The Metabolic Building Blocks of a Minimal Cell

 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Inference of Minimal Metabolic Networks
2.2. Reconstruction of the Directed Acyclic Graph of Metabolic Networks
3. Results and Discussion
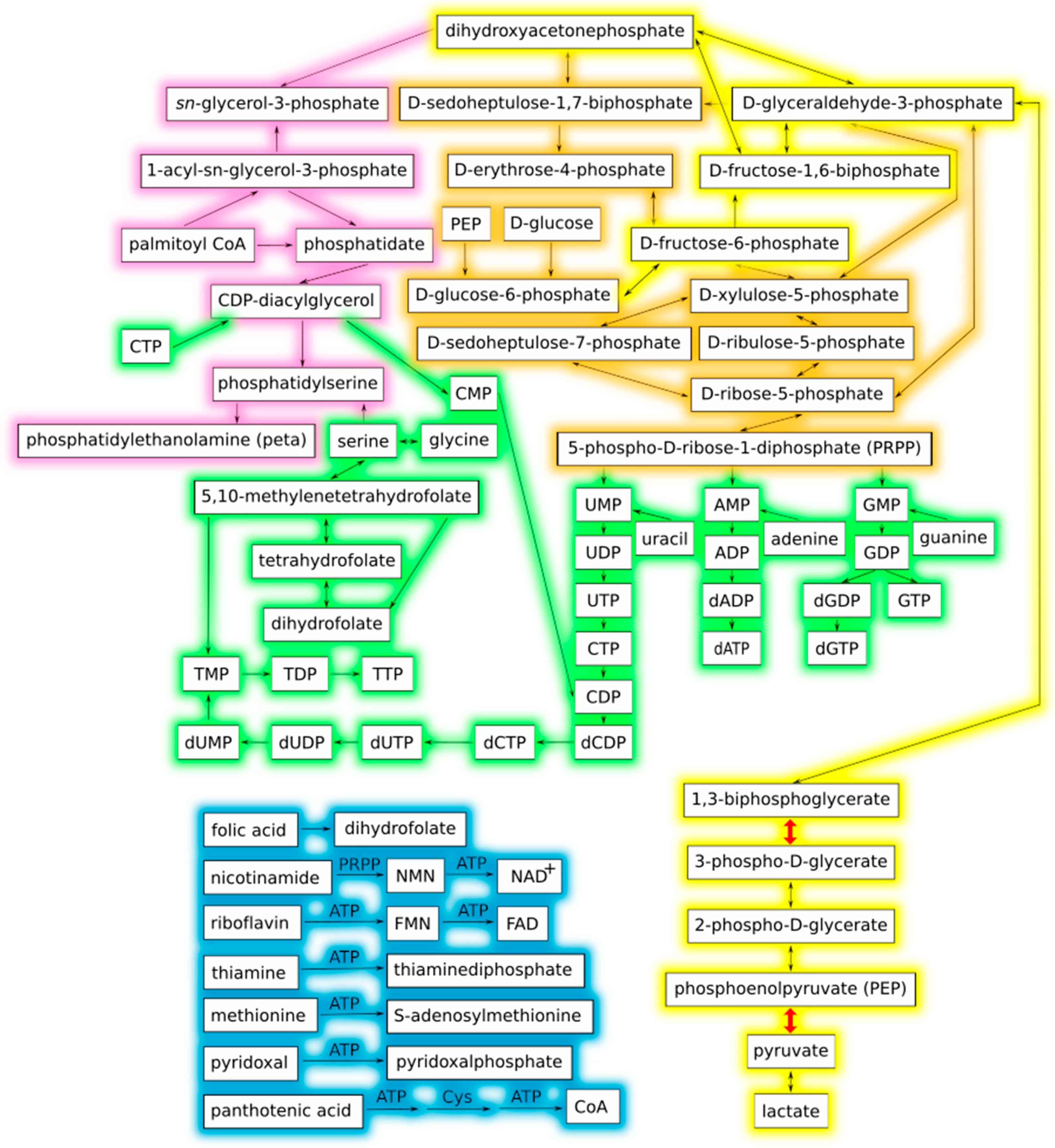
3.1. Theoretical Minimal Metabolic Network
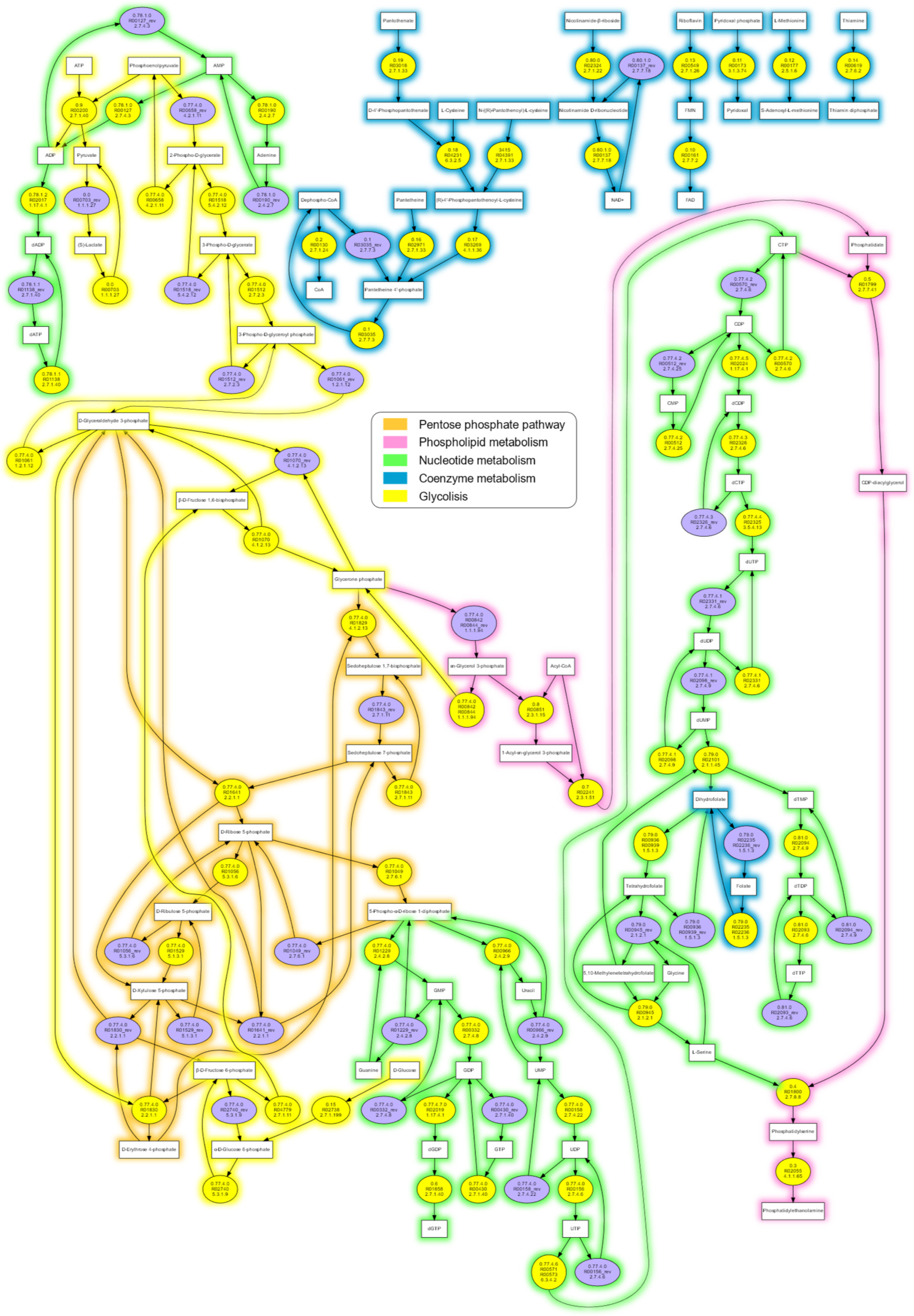
3.2. The MetaDAG Methodology: Analysis of the Composition and Connectivity of a Network at a Glance
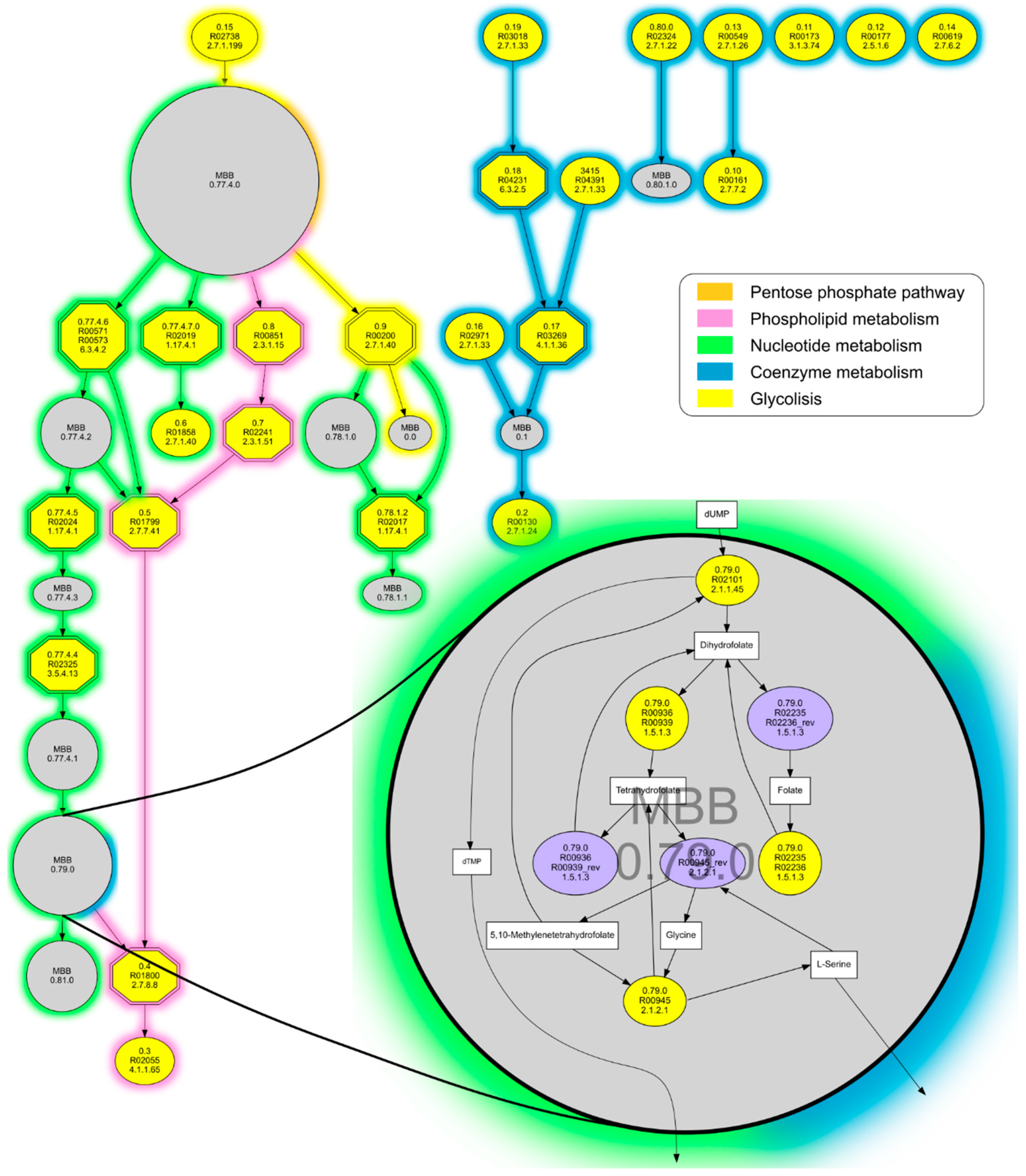
3.3. The m-DAG of “Candidatus Nasuia Deltocephalinicola”
3.4. The First Semisynthetic Viable Cell and Its m-DAG’s Reconstruction
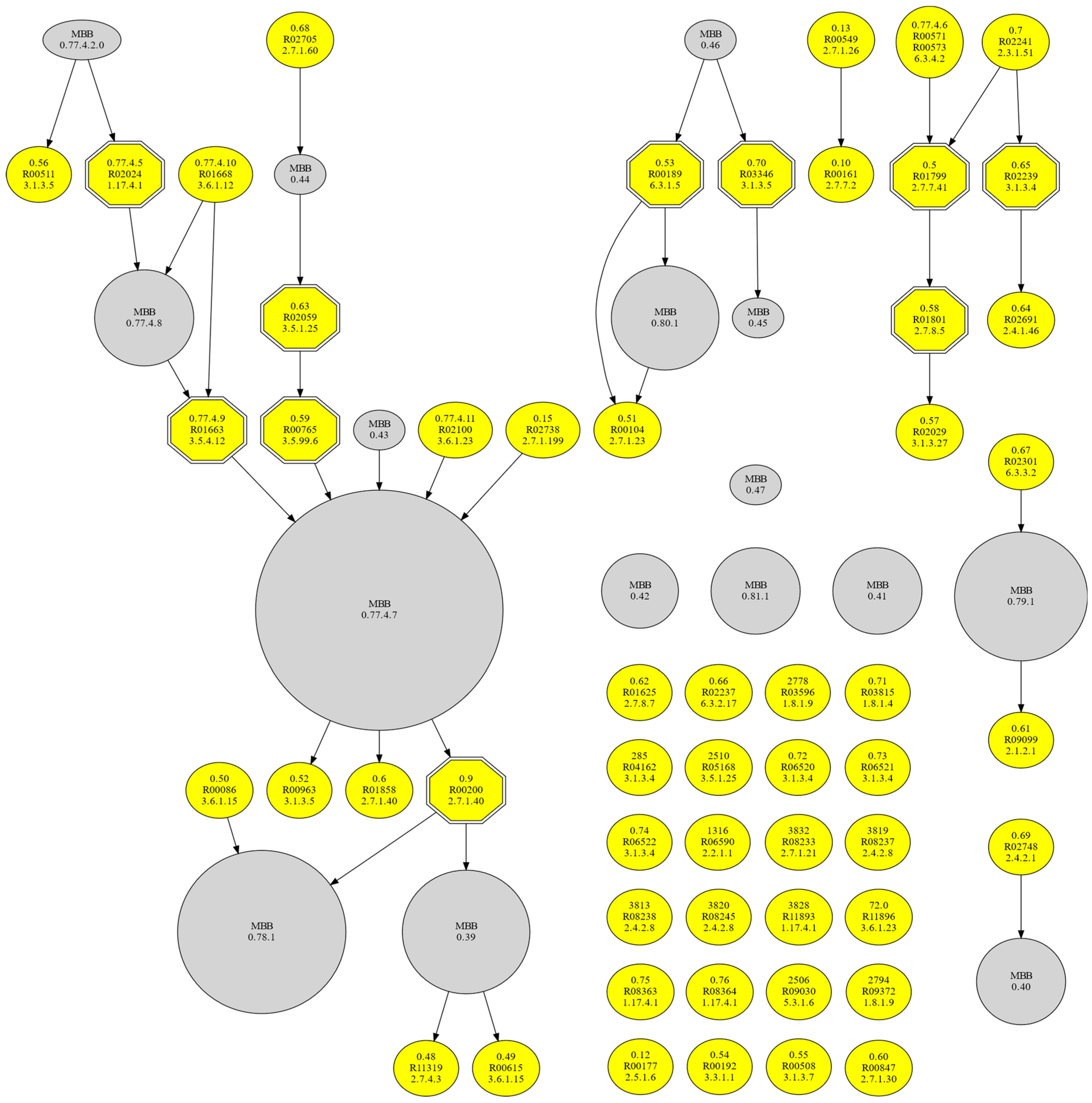
3.5. Resemblance of the MBBs of the Minimal m-DAGs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Moya, A.; Gil, R.; Latorre, A.; Peretó, J.; Garcillán-Barcia, M.P.; De La Cruz, F. Toward minimal bacterial cells: Evolution vs. design. FEMS Microbiol. Rev. 2009, 33, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luisi, P.L. Toward the engineering of minimal living cells. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2002, 268, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L.; Ferri, F.; Stano, P. Approaches to semi-synthetic minimal cells: A review. Naturwissenschaften 2005, 93, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.C.; Patil, K.R.; Rocha, I. Systems Biology Perspectives on Minimal and Simpler Cells. Microbiol. Mol. Biol. Rev. 2014, 78, 487–509. [Google Scholar] [CrossRef] [Green Version]
- Stano, P.; Luisi, P.L. On the Construction of Minimal Cell Models in Synthetic Biology and Origins of Life Studies. In Design and Analysis of Biomolecular Circuits; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2011; pp. 337–368. [Google Scholar]
- Bedau, M.A.; Parke, E.C.; Tangen, U.; Hantsche-Tangen, B. Social and ethical checkpoints for bottom-up synthetic biology, or protocells. Syst. Synth. Biol. 2009, 3, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Mantri, S.; Sapra, K.T. Evolving protocells to prototissues: Rational design of a missing link. Biochem. Soc. Trans. 2013, 41, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Stano, P. Is Research on Synthetic Cells. Moving to the Next Level? Life 2018, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Glass, J.I.; Merryman, C.; Wise, K.S.; Hutchison, C.A.; Smith, H.O. Minimal Cells—Real and Imagined. Cold Spring Harb. Perspect. Biol. 2017, 9, a023861. [Google Scholar] [CrossRef] [Green Version]
- Gil, R. The Minimal Gene-Set Machinery. In Encyclopedia of Molecular Cell Biology and Molecular Medicine; Wiley: Hoboken, NJ, USA, 2014; pp. 1–36. [Google Scholar]
- Luo, H.; Lin, Y.; Gao, F.; Zhang, Z.; Zhang, R. DEG 10, an update of the database of essential genes that includes both protein-coding genes and noncoding genomic elements: Table 1. Nucleic Acids Res. 2014, 42, D574–D580. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Rocha, C.G.; Fang, G.; Schmidt, M.; Ussery, D.W.; Danchin, A. From essential to persistent genes: A functional approach to constructing synthetic life. Trends Genet. 2013, 29, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Gil, R.; Silva, F.J.; Zientz, E.; Delmotte, F.; González-Candelas, F.; Latorre, A.; Rausell, C.; Kamerbeek, J.; Gadau, J.; Hölldobler, B.; et al. The genome sequence of Blochmannia floridanus: Comparative analysis of reduced genomes. Proc. Natl. Acad. Sci. USA 2003, 100, 9388–9393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mushegian, A.; Koonin, E.V. A minimal gene set for cellular life derived by comparison of complete bacterial genomes. Proc. Natl. Acad. Sci. USA 1996, 93, 10268–10273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, R.; Silva, F.J.; Peretó, J.; Moya, A. Determination of the Core of a Minimal Bacterial Gene Set. Microbiol. Mol. Biol. Rev. 2004, 68, 518–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewett, M.C.; Forster, A.C. Update on designing and building minimal cells. Curr. Opin. Biotechnol. 2010, 21, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Hutchison, C.; Chuang, R.-Y.; Noskov, V.N.; Assad-Garcia, N.; Deerinck, T.J.; Ellisman, M.H.; Gill, J.; Kannan, K.; Karas, B.J.; Ma, L.; et al. Design and synthesis of a minimal bacterial genome. Science 2016, 351, aad6253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venetz, J.E.; Del Medico, L.; Wölfle, A.; Schächle, P.; Bucher, Y.; Appert, D.; Tschan, F.; Flores-Tinoco, C.E.; Van Kooten, M.; Guennoun, R.; et al. Chemical synthesis rewriting of a bacterial genome to achieve design flexibility and biological functionality. Proc. Natl. Acad. Sci. USA 2019, 116, 8070–8079. [Google Scholar] [CrossRef] [Green Version]
- Gabaldón, T.; Peretó, J.; Montero, F.; Gil, R.; Latorre, A.; Moya, A. Structural analyses of a hypothetical minimal metabolism. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1751–1762. [Google Scholar] [CrossRef]
- Glass, J.I.; Assad-Garcia, N.; Alperovich, N.; Yooseph, S.; Lewis, M.R.; Maruf, M.; Iii, C.A.H.; Smith, H.O.; Venter, J.C. Essential genes of a minimal bacterium. Proc. Natl. Acad. Sci. USA 2006, 103, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Alberich, R.; Castro, J.A.; Llabrés, M.; Rodríguez, P.P. Metabolomics analysis: Finding out metabolic building blocks. PLoS ONE 2017, 12, e0177031. [Google Scholar] [CrossRef]
- Bennett, G.M.; Moran, N.A. Small, Smaller, Smallest: The Origins and Evolution of Ancient Dual Symbioses in a Phloem-Feeding Insect. Genome Biol. Evol. 2013, 5, 1675–1688. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Prieto, M.; Vargas-Chávez, C.; Llabrés, M.; Palmer, P.; Latorre, A.; Moya, A. An update on the Symbiotic Genomes Database (SymGenDB): A collection of metadata, genomic, genetic and protein sequences, orthologs and metabolic networks of symbiotic organisms. Database 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.F.; Georgellis, D. Bacterial Lipid Domains and Their Role in Cell Processes. In Biogenesis of Fatty Acids, Lipids and Membranes; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2019; pp. 575–592. [Google Scholar]
- Romano, A.; Conway, T. Evolution of carbohydrate metabolic pathways. Res. Microbiol. 1996, 147, 448–455. [Google Scholar] [CrossRef]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. B Boil. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef] [PubMed]
- Toft, C.; Andersson, S.G.E. Evolutionary microbial genomics: Insights into bacterial host adaptation. Nat. Rev. Genet. 2010, 11, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Vigneron, A.; Masson, F.; Vallier, A.; Balmand, S.; Rey, M.; Vincent-Monégat, C.; Aksoy, E.; Aubailly-Giraud, E.; Zaidman-Rémy, A.; Heddi, A. Insects Recycle Endosymbionts when the Benefit is Over. Curr. Biol. 2014, 24, 2267–2273. [Google Scholar] [CrossRef] [Green Version]
- Tamas, I. 50 Million Years of Genomic Stasis in Endosymbiotic Bacteria. Science 2002, 296, 2376–2379. [Google Scholar] [CrossRef] [Green Version]
- McCutcheon, J.P.; McDonald, B.R.; Moran, N.A. Convergent evolution of metabolic roles in bacterial co-symbionts of insects. Proc. Natl. Acad. Sci. USA 2009, 106, 15394–15399. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate KEGG ID | Reaction ID | Enzyme Name (E.C. Number) | Definition | Product KEGG ID | MBB ID |
|---|---|---|---|---|---|
| C00020 | R00127 r | adenylate kinase (2.7.4.3) | ATP + AMP ↔ 2 ADP | C00008 | 0.15 |
| C00882 | R00130 | dephospho-CoA kinase (2.7.1.24) | ATP + Dephospho-CoA → ADP + CoA | C00010 | 0.2 |
| C00455 | R00137 r | nicotinamide-nucleotide adenylyltransferase (2.7.7.1) | Diphosphate + NAD+ ↔ ATP + Nicotinamide d-ribonucleotide | C00003 | 0.80.1.0 |
| C00015 | R00156 r | nucleoside-diphosphate kinase (2.7.4.6) | ATP + UDP ↔ ADP + UTP | C00075 | 0.77.4.0 |
| C00105 | R00158 r | UMP/CMP kinase (2.7.4.14) | ATP + UMP ↔ ADP + UDP | C00015 | 0.77.4.0 |
| C00061 | R00161 | FAD synthase (2.7.7.2) | ATP + FMN → Diphosphate + FAD | C00016 | 0.10 |
| C00018 | R00173 | pyridoxal phosphatase (3.1.3.74) | Pyridoxal phosphate + H2O → Pyridoxal + Orthophosphate | C00250 | 0.11 |
| C00073 | R00177 | methionine adenosyltransferase (2.5.1.6) | ATP + l-Methionine + H2O → Orthophosphate + Diphosphate + S-Adenosyl-l-methionine | C00019 | 0.12 |
| C00020 + C0013 | R00190 r | adenine phosphoribosyltransferase (2.4.2.7) | AMP + Diphosphate ↔ Adenine + 5-Phospho-alpha-d-ribose 1-diphosphate | C00147 + C00119 | 0.78.1.0 |
| C00074 + C00008 | R00200 | pyruvate kinase (2.7.1.40) | ADP + Phosphoenolpyruvate → ATP + Pyruvate | C00022 | 0.9 |
| C00144 | R00332 r | guanylate kinase (2.7.4.8) | ATP + GMP ↔ ADP + GDP | C00035 | 0.77.4.0 |
| C00044 | R00430 r | pyruvate kinase (2.7.1.40) | GTP + Pyruvate ↔ GDP + Phosphoenolpyruvate | C00035 | 0.77.4.0 |
| C00055 | R00512 r | (d)CMP kinase (2.7.4.25) | ATP + CMP ↔ ADP + CDP | C00112 | 0.77.4.2 |
| C00255 | R00549 | riboflavin kinase (2.7.1.26) | ATP + Riboflavin → ADP + FMN | C00061 | 0.13 |
| C00112 | R00570 r | nucleoside diphosphate kinase (2.7.4.6) | ATP + CDP ↔ ADP + CTP | C00063 | 0.77.4.2 |
| C00075 | R00571, R00573 | CTP synthase (6.3.4.2) | ATP + UTP + Ammonia → ADP + Orthophosphate + CTP | C00063 | 0.77.4.6 |
| C00378 | R00619 | thiamine diphosphokinase (2.7.6.2) | ATP + Thiamine → AMP + Thiamin diphosphate | C00068 | 0.14 |
| C00631 | R00658 r | enolase (4.2.1.11) | 2-Phospho-d-glycerate ↔ Phosphoenolpyruvate + H2O | C00074 | 0.77.4.0 |
| C00186 | R00703 r | lactate dehydrogenase (1.1.1.27) | (S)-Lactate + NAD+ ↔ Pyruvate + NADH + H+ | C00022 | 0.0 |
| C00093 | R00842|R00844 r | sn-glycerol-3-phosphate dehydrogenase (1.1.1.94) | sn-Glycerol 3-phosphate + NAD+ ↔ Glycerone phosphate + NADH + H+ | C00111 | 0.77.4.0 |
| C00093 + C00040 | R00851 | arylamine N-acetyltransferase (2.3.1.15) | sn-Glycerol 3-phosphate + Acyl-CoA → 1-Acyl-sn-glycerol 3-phosphate + CoA | C00681 | 0.8 |
| C00415 | R00936|R00939 r | dihydrofolate reductase (1.5.1.3) | Dihydrofolate + NADH + H+ ↔ Tetrahydrofolate + NAD + | C00101 | 0.79.0 |
| C00037 + C00143 | R00945 r | glycine hydroxymethyltransferase (2.1.2.1) | 5,10-Methylenetetrahydrofolate + Glycine + H2O ↔ Tetrahydrofolate + l-Serine | C00065 + C00101 | 0.79.0 |
| C00105 | R00966 r | uracil phosphoribosyltransferase (2.4.2.9) | UMP + Diphosphate ↔ Uracil + 5-Phospho-alpha-d-ribose 1-diphosphate | C00106 + C00119 | 0.77.4.0 |
| C00117 | R01049 r | phosphoribosylpyrophosphate synthetase (2.7.6.1) | ATP + d-Ribose 5-phosphate ↔ AMP + 5-Phospho-alpha-d-ribose 1-diphosphate | C00119 | 0.77.4.0 |
| C00117 | R01056 r | ribose-5-phosphate isomerase (5.3.1.6) | d-Ribose 5-phosphate ↔ D-Ribulose 5-phosphate | C00199 | 0.77.4.0 |
| C00118 | R01061 r | glyceraldehyde-3-phosphate dehydrogenase (1.2.1.12) | d-Glyceraldehyde 3-phosphate + Orthophosphate + NAD+ ↔ 3-Phospho-d-glyceroyl phosphate + NADH + H+ | C00236 | 0.77.4.0 |
| C05378 | R01070 r | fructose-1,6-bisphosphate aldolase (4.1.2.13) | beta-d-Fructose 1,6-bisphosphate ↔ Glycerone phosphate + d-Glyceraldehyde 3-phosphate | C00111 + C00118 | 0.77.4.0 |
| C00131 | R01138 r | pyruvate kinase (2.7.1.40) | dATP + Pyruvate ↔ dADP + Phosphoenolpyruvate | C00206 | 0.78.1.1 |
| C00119 + C00242 | R01229 r | hypoxanthine phosphoribosyltransferase (2.4.2.8) | Guanine + 5-Phospho-alpha-d-ribose 1-diphosphate ↔ GMP + Diphosphate | C00144 | 0.77.4.0 |
| C00361 | R01858 | pyruvate kinase (2.7.1.40) | dGDP + Phosphoenolpyruvate → dGTP + Pyruvate | C00286 | 0.6 |
| C00008 | R02017 | ribonucleoside diphosphate reductase (1.17.4.1) | Thioredoxin + ADP → dADP + Thioredoxin disulfide + H2O | C00206 | 0.78.1.2 |
| C00035 | R02019 | ribonucleoside diphosphate reductase (1.17.4.1) | GDP + Thioredoxin → dGDP + Thioredoxin disulfide + H2O | C00361 | 0.77.4.7.0 |
| C00112 | R02024 | ribonucleoside diphosphate reductase (1.17.4.1) | Thioredoxin + CDP → dCDP + Thioredoxin disulfide + H2O | C00705 | 0.77.4.5 |
| C00197 | R01512 r | phosphoglycerate kinase (2.7.2.3) | ATP + 3-Phospho-d-glycerate ↔ ADP + 3-Phospho-d-glyceroyl phosphate | C00236 | 0.77.4.0 |
| C00631 | R01518 r | phosphoglycerate mutase (2,3-diphosphoglycerate-independent) (5.4.2.12) | 2-Phospho-d-glycerate ↔ 3-Phospho-d-glycerate | C00197 | 0.77.4.0 |
| C00199 | R01529 r | ribulose-phosphate 3-epimerase (5.1.3.1) | d-Ribulose 5-phosphate ↔ d-Xylulose 5-phosphate | C00231 | 0.77.4.0 |
| C00118 + C05382 | R01641 r | transketolase (2.2.1.1) | Sedoheptulose 7-phosphate + d-Glyceraldehyde 3-phosphate ↔ d-Ribose 5-phosphate + d-Xylulose 5-phosphate | C00117 + C00231 | 0.77.4.0 |
| C00063 + C00416 | R01799 r | phosphatidate cytidyltransferase (2.7.7.41) | CTP + Phosphatidate → Diphosphate + CDP-diacylglycerol | C00269 | 0.5 |
| C00065 + C00269 | R01800 | phosphatidylserine synthase (2.7.8.8) | -diacylglycerol + L-Serine → CMP + Phosphatidylserine | C00055 + C02737 | 0.4 |
| C00279 + C00111 | R01829 | fructose-1,6-bisphosphate aldolase (4.1.2.13) | Glycerone phosphate + d-Erythrose 4-phosphate → Sedoheptulose 1,7-bisphosphate | C00447 | 0.77.4.0 |
| C00118 + C05345 | R01830 r | transketolase (2.2.1.1) | beta-d-Fructose 6-phosphate + d-Glyceraldehyde 3-phosphate → d-Erythrose 4-phosphate + D-Xylulose 5-phosphate | C00231 + C00279 | 0.77.4.0 |
| C00363 | R02093 r | nucleoside diphosphate kinase (2.7.4.6) | ATP + dTDP ↔ ADP + dTTP | C00459 | 0.81.0 |
| C00364 | R02094 r | thymidine monophosphate kinase (2.7.4.9) | ATP + dTMP ↔ ADP + dTDP | C00363 | 0.81.0 |
| C00365 | R02098 r | thymidine monophosphate kinase (2.7.4.9) | ATP + dUMP ↔ ADP + dUDP | C01346 | 0.77.4.1 |
| C00143 + C00365 | R02101 | thymidylate synthase (2.1.1.45) | dUMP + 5,10-Methylenetetrahydrofolate → Dihydrofolate + dTMP | C00364 + C00415 | 0.79.0 |
| C00040 + C00681 | R02241 | 1-acyl-sn-glycerol-3-phosphate acyltransferase (2.3.1.51) | 1-Acyl-sn-glycerol 3-phosphate + Acyl-CoA → Phosphatidate + CoA | C00416 | 0.7 |
| C00458 | R02325 | dCTP deaminase (3.5.4.13) | dCTP + H2O → dUTP + Ammonia | C00460 | 0.77.4.4 |
| C00705 | R02326 r | nucleoside diphosphate kinase (2.7.4.6) | ATP + dCDP ↔ ADP + dCTP | C00458 | 0.77.4.3 |
| C01346 | R02331 r | nucleoside diphosphate kinase (2.7.4.6) | ATP + dUDP ↔ ADP + dUTP | C00460 | 0.77.4.1 |
| C02737 | R02055 | phosphatidylserine decarboxylase (4.1.1.65) | Phosphatidylserine → Phosphatidylethanolamine + CO2 | C00350 | 0.3 |
| C00504 | R02235|R02236 r | dihydrofolate reductase (1.5.1.3) | Folate + NADH + H+ ↔ Dihydrofolate + NAD+ | C00415 | 0.79.0 |
| C03150 | R02324 | ribosylnicotinamide kinase (2.7.1.22) | ATP + Nicotinamide-beta-riboside → ADP + Nicotinamide D-ribonucleotide | C00455 | 0.80.0 |
| C00031 | R02738 | protein-Npi-phosphohistidine---d-glucose phosphotransferase (2.7.1.199) | Protein N(pi)-phospho-L-histidine + D-Glucose → Protein histidine + alpha-d-Glucose 6-phosphate | C00668 | 0.15 |
| C00668 | R02740 r | glucose-6-phosphate isomerase (5.3.1.9) | alpha-d-Glucose 6-phosphate ↔ beta-d-Fructose 6-phosphate | C05345 | 0.77.4.0 |
| C00831 | R02971 | pantetheine kinase (2.7.1.34) | ATP + Pantetheine → ADP + Pantetheine 4’-phosphate | C01134 | 0.16 |
| C00864 | R03018 | pantothenate kinase (2.7.1.33) | ATP + Pantothenate → ADP + D-4’-Phosphopantothenate | C03492 | 0.19 |
| C01134 | R03035 r | pantetheine-phosphate adenylyltransferase (2.7.7.3) | ATP + Pantetheine 4’-phosphate → Diphosphate + Dephospho-CoA | C00882 | 0.1 |
| C03492 | R04231 r | phosphopantothenoylcysteine synthetase (6.3.2.5) | CTP + D-4’-Phosphopantothenate + l-Cysteine ↔ CMP + Diphosphate + (R)-4’-Phosphopantothenoyl-L-cysteine | C04352 | 0.18 |
| C04079 | R04391 r | pantothenate kinase (2.7.1.33) | ATP + N-((R)-Pantothenoyl)-L-cysteine ↔ ADP + (R)-4’-Phosphopantothenoyl-l-cysteine | C04352 | 3415 |
| C05345 | R04779 r | 6-phosphofructokinase (2.7.1.11) | ATP + beta-d-Fructose 6-phosphate ↔ ADP + beta-d-Fructose 1,6-bisphosphate | C05378 | 0.77.4.0 |
| C04352 | R03269 | phosphopantothenoylcysteine decarboxylase (4.1.1.36) | (R)-4’-Phosphopantothenoyl-L-cysteine → Pantetheine 4’-phosphate | C01134 | 0.17 |
| C05382 | R01843 r | 6-phosphofructokinase (2.7.1.11) | ATP + Sedoheptulose 7-phosphate ↔ ADP + Sedoheptulose 1,7-bisphosphate | C00447 | 0.77.4.0 |
| Reaction ID | Metabolic Pathway |
|---|---|
| R02019 | Purine metabolism |
| R02017 | |
| R00571/R00573 | Pyrimidine metabolism |
| R02024 | |
| R02325 | |
| R00851 | Glycerolipid metabolism, Glycerophospholipid metabolism |
| R02241 | |
| R01799 | Glycerophospholipid metabolism |
| R01800 | |
| R00200 | Glycolysis, part of the pyruvate metabolism |
| R04231 | Pantothenate and CoA biosynthesis |
| R03269 |
| Reaction ID | Metabolic Pathway |
|---|---|
| R09372 | Selenocompound metabolism |
| R00443 | Purine metabolism, Glycerophospholipid metabolism |
| R03012 | Histidine metabolism |
| R01163 | Histidine metabolism |
| R01288 | Cysteine and methionine metabolism, Sulfur metabolism |
| Reaction ID | Metabolic Pathway |
|---|---|
| R02024 | Pyrimidine metabolism |
| R01663 | |
| R02059 | Amino sugar and nucleotide sugar metabolism |
| R00765 | |
| R00200 | Glycolysis, part of the pyruvate metabolism |
| R00189 | Nicotinate and nicotinamide metabolism |
| R03346 | |
| R01799 | Glycerophospholipid metabolism |
| R01801 | |
| R02239 |
| Model Cell | MBB ID | # Reactions | Reaction ID |
|---|---|---|---|
| Minimal cell | 0.77.4.0 | 21 | R00156 (2.7.4.6) r, R00158 (2.7.4.22) r, R00332 (2.7.4.8) r, R00430 (2.7.1.40) r, R00658 (4.2.1.11) r, R00842|R00844 (1.1.1.94) r, R00966 (2.4.2.9) r, R01049 (2.7.6.1) r, R01056 (5.3.1.6) r, R01061 (1.2.1.12) r, R01070 (4.1.2.13) r, R01229 (2.4.2.8) r, R01512 (2.7.2.3) r, R01518 (5.4.2.12) r, R01529 (5.1.3.1) r, R01641 (2.2.1.1) r, R01829 (4.1.2.13), R01830 (2.2.1.1) r, R01843 (2.7.1.11) r, R02740 (5.3.1.9) r, R04779 (2.7.1.11) |
| 0.77.4.1 | 2 | R02098 (2.7.4.9) r, R02331 (2.7.4.6) r | |
| 0.77.4.2 | 2 | R00512 (2.7.4.25) r, R00570 (2.7.4.6) r | |
| 0.77.4.7.0 | 1 | R02019 (1.17.4.1) | |
| 0.78.1.0 | 2 | R00127 (2.7.4.3) r, R00190 (2.4.2.7) r | |
| 0.78.1.1 | 1 | R01138 (2.7.1.40) r | |
| 0.78.1.2 | 1 | R02017 (1.17.4.1) | |
| 0.79.0 | 4 | R00936|R00939 (1.5.1.3) r, R00945 (2.1.2.1) r, R02101 (2.1.1.45), R02235|R02236 (1.5.1.3) r | |
| 0.80.1.0 | 1 | R00137 (2.7.7.18) r | |
| 0.81.0 | 2 | R02093 (2.7.4.6) r, R02094 (2.7.4.9) r | |
| JCVI-syn 3.0 | 0.39 | 6 | R00014 (1.2.4.1), R00230 (2.3.1.8) r, R00315 (2.7.2.1) r, R02569 (2.3.1.12) r, R03270 (1.2.4.1), R07618 (1.8.1.4) r |
| 0.40 | 3 | R01126 (3.1.3.5), R01132 (2.4.2.8) r, R01863 (2.4.2.1) r | |
| 0.41 | 3 | R02142 (2.4.2.8) r, R02297 (2.4.2.1) r, R02719 (3.1.3.5) | |
| 0.42 | 2 | R00921 (2.3.1.8) r, R01353 (2.7.2.1) r | |
| 0.77.4.2.0 | 1 | R00512 (2.7.4.25) r | |
| 0.77.4.7 | 49 | R00158 (2.7.4.22) r, R00289 (2.7.7.9) r, R00291 (5.1.3.2) r, R00332 (2.7.4.8) r, R00430 (2.7.1.40) r, R00505 (5.4.99.9) r, R00658 (4.2.1.11) r, R00959 (5.4.2.5) r, R00966 (2.4.2.9) r, R01015 (5.3.1.1) r, R01049 (2.7.6.1)r, R01056 (5.3.1.6) r, R01057 (5.4.2.7) r, R01058 (1.2.1.9), R01061 (1.2.1.12) r, R01066 (4.1.2.4) r, R01067 (2.2.1.1), R01068 (4.1.2.13) r, R01070 (4.1.2.13) r, R01227 (3.1.3.5), R01229 (2.4.2.7)r, R01229 (2.4.2.8) r, R01512 (2.7.2.3) r, R01518 (5.4.2.12) r, R01529 (5.1.3.1) r, R01641 (2.2.1.1) r, R01819 (5.3.1.8) r, R01827 (2.2.1.2) r, R01829 (4.1.2.13), R01830 (2.2.1.1) r, R01843 (2.7.1.11) r, R01967 (2.7.1.113) r, R01968 (3.1.3.5)r, R01969 (2.4.2.1) r, R02018 (1.17.4.1), R02019 (1.17.4.1), R02090 (2.7.4.8) r, R02098 (2.7.4.9) r, R02099 (2.7.1.21), R02102 (3.1.3.5), R02102 (3.1.3.89), R02147 (2.4.2.1)r, R02484 (2.4.2.1), R02568 (4.1.2.13) r, R02739 (5.3.1.9) r, R02740 (5.3.1.9) r, R02749 (5.4.2.7) r, R03321 (5.3.1.9) r, R04779 (2.7.1.11) | |
| 0.77.4.8 | 5 | R01664 (3.1.3.5), R01664 (3.1.3.89), R01665 (2.7.4.25) r, R01666 (2.7.1.74), R01667 (3.6.1.12) | |
| 0.78.1 | 12 | R00127 (2.7.4.3) r, R00183 (3.1.3.5), R00185 (2.7.1.74) r, R00190 (2.4.2.7) r, R01138 (2.7.1.40) r, R01547 (2.7.4.11) r, R01547 (2.7.4.3) r, R01561 (2.4.2.1) r, R02017 (1.17.4.1), R02088 (3.1.3.5) r, R02089 (2.7.1.76), R02557 (2.4.2.1) r | |
| 0.79.1 | 6 | R00942 (6.3.2.17) r, R00945(2.1.2.1) r, R01220(1.5.1.5) r, R01655 (3.5.4.9) r, R03940 (2.1.2.9), R04241 (6.3.2.17) r | |
| 0.80.1 | 4 | R00137 (2.7.7.18) r, R01271 (2.4.2.12) r, R02294 (2.4.2.1) r, R02323 (3.1.3.5) | |
| 0.81.1 | 4 | R01567 (2.7.1.21), R01569 (3.1.3.5), R01569 (3.1.3.89), R02094 (2.7.4.9) r | |
| Nasuia | 0.77.0 | 2 | R00435 (2.7.7.6) r, R00441 (2.7.7.6) r |
| 0.78.0 | 2 | R00375 (2.7.7.7) r, R00376 (2.7.7.7) r |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Prieto, M.; Gil, R.; Llabrés, M.; Palmer-Rodríguez, P.; Moya, A. The Metabolic Building Blocks of a Minimal Cell. Biology 2021, 10, 5. https://doi.org/10.3390/biology10010005
Reyes-Prieto M, Gil R, Llabrés M, Palmer-Rodríguez P, Moya A. The Metabolic Building Blocks of a Minimal Cell. Biology. 2021; 10(1):5. https://doi.org/10.3390/biology10010005
Chicago/Turabian StyleReyes-Prieto, Mariana, Rosario Gil, Mercè Llabrés, Pere Palmer-Rodríguez, and Andrés Moya. 2021. "The Metabolic Building Blocks of a Minimal Cell" Biology 10, no. 1: 5. https://doi.org/10.3390/biology10010005
APA StyleReyes-Prieto, M., Gil, R., Llabrés, M., Palmer-Rodríguez, P., & Moya, A. (2021). The Metabolic Building Blocks of a Minimal Cell. Biology, 10(1), 5. https://doi.org/10.3390/biology10010005