypertension. Eur. J. Clin. Nutr. 2015, 69, 1161–1168. [Google Scholar] [CrossRef]
- Jackman, K.A.; Woodman, O.L.; Chrissobolis, S.; Sobey, C.G. Vasorelaxant and antioxidant activity of the isoflavone metabolite equol in carotid and cerebral arteries. Brain Res. 2007, 1141, 99–107. [Google Scholar] [CrossRef]
- Kelly, G.E.; Joannou, G.E.; Reeder, A.Y.; Nelson, C.; Waring, M.A. The Variable Metabolic Response to Dietary Isoflavones in Humans. Proc. Soc. Exp. Biol. Med. 1995, 208, 40–43. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Patisaul, H.B.; Williams, C.J. Reproductive Consequences of Developmental Phytoestrogen Exposure. Reproduction 2012, 143, 247–260. [Google Scholar] [CrossRef]
- Piotrowska, K.; Baranowska-Bosiacka, I.; Marchlewicz, M.; Gutowska, I.; Noceń, I.; Zawiślak, M.; Chlubek, D.; Wiszniewska, B. Changes in male reproductive system and mineral metabolism induced by soy isoflavones administered to rats from prenatal life until sexual maturity. Nutrition 2011, 27, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Talsness, C.; Grote, K.; Kuriyama, S.; Presibella, K.; Sterner-Kock, A.; Poça, K.; Chahoud, I. Prenatal Exposure to the Phytoestrogen Daidzein Resulted in Persistent Changes in Ovarian Surface Epithelial Cell Height, Folliculogenesis, and Estrus Phase Length in Adult Sprague-Dawley Rat Offspring. J. Toxicol. Environ. Health Part A Curr. Issues 2015, 78, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; Bredfeldt, T.; Everitt, J.I.; Lin, K.; Berry, T.; Kannan, K.; Mittelstadt, M.L.; Ho, S.M.; Walker, C.L. Environmental estrogens differentially engage the histone methyltransferase EZH2 to increase risk of uterine tumorigenesis. Mol. Cancer Res. 2012, 10, 546–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Doorn-Khosrovani, S.B.v.W.; Janssen, J.; Maas, L.M.; Godschalk, R.W.L.; Nijhuis, J.G.; van Schooten, F.J. Dietary flavonoids induce MLL translocations in primary human CD34+ cells. Carcinogenesis 2007, 28, 1703–1709. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, B.; Thorstensen, E.B.; Ponnampalam, A.P.; Mitchell, M.D. Transplacental Transfer and Biotransformation of Genistein in Human Placenta. Placenta 2010, 31, 506–511. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Zimmer-Nechemias, L.; Cai, J.; Heubi, J.E. Exposure of infants to phyto-oestrogens from soy-based infant formula. Lancet 1997, 350, 23–27. [Google Scholar] [CrossRef]
- Franke, A.A.; Custer, L.J. Daidzein and genistein concentrations in human milk after soy consumption. Clin. Chem. 1996, 42, 955–964. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.A.; Lai, J.F.; Halm, B.M. Absorption, distribution, metabolism, and excretion of isoflavonoids after soy intake. Arch. Biochem. Biophys. 2014, 559, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.; Lynn, H.; Lau, W.Y.; Leung, J.; Lau, E.; Wong, S.Y.S.; Kwok, T. Nutrient intake and psychological health in an elderly Chinese population. Int. J. Geriatr. Psychiatry 2006, 21, 1036–1043. [Google Scholar] [CrossRef]
- Hogervorst, E.; Sadjimim, T.; Yesufu, A.; Kreager, P.; Rahardjo, T.B. High tofu intake is associated with worse memory in elderly Indonesian men and women. Dement. Geriatr. Cogn. Disord. 2008, 26, 50–57. [Google Scholar] [CrossRef]
- White, L.R.; White, L.R.; Petrovitch, H.; White, L.R.; Petrovitch, H.; Ross, G.W.; Masaki, K.; Hardman, J.; White, L.R.; Ross, G.W.; et al. Brain Aging and Midlife Tofu Consumption. J. Am. Coll. Nutr. 2000, 19, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Coates, P.; Betz, J.M.; Blackman, M.R.; Cragg, G.; Levine, M.; Moss, J.; White, J. Encyclopedia of Dietary Supplements Encyclopedia of Dietary Supplements; Taylor & Francis Group LLC: Boca Raton, FL, USA; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781439819289. [Google Scholar]
- Chang, H.C.; Doerge, D.R. Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol. Appl. Pharmacol. 2000, 168, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Radović, B.; Mentrup, B.; Köhrle, J. Genistein and other soya isoflavones are potent ligands for transthyretin in serum and cerebrospinal fluid. Br. J. Nutr. 2006, 95, 1171–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chorazy, P.A.; Himelhoch, S.; Hopwood, N.J.; Greger, N.G.; Postellon, D.C. Persistent hypothyroidism in an infant receiving a soy formula: Case report and review of the literature. Pediatrics 1995, 96, 148–149. [Google Scholar] [PubMed]
- Dillingham, B.L.; McVeigh, B.L.; Lampe, J.W.; Duncan, A.M. Soy Protein Isolates of Varied Isoflavone Content Do Not Influence Serum Thyroid Hormones in Healthy Young Men. Thyroid 2007, 17, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Redmond, G. Effects of soy protein and soybean isoflavones on thyroid function in healthy adults and hypothyroid patients: A review of the relevant literature. Thyroid 2006, 16, 249–258. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Bao, B.; Ahmad, A.; Sarkar, F.H. Induction of cancer cell death by isoflavone: The role of multiple signaling pathways. Nutrients 2011, 3, 877–896. [Google Scholar] [CrossRef] [Green Version]
- Van Die, M.D.; Bone, K.M.; Williams, S.G.; Pirotta, M.V. Soy and soy isoflavones in prostate cancer: A systematic review and meta-analysis of randomized controlled trials. BJU Int. 2014, 113, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Jing, X.; Li, H.; Zhao, X.; Wang, D. Soy isoflavone consumption and colorectal cancer risk: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 25939. [Google Scholar] [CrossRef]
- Sun, C.L.; Yuan, J.M.; Wang, X.L.; Gao, Y.T.; Ross, R.K.; Yu, M.C. Dietary soy and increased risk of bladder cancer: A prospective cohort study of men in Shanghai, China. Int. J. Cancer 2004, 112, 319–323. [Google Scholar] [CrossRef]
- Sun, C.L.; Yuan, J.M.; Arakawa, K.; Low, S.H.; Lee, H.P.; Yu, M.C. Dietary soy and increased risk of bladder cancer: The Singapore Chinese Health Study. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1674–1677. [Google Scholar]
- Rossi, M.; Strikoudi, P.; Spei, M.E.; Parpinel, M.; Serraino, D.; Montella, M.; Libra, M.; La Vecchia, C.; Rosato, V. Flavonoids and bladder cancer risk. Cancer Causes Control 2019, 30, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Messing, E.; Gee, J.R.; Saltzstein, D.R.; Kim, K.M.; DiSant’Agnese, A.; Kolesar, J.; Harris, L.; Faerber, A.; Havighurst, T.; Young, J.M.; et al. A phase 2 cancer chemoprevention biomarker trial of isoflavone G-2535 (genistein) in presurgical bladder cancer patients. Cancer Prev. Res. 2012, 5, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, M.J.; Wood, C.E. Soy isoflavones, estrogen therapy, and breast cancer risk: Analysis and commentary. Nutr. J. 2008, 7, 1–12. [Google Scholar]
- Shu, X.O.; Zheng, Y.; Cai, H.; Gu, K.; Chen, Z.; Zheng, W.; Lu, W. Soy food intake and breast cancer survival. JAMA J. Am. Med. Assoc. 2009, 302, 2437–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.R.; Joubert, C.; Boniol, M.; Hery, C.; Ahn, S.H.; Won, Y.J.; Nishino, Y.; Sobue, T.; Chen, C.J.; You, S.L.; et al. Recent trends and patterns in breast cancer incidence among Eastern and Southeastern Asian women. Cancer Causes Control 2010, 21, 1777–1785. [Google Scholar] [CrossRef]
- van Erp-Baart, M.-A.J.; Brants, H.A.M.; Kiely, M.; Mulligan, A.; Turrini, A.; Sermoneta, C.; Kilkkinen, A.; Valsta, L.M. Isoflavone intake in four different European countries: The VENUS approach. Br. J. Nutr. 2003, 89, S25–S30. [Google Scholar] [CrossRef] [Green Version]
- Messina, M.; Nagata, C.; Wu, A.H. Estimated Asian adult soy protein and isoflavone intakes. Nutr. Cancer 2006, 55, 1–12. [Google Scholar] [CrossRef]
- Messina, M.; Hilakivi-Clarke, L. Early intake appears to be the key to the proposed protective effects of soy intake against breast cancer. Nutr. Cancer 2009, 61, 792–798. [Google Scholar] [CrossRef]
- Nishio, K.; Niwa, Y.; Toyoshima, H.; Tamakoshi, K.; Kondo, T.; Yatsuya, H.; Yamamoto, A.; Suzuki, S.; Tokudome, S.; Lin, Y.; et al. Consumption of soy foods and the risk of breast cancer: Findings from the Japan Collaborative Cohort (JACC) Study. Cancer Causes Control 2007, 18, 801–808. [Google Scholar] [CrossRef]
- McMichael-Phillips, D.F.; Harding, C.; Morton, M.; Roberts, S.A.; Howell, A.; Potten, C.S.; Bundred, N.J. Effects of soy-protein supplementation on epithelial proliferation in the histologically normal human breast. Am. J. Clin. Nutr. 1998, 68, 1431S–1435S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn-Ross, P.L.; John, E.M.; Canchola, A.J.; Stewart, S.L.; Lee, M.M. Phytoestrogen intake and endometrial cancer risk. J. Natl. Cancer Inst. 2003, 95, 1158–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.H.; Zheng, W.; Xiang, Y.B.; Ruan, Z.X.; Cheng, J.R.; Dai, Q.; Gao, Y.T.; Shu, X.O. Soya food intake and risk of endometrial cancer among Chinese women in Shanghai: Population based case-control study. Br. Med. J. 2004, 328, 1285–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.J.; Meyer, W.R.; Lessey, B.A.; Oi, R.H.; DeWire, R.E.; Fritz, M.A. Soy protein isolate with isoflavones does not prevent estradiol-induced endometrial hyperplasia in postmenopausal women: A pilot trial. Menopause 2003, 10, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, R.J.; Croker, K.P.; Neil, H.G. Failure of sperm transport in relation to ewe infertility following prolonged grazing on oestrogenic pastures. Aust. J. Agric. Res. 1967, 18, 740–753. [Google Scholar] [CrossRef]
- Mustonen, E.; Taponen, S.; Andersson, M.; Sukura, A.; Katila, T.; Taponen, J. Fertility and growth of nulliparous ewes after feeding red clover silage with high phyto-oestrogen concentrations. Animal 2014, 8, 1699–1705. [Google Scholar] [CrossRef] [Green Version]
- Sandoo, A.; Veldhuijzen van Zanten, J.J.C.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. Open Cardiovasc. Med. J. 2015, 4, 302–312. [Google Scholar] [CrossRef]
- Aalkjær, C.; Boedtkjer, D.; Matchkov, V. Vasomotion—What is currently thought? Acta Physiol. 2011, 202, 253–269. [Google Scholar] [CrossRef]
- Wong, B.J. Sensory nerves and nitric oxide contribute to reflex cutaneous vasodilation in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, 651–656. [Google Scholar] [CrossRef]
- Ghosh, D.; Syed, A.U.; Prada, M.P.; Nystoriak, M.A.; Santana, L.F.; Nieves-Cintrón, M.; Navedo, M.F. Calcium Channels in Vascular Smooth Muscle. Adv. Pharmacol. 2017, 78, 49–87. [Google Scholar]
- Fernández-Tenorio, M.; Porras-González, C.; Castellano, A.; Del Valle-Rodríguez, A.; López-Barneo, J.; Ureña, J. Metabotropic Regulation of RhoA/Rho-Associated Kinase by New Mechanism for Depolarization-Evoked Mammalian Arterial Contraction SR. Circ. Res. 2011, 108, 1348–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chambliss, K.L.; Shaul, P.W. Estrogen modulation of endothelial nitric oxide synthase. Endocr. Rev. 2002, 23, 665–686. [Google Scholar] [CrossRef] [Green Version]
- Campbell, W.B.; Gauthier, K.M. Role of the 15-Lipoxygenase-EDHF Pathway. J. Cardiovasc. Pharmacol. 2014, 61, 176–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, S.W.; Yeum, C.H.; Webb, C.; Campbell, G. Serotonin Stimulates Protein Tyrosyl Phosphorylation and Vascular Contraction via Tyrosine Kinase. J. Vasc. Res. 1996, 33, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Nevala, R.; Korpela, R.; Vapaatalo, H. Plant Derived Estrogens Relax Rat Mesenteric Artery In Vitro. Life Sci. 1998, 63, PL95–PL100. [Google Scholar] [CrossRef]
- Mishra, S.K.; Abbot, S.E.; Choudhury, Z.; Cheng, M.; Khatab, N.; Maycock, N.J.R.; Zavery, A.; Aaronson, P.I. Endothelium-dependent relaxation of rat aorta and main pulmonary artery by the phytoestrogens genistein and daidzein. Cardiovasc. Res. 2000, 46, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Ajay, M.; Gilani, A.U.H.; Mustafa, M.R. Effects of flavonoids on vascular smooth muscle of the isolated rat thoracic aorta. Life Sci. 2003, 74, 603–612. [Google Scholar] [CrossRef]
- Galán-Martínez, L.; Herrera-Estrada, I.; Fleites-Vázquez, A. Direct actions of the flavonoids naringenin, quercetin and genistein on rat cardiac and vascular muscles. J. Pharm. Pharmacogn. Res. 2018, 6, 158–166. [Google Scholar]
- Je, H.D.; Sohn, U.D. Inhibitory effect of genistein on agonist-induced modulation of vascular contractility. Mol. Cells 2009, 27, 191–198. [Google Scholar] [CrossRef]
- Filipeanu, C.M.; Brailoiu, E.; Huhurez, G.; Slatineanu, S.; Baltatu, O.; Branisteanu, D.D. Multiple effects of tyrosine kinase inhibitors on vascular smooth muscle contraction. Eur. J. Pharmacol. 1995, 281, 29–35. [Google Scholar] [CrossRef]
- Sun, T.; Liu, R.; Cao, Y.X. Vasorelaxant and antihypertensive effects of formononetin through endothelium-dependent and -independent mechanisms. Acta Pharmacol. Sin. 2011, 32, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Torregrosa, G.; Burguete, M.C.; Pérez-Asensio, F.J.; Salom, J.B.; Gil, J.V.; Alborch, E. Pharmacological profile of phytoestrogens in cerebral vessels: In vitro study with rabbit basilar artery. Eur. J. Pharmacol. 2003, 482, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Altavilla, D.; Squadrito, G.; Saitta, A.; Cucinotta, D.; Minutoli, L.; Deodato, B.; Ferlito, M.; Campo, G.M.; Bova, A.; et al. Genistein supplementation and estrogen replacement therapy improve endothelial dysfunction induced by ovariectomy in rats. Cardiovasc. Res. 2000, 45, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Pinna, C.; Sala, A. Sex-specific activity of hesperidin, diosmin and genistein on human umbilical vein. Biomed. Res. Clin. Pract. 2019, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Roghani, M.; Vaez Mahdavi, M.R.; Jalali-Nadoushan, M.R.; Baluchnejadmojarad, T.; Naderi, G.; Roghani-Dehkordi, F.; Taghi Joghataei, M.; Kord, M. Chronic administration of daidzein, a soybean isoflavone, improves endothelial dysfunction and attenuates oxidative stress in streptozotocin- induced diabetic rats. Phyther. Res. 2013, 27, 112–117. [Google Scholar] [CrossRef]
- Prawez, S.; Ahanger, A.A.; Singh, T.U.; Mishra, S.K.; Sarkar, S.N.; Rahman, S. Chronic administration of phytoestrogen “daidzein” to ameliorate mean arterial pressure and vascular function in N-G-nitro-L-arginine methyl ester hypertensive rats. Vet. Arh. 2015, 85, 451–464. [Google Scholar]
- Woodman, O.L.; Missen, M.A.; Boujaoude, M. Daidzein and 17β-estradiol enhance nitric oxide synthase activity associated with an increase in calmodulin and a decrease in caveolin-1. J. Cardiovasc. Pharmacol. 2004, 44, 155–163. [Google Scholar] [CrossRef]
- Sobey, C.G.; Weiler, J.M.; Boujaoude, M.; Woodman, O.L. Effect of short-term phytoestrogen treatment in male rats on nitric oxide-mediated responses of carotid and cerebral arteries: Comparison with 17β-estradiol. J. Pharmacol. Exp. Ther. 2004, 310, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Singh, M.; Sharma, P.L. Ameliorative effect of daidzein: A caveolin-1 inhibitor in vascular endothelium dysfunction induced by ovariectomy. Indian J. Exp. Biol. 2012, 50, 28–34. [Google Scholar]
- Chen, Z.; Oliveira, S.D.S.; Zimnicka, A.M.; Jiang, Y.; Sharma, T.; Chen, S.; Lazarov, O.; Bonini, M.G.; Haus, J.M.; Minshall, R.D. Reciprocal regulation of eNOS and caveolin-1 functions in endothelial cells. Mol. Biol. Cell 2018, 29, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- GOVERS, R.; RABELINK, T.J. Cellular regulation of endothelial nitric oxide synthase. Am. J. Physiol. Ren. Physiol. 2001, 280, F193–F206. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, K.; Nakaki, T.; Marumo, T.; Suzuki, H.; Kato, R.; Saruta, T. Up-regulation of nitric oxide synthase by estradiol in human aorticendothelial cells. FEBS Lett. 1995, 360, 291–293. [Google Scholar]
- Michel, J.B.; Feron, O.; Sacks, D.; Michel, T. Reciprocal regulation of endothelial nitric-oxide synthase by Ca2+-calmodulin and caveolin. J. Biol. Chem. 1997, 272, 15583–15586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.H.; Li, Q.; Wu, M.Y.; Guo, D.J.; Chen, H.L.; Chen, S.L.; Seto, S.W.; Au, A.L.S.; Poon, C.C.W.; Leung, G.P.H.; et al. Formononetin, an isoflavone, relaxes rat isolated aorta through endothelium-dependent and endothelium-independent pathways. J. Nutr. Biochem. 2010, 21, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.H.L.; Vong, C.T.; Leung, G.P.H.; Seto, S.W.; Kwan, Y.W.; Lee, S.M.Y.; Hoi, M.P.M. Calycosin and Formononetin Induce Endothelium-Dependent Vasodilation by the Activation of Large-Conductance Ca2+-Activated K+ Channels (BKCa). Evid. Based Complement. Altern. Med. 2016, 2016, 5272531. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zhong, Y.; Tang, T.; Luo, J.; Cui, H.; Fan, R.; Wang, Y.; Wang, D. Formononetin induces vasorelaxation in rat thoracic aorta via regulation of the PI3K/PTEN/Akt signaling pathway. Drug Des. Devel. Ther. 2018, 12, 3675–3684. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Cao, L.; Ping, N.N.; Wu, Y.; Liu, D.Z.; Cao, Y.X. Formononetin upregulates nitric oxide synthase in arterial endothelium through estrogen receptors and MAPK pathways. J. Pharm. Pharmacol. 2016, 68, 342–351. [Google Scholar] [CrossRef]
- Zheng, J.; Bird, I.M.; Melsaether, A.N.; Magness, R.R. Activation of the mitogen-activated protein kinase cascade is necessary but not sufficient for basic fibroblast growth factor- and epidermal growth factor-stimulated expression of endothelial nitric oxide synthase in ovine fetoplacental artery endothelial. Endocrinology 1999, 140, 1399–1407. [Google Scholar] [CrossRef]
- Mata-Greenwood, E.; Liao, W.-X.; Zheng, J.; Chen, D.-B. Differential Activation of Multiple Signaling Pathways Dictates eNOS Upregulation by FGF2 but not VEGF in Placental Artery Endothelial Cells. Placenta 2008, 29, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Jiang, Y.; Liu, J.; Zhao, K.; Mo, Y.; Liu, Z.; Zeng, Y. Downregulation of human endothelial nitric oxide synthase promoter activity by p38 mitogen-activated protein kinase activation. Biochem. Cell Biol. 2006, 84, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Siow, R.C.M.; Li, F.Y.L.; Rowlands, D.J.; de Winter, P.; Mann, G.E. Cardiovascular targets for estrogens and phytoestrogens: Transcriptional regulation of nitric oxide synthase and antioxidant defense genes. Free Radic. Biol. Med. 2007, 42, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.P.; Gao, Q.; Mei, R.H.; Zhao, M.H.; Lu, Y.; Li, X.Y.; Bruce, I.C.; Xia, Q. Mechanisms underlying biochanin A-induced relaxation of the aorta differ between normotensive and hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2006, 33, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Jung, W.S.; Cho, N.S.; Ryu, K.H.; Jun, J.Y.; Shin, B.C.; Chung, J.H.; Yeum, C.H. Mechanisms of phytoestrogen biochanin A-induced vasorelaxation in renovascular hypertensive rats. Kidney Res. Clin. Pract. 2014, 33, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Migkos, T.; Pourová, J.; Vopršalová, M.; Auger, C.; Schini-Kerth, V.; Mladěnka, P. Biochanin A, the Most Potent of 16 Isoflavones, Induces Relaxation of the Coronary Artery through the Calcium Channel and cGMP-dependent Pathway. Planta Med. 2020, 86, 708–716. [Google Scholar] [CrossRef]
- Chin-Dusting, J.P.F.; Fisher, L.J.; Lewis, T.V.; Piekarska, A.; Nestel, P.J.; Husband, A. The vascular activity of some isoflavone metabolites: Implications for a cardioprotective role. Br. J. Pharmacol. 2001, 133, 595–605. [Google Scholar] [CrossRef]
- Joy, S.; Siow, R.C.M.; Rowlands, D.J.; Becker, M.; Wyatt, A.W.; Aaronson, P.I.; Coen, C.W.; Kallo, I.; Jacob, R.; Mann, G.E. The isoflavone equol mediates rapid vascular relaxation: Ca2+-independent activation of endothelial nitric-oxide synthase/Hsp90 involving ERK1/2 and Akt phosphorylation in human endothelial cells. J. Biol. Chem. 2006, 281, 27335–27345. [Google Scholar] [CrossRef] [Green Version]
- Ohkura, Y.; Obayashi, S.; Yamada, K.; Yamada, M.; Kubota, T. S-equol partially restored endothelial nitric oxide production in isoflavone-deficient ovariectomized rats. J. Cardiovasc. Pharmacol. 2015, 65, 500–507. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, M.Y.; Park, H.M. The Effect of Eqoul, a Metabolite of Isoflavone, on Endothelial Cell-independent Vasodilatation of Human Uterine Artery In Vitro. J. Bone Metab. 2015, 22, 57. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Wang, X.; Weakley, S.M.; Kougias, P.; Lin, P.H.; Yao, Q.; Chen, C. The soybean isoflavonoid equol blocks ritonavir-induced endothelial dysfunction in porcine pulmonary arteries and human pulmonary artery endothelial cells. J. Nutr. 2010, 140, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.W.; Keung, W.; Xu, Y.C.; Ng, K.F.; Leung, G.P.; Vanhoutte, P.M.; Choy, P.C.; Man, R.Y. Genistein potentiates protein kinase A activity in porcine coronary artery. Mol. Cell. Biochem. 2008, 311, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, B.N.; Wang, S.B.; Wang, S.H.; Du, G.H. Vasorelaxant effect of formononetin in the rat thoracic aorta and its mechanisms. J. Asian Nat. Prod. Res. 2012, 14, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.P.; Mei, R.H.; Li, X.Y.; Zhao, M.H.; Lu, Y.; Bruce, I.C.; Xia, Q. Endothelium-independent vasorelaxant effect of the phyto-oestrogen biochanin A on rat thoracic aorta. In Proceedings of the 2005 IEEE Engineering in Medicine and Biology 27th Annual Conference, Shanghai, China, 17–18 January 2006; Volume 3, pp. 2244–2247. [Google Scholar]
- Gimenez, I.; Lou, M.; Vargas, F.; Alvarez-Guerra, M.; Mayoral, J.A.; Martinez, R.M.; Garay, R.P.; Alda, J.O. Renal and vascular actions of equol in the rat. J. Hypertens. 1997, 15, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, S.V.; Aaronson, P.I. Inhibition of vascular smooth muscle cell K+ currents by tyrosine kinase inhibitors genistein and ST 638. Circ. Res. 1995, 76, 310–316. [Google Scholar] [CrossRef]
- Woodman, O.L.; Boujaoude, M. Chronic treatment of male rats with daidzein and 17β-oestradiol induces the contribution of EDHF to endothelium-dependent relaxation. Br. J. Pharmacol. 2004, 141, 322–328. [Google Scholar] [CrossRef]
- Zhang, H.T.; Wang, Y.; Deng, X.L.; Dong, M.Q.; Zhao, L.M.; Wang, Y.W. Daidzein relaxes rat cerebral basilar artery via activation of large-conductance Ca2+-activated K+ channels in vascular smooth muscle cells. Eur. J. Pharmacol. 2010, 630, 100–106. [Google Scholar] [CrossRef]
- Nevala, R.; Paukku, K.; Korpela, R.; Vapaatalo, H. Calcium-sensitive potassium channel inhibitors antagonize genistein- and daidzein-induced arterial relaxation in vitro. Life Sci. 2001, 69, 1407–1417. [Google Scholar] [CrossRef]
- Chan, H.Y.; Yao, X.; Tsang, S.Y.; Chan, F.L.; Lau, C.W.; Huang, Y. Different role of endothelium/nitric oxide in 17β-estradiol- and progesterone-induced relaxation in rat arteries. Life Sci. 2001, 69, 1609–1617. [Google Scholar] [CrossRef]
- Kang, K.T. Endothelium-derived relaxing factors of small resistance arteries in hypertension. Toxicol. Res. 2014, 30, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Garland, C.J.; Dora, K.A. EDH: Endothelium-dependent hyperpolarization and microvascular signalling. Acta Physiol. 2017, 219, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Takayanagi, K.; Kobayashi, S.; Kojima, M.; Taguchi, K.; Kobayashi, T. Effect of equol on vasocontractions in rat carotid arteries treated with high insulin. Biol. Pharm. Bull. 2019, 42, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Wang, Y.; Song, Z.; Zhao, L.M.; Li, G.R.; Deng, X.L. Equol increases cerebral blood flow in rats via activation of large-conductance Ca2+-activated K+ channels in vascular smooth muscle cells. Pharmacol. Res. 2016, 107, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Cowled, P.A.; Fitridge, R.A. Nitric oxide and skeletal muscle reperfusion injury: Current controversies (research review). J. Surg. Res. 2005, 128, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Valero, M.S.; Garay, R.P.; Gros, P.; Alda, J.O. Cystic fibrosis transmembrane conductance regulator (CFTR) chloride channel and Na-K-Cl cotransporter NKCC1 isoform mediate the vasorelaxant action of genistein in isolated rat aorta. Eur. J. Pharmacol. 2006, 544, 126–131. [Google Scholar] [CrossRef]
- Satake, N.; Shibata, S. The potentiating effect of genistein on the relaxation induced by isoproterenol in rat aortic rings. Gen. Pharmacol. 1999, 33, 221–227. [Google Scholar] [CrossRef]
- Abebe, W.; Agrawal, D.K. Role of tyrosine kinases in norepinephrine-induced contraction of vascular smooth muscle. J. Cardiovasc. Pharmacol. 1995, 26, 153–159. [Google Scholar] [CrossRef]
- Inoue, R.; Waniishi, Y.; Yamada, K.; Ito, Y. A possible role of tyrosine kinases in the regulation of muscarinic receptor-activated cation channels in guinea pig ileum. Biochem. Biophys. Res. Commun. 1994, 203, 1392–1397. [Google Scholar] [CrossRef]
- Liu, H.; Li, K.; Sperelakis, N. Tyrosine kinase inhibitor, genistein, inhibits macroscopic L-type calcium current in rat portal vein smooth muscle cells. Can. J. Physiol. Pharmacol. 1997, 75, 1058–1062. [Google Scholar] [CrossRef]
- Kitazono, T.; Ibayashi, S.; Nagao, T.; Kagiyama, T.; Kitayama, J.; Fujishima, M. Role of tyrosine kinase in serotonin-induced constriction of the basilar artery in vivo. Stroke 1998, 29, 494–498. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Ishida, J.; Nakagawa, S.; Ogawara, H.; Watanabe, S.; Itoh, N.; Shibuya, M.; Fukami, Y. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 1987, 262, 5592–5595. [Google Scholar] [CrossRef]
- Zerrouk, A.; Auguet, M.; Dabiré, H.; Brisac, A.M.; Safar, M.; Chabrier, P.E. Differential effects of tyrosine kinase inhibitors on contraction and relaxation of the aortas of normotensive and hypertensive rats. Eur. J. Pharmacol. 1999, 374, 49–58. [Google Scholar] [CrossRef]
- Nevala, R.; Lassila, M.; Finckenberg, P.; Paukku, K.; Korpela, R.; Vapaatalo, H. Genistein treatment reduces arterial contractions by inhibiting tyrosine kinases in ovariectomized hypertensive rats. Eur. J. Pharmacol. 2002, 452, 87–96. [Google Scholar] [CrossRef]
- Lee, M.Y.K.; Man, R.Y.K. The phytoestrogen genistein enhances endothelium-independent relaxation in the porcine coronary artery. Eur. J. Pharmacol. 2003, 481, 227–232. [Google Scholar] [CrossRef]
- Kimoto, Y.; Kinoshita, H.; Nakahata, K.; Dojo, M.; Hatano, Y. Inhibitory effects of lidocaine and mexiletine on vasorelaxation mediated by adenosine triphosphate-sensitive K+ channels and the role of kinases in the porcine coronary artery. Anesthesiology 2005, 102, 581–587. [Google Scholar] [CrossRef]
- Zhou, R.; Liu, L.; Hu, D. Involvement of BKCa α subunit tyrosine phosphorylation in vascular hyporesponsiveness of superior mesenteric artery following hemorrhagic shock in rats. Cardiovasc. Res. 2005, 68, 327–335. [Google Scholar] [CrossRef]
- Duarte, J.; Ocete, M.A.; Pérez-Vizcaino, F.; Zarzuelo, A.; Tamargo, J. Effect of tyrosine kinase and tyrosine phosphatase inhibitors on aortic contraction and induction of nitric oxide synthase. Eur. J. Pharmacol. 1997, 338, 25–33. [Google Scholar] [CrossRef]
- Bermejo, A.; Zarzuelo, A.; Duarte, J. In vivo vascular effects of genistein on a rat model of septic shock induced by lipopolysaccharide. J. Cardiovasc. Pharmacol. 2003, 42, 329–338. [Google Scholar] [CrossRef]
- Moritoki, H.; Hisayama, T.; Takeuchi, S.; Kondoh, W.; Takeji, Y. Possible involvement of tyrosine kinase in the LPS-promoted initiation of L-arginine-induced relaxation of rat aorta mediated by induction of NO synthase. Life Sci. 1995, 57, 2–7. [Google Scholar] [CrossRef]
- Yin, G.; Yan, C.; Berk, B.C. Angiotensin II signaling pathways mediated by tyrosine kinases. Int. J. Biochem. Cell Biol. 2003, 35, 780–783. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Yang, C.; Li, S.N. Effects of genistein on angiotensin-converting enzyme in rats. Life Sci. 2006, 79, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tazzeo, T.; Lippton, H.; Janssen, L.J. Role of tyrosine phosphorylation in U46619-induced vasoconstriction of pulmonary vasculature and its modulation by genistein, daidzein, and equol. J. Cardiovasc. Pharmacol. 2007, 50, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, Y.; Wang, M.; Wang, Y.; Tausif, R.; Yang, Y. Genistein rescues hypoxia-induced pulmonary arterial hypertension through estrogen receptor and β-adrenoceptor signaling. J. Nutr. Biochem. 2018, 58, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Matori, H.; Umar, S.; Nadadur, R.D.; Sharma, S.; Partow-Navid, R.; Afkhami, M.; Amjedi, M.; Eghbali, M. Genistein, a Soy Phytoestrogen, Reverses Severe Pulmonary Hypertension and Prevents Right Heart Failure in Rats Humann. Hypertension 2012, 60, 425–430. [Google Scholar] [CrossRef]
- Suenaga, H.; Kamata, K. Lysophosphatidylcholine potentiates vascular contractile responses in rat aorta via activation of tyrosine kinase. Br. J. Pharmacol. 2002, 135, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Kitayama, J.; Kitazono, T.; Ooboshi, H.; Ago, T.; Ohgami, T.; Fujishima, M.; Ibayashi, S. Chronic administration of a tyrosine kinase inhibitor restores functional and morphological changes of the basilar artery during chronic hypertension. J. Hypertens. 2002, 20, 2205–2211. [Google Scholar] [CrossRef]
- Kuiper, G.G.J.M.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; Van Der Saag, P.T.; Van Der Burg, B.; Gustafsson, J.Å. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Escande, A.; Pillon, A.; Servant, N.; Cravedi, J.P.; Larrea, F.; Muhn, P.; Nicolas, J.C.; Cavaillès, V.; Balaguer, P. Evaluation of ligand selectivity using reporter cell lines stably expressing estrogen receptor alpha or beta. Biochem. Pharmacol. 2006, 71, 1459–1469. [Google Scholar] [CrossRef]
- Song, T.T.; Hendrich, S.; Murphy, P.A. Estrogenic activity of glycitein, a soy isoflavone. J. Agric. Food Chem. 1999, 47, 1607–1610. [Google Scholar] [CrossRef] [Green Version]
- Michael, E.; Mendelsohn, R.H.K. The protective effects of estrogen on the cardiovascular system. N. Engl. J. Med. 1999, 340, 1801–1811. [Google Scholar]
- Razandi, M.; Pedram, A.; Park, S.T.; Levin, E.R. Proximal events in signaling by plasma membrane estrogen receptors. J. Biol. Chem. 2003, 278, 2701–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.H.Y.; Leung, G.P.H.; Leung, S.W.S.; Vanhoutte, P.M.; Man, R.Y.K. Genistein enhances relaxation of the spontaneously hypertensive rat aorta by transactivation of epidermal growth factor receptor following binding to membrane estrogen receptors-α and activation of a G protein-coupled, endothelial nitric oxide synthase-dependent pathway. Pharmacol. Res. 2011, 63, 181–189. [Google Scholar] [PubMed]
- Sun, T.; Wang, J.; Huang, L.H.; Cao, Y.X. Antihypertensive effect of formononetin through regulating the expressions of eNOS, 5-HT2A/1B receptors and α1-adrenoceptors in spontaneously rat arteries. Eur. J. Pharmacol. 2013, 699, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Banie, L.; Lin, G.; Ning, H.; Wang, G.; Lue, T.F.; Lin, C.S. Effects of Estrogen, Raloxifene and Levormeloxifene on α1A-Adrenergic Receptor Expression. J. Urol. 2008, 180, 2241–2246. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Bai, Y.H.; Wang, S.T.; Zhu, Z.M.; Zhang, Y.W. Research on antioxidant effects and estrogenic effect of formononetin from Trifolium pratense (red clover). Phytomedicine 2009, 16, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Laniyonu, A.; Saifeddine, M.; Ahmad, S.; Hollenberg, M.D. Regulation of vascular and gastric smooth muscle contractility by pervanadate. Br. J. Pharmacol. 1994, 113, 403–410. [Google Scholar] [CrossRef]
- Herrera, M.D.; Zarzuelo, A.; Jiménez, J.; Marhuenda, E.; Duarte, J. Effects of flavonoids on rat aortic smooth muscle contractility: Structure-activity relationships. Gen. Pharmacol. 1996, 27, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Vera, R.; Jiménez, R.; Lodi, F.; Sánchez, M.; Galisteo, M.; Zarzuelo, A.; Pérez-Vizcaíno, F.; Duarte, J. Genistein restores caveolin-1 and AT-1 receptor expression and vascular function in large vessels of ovariectomized hypertensive rats. Menopause 2007, 14, 933–940. [Google Scholar] [CrossRef]
- Baluchnejadmojarad, T.; Roghani, M. Chronic administration of genistein improves aortic reactivity of streptozotocin-diabetic rats: Mode of action. Vasc. Pharmacol. 2008, 49, 1–5. [Google Scholar] [CrossRef]
- Honoré, E.K.; Williams, J.K.; Anthony, M.S.; Clarkson, T.B. Soy isoflavones enhance coronary vascular reactivity in atherosclerotic female macaques. Fertil. Steril. 1997, 67, 148–154. [Google Scholar]
- Cho, T.M.; Peng, N.; Clark, J.T.; Novak, L.; Roysommuti, S.; Prasain, J.; Wyss, J.M. Genistein attenuates the hypertensive effects of dietary NaCl in hypertensive male rats. Endocrinology 2007, 148, 5396–5402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, H.A.; Dean, T.S.; Sanders, T.A.B.; Jackson, G.; Ritter, J.M.; Chowienczyk, P.J. The phytoestrogen genistein produces acute nitric oxide-dependent dilation of human forearm vasculature with similar potency to 17βestradiol. Circulation 2001, 103, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squadrito, F.; Altavilla, D.; Morabito, N.; Crisafulli, A.; D’Anna, R.; Corrado, F.; Ruggeri, P.; Campo, G.M.; Calapai, G.; Caputi, A.P.; et al. The effect of the phytoestrogen genistein on plasma nitric oxide concentrations, endothelin-1 levels and endothelium dependent vasodilation in postmenopausal women. Atherosclerosis 2002, 163, 339–347. [Google Scholar] [CrossRef]
- Squadrito, F.; Altavilla, D.; Crisafulli, A.; Saitta, A.; Cucinotta, D.; Morabito, N.; Anna, R.D.; Corrado, F.; Ruggeri, P.; Frisina, N.; et al. Effect of Genistein on Endothelial Function in Double-Blind, Controlled Study. Am. J. Med. 2003, 114, 470–476. [Google Scholar] [CrossRef]
- Cao, Y.X.; Yang, X.J.; Liu, J.; Li, K.X. Effects of daidzein sulfates on blood pressure and artery of rats. Basic Clin. Pharmacol. Toxicol. 2006, 99, 425–430. [Google Scholar] [CrossRef]
- Sachdeva, C.; Mishra, N.; Sharma, S. Development and characterization of enteric-coated microparticles of biochanin A for their beneficial pharmacological potential in estrogen deficient-hypertension. Drug Deliv. 2016, 23, 2044–2057. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.H.; Tsai, T.Y. Effects of equol on deoxycorticosterone acetate salt-induced hypertension and associated vascular dementia in rats. Food Funct. 2016, 7, 3444–3457. [Google Scholar] [CrossRef]
- Chin-Dusting, J.P.F.; Boak, L.; Husband, A.; Nestel, P.J. The isoflavone metabolite dehydroequol produces vasodilatation in human resistance arteries via a nitric oxide-dependent mechanism. Atherosclerosis 2004, 176, 45–48. [Google Scholar] [CrossRef]
- Nestel, P.; Fujii, A.; Zhang, L. An isoflavone metabolite reduces arterial stiffness and blood pressure in overweight men and postmenopausal women. Atherosclerosis 2007, 192, 184–189. [Google Scholar] [CrossRef]
- Morton, M.S.; Wilcox, G.; Wahlqvist, M.L.; Griffiths, K. Determination of lignans and isoflavonoids in human female plasma following dietary supplementation. J. Endocrinol. 1994, 142, 251–259. [Google Scholar] [CrossRef]
- Drawz, P.; Ghazi, L. Advances in understanding the renin-angiotensin-aldosterone system (RAAS) in blood pressure control and recent pivotal trials of RAAS blockade in heart failure and diabetic nephropathy. F1000Research 2017, 6, 1–10. [Google Scholar]
- Montenegro, M.F.; Pessa, L.R.; Tanus-santos, J.E. Isoflavone genistein inhibits the angiotensin-converting enzyme and alters the vascular responses to angiotensin I and bradykinin. Eur. J. Pharmacol. 2009, 607, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-W.; Ikeda, K.; Yamori, Y. Genistein Inhibits Expressions of NADPH Oxidase p22phox and Angiotensin II Type 1 Receptor in Aortic Endothelial Cells from Stroke-Prone Spontaneously Hypertensive Rats. Hypertens. Res. 2004, 27, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce, N.; Eyster, K.M.; Williams, J.L.; Wixon, R.; Wang, C.; Martin, D.S. Effect of Genistein on Cardiovascular Responses to Angiotensin II in Conscious Unrestrained Rats. J. Cardiovasc. Pharmacol. 2000, 36, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Pai, K.V. Angiotensin Converting Enzyme Inhibition Activity of Daidzein. Sci. Fed. Dent. Oral Res. J. 2017, 1, 92–98. [Google Scholar] [CrossRef]
- Ma, H.J.; Liu, Y.X.; Wang, F.W.; Wang, L.X.; He, R.R.; Wu, Y.M. Genistein inhibits carotid sinus baroreceptor activity in anesthetized male rats. Acta Pharmacol. Sin. 2005, 26, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Man, H.Y.; Erclik, T.; Becker, L.E.; Wang, Y.T. Modulation of baroreflex sensitivity by the state of protein tyrosine phosphorylation in the brainstem of the rat. Brain Res. 1998, 792, 141–148. [Google Scholar] [CrossRef]
- Toyohira, Y.; Ueno, S.; Tsutsui, M.; Itoh, H.; Sakai, N.; Saito, N.; Takahashi, K.; Yanagihara, N. Stimulatory effects of the soy phytoestrogen genistein on noradrenaline transporter and serotonin transporter activity. Mol. Nutr. Food Res. 2010, 54, 516–524. [Google Scholar] [CrossRef]
- Yanagihara, N.; Toyohira, Y.; Liu, M.; Ueno, S.; Tsutsui, M.; Zhang, H. Insights into the Pharmacological Effects of Soy Isoflavones on Catecholamine System. Soybean Health 2011. [Google Scholar] [CrossRef] [Green Version]
- Masilamani, M.; Wei, J.; Sampson, H.A. Regulation of the immune response by soybean isoflavones. Immunol. Res. 2012, 54, 95–110. [Google Scholar] [CrossRef]
- Palanisamy, N.; Venkataraman, A.C. Beneficial effect of genistein on lowering blood pressure and kidney toxicity in fructose-fed hypertensive rats. Br. J. Nutr. 2013, 109, 1806–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Authors | Study Type (Number of Studies Considered) | Isoflavone-Containing Product | Main Results |
|---|---|---|---|
| Hooper et al. (2008) [46] | Systematic review and meta-analysis of randomized clinical trials (n = 83) | Soybeans, soy protein isolate, and isoflavone extracts | Significant decrease in DBP |
| Arenas et al. (2008) [47] | Systematic review and meta-analysis of observation studies and clinical trials (n = 14) | Soy products | No significant variations in SBP or DBP between subjects treated with isoflavones and non-treated subjects |
| Taku et al. (2010) [48] | Systematic review and meta-analysis of randomized clinical trials (n = 14) | Isoflavones extract | Significant decrease in SBP in normotensive and in pre-hypertensive patients, with greater effects in interventions longer than 3 months |
| Dong et al. (2011) [49] | Systematic review and meta-analysis of randomized clinical trials (n = 27) | Soy protein containing isoflavones | Significant decrease in SBP and DBP in normotensive and in hypertensive subjects, more markedly in the latter. Blood pressure reductions were related to the pre-treatment BP levels of the subjects and the type of control diet used as comparison |
| Liu et al. (2011) [50] | Systematic review and meta-analysis of randomized clinical trials (n = 11) | Soy protein containing isoflavones | Significant decrease in SBP and DBP in hypertensive subjects (n = 5 trials) but not in normotensive subjects (n = 6 trials) |
| Yan et al. (2017) [51] | Systematic review and meta-analysis of observational studies (n = 17) | Soy products | Significant negative correlation between soy intake and the risk of cardiovascular disease, stroke, and coronary heart disease |
| Namazi et al. (2018) [52] | Systematic review and meta-analysis of cohort studies (n = 7) | Soy products | No significant association between high consumption of soy products and lower risk of mortality from cardiovascular disease |
| Nachvak et al. (2019) [53] | Systematic review and meta-analysis of prospective studies (n = 24) | Soybeans and soy products | Inverse relationship between consumption of soy products and cardiovascular mortality |
| Li et al. (2020) [54] | Umbrella review of epidemiological and clinical studies (n = 114) | Soybeans and soy products | Generally, isoflavone consumption is more beneficial than detrimental. A beneficial role in cardiovascular disease was observed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, H. The Vascular Effects of Isolated Isoflavones—A Focus on the Determinants of Blood Pressure Regulation. Biology 2021, 10, 49. https://doi.org/10.3390/biology10010049
Silva H. The Vascular Effects of Isolated Isoflavones—A Focus on the Determinants of Blood Pressure Regulation. Biology. 2021; 10(1):49. https://doi.org/10.3390/biology10010049
Chicago/Turabian StyleSilva, Henrique. 2021. "The Vascular Effects of Isolated Isoflavones—A Focus on the Determinants of Blood Pressure Regulation" Biology 10, no. 1: 49. https://doi.org/10.3390/biology10010049
APA StyleSilva, H. (2021). The Vascular Effects of Isolated Isoflavones—A Focus on the Determinants of Blood Pressure Regulation. Biology, 10(1), 49. https://doi.org/10.3390/biology10010049