Antibacterial Properties of Fucoidans from the Brown Algae Fucus vesiculosus L. of the Barents Sea

 ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
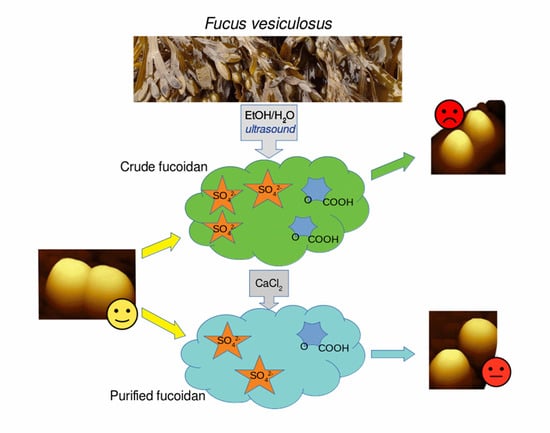
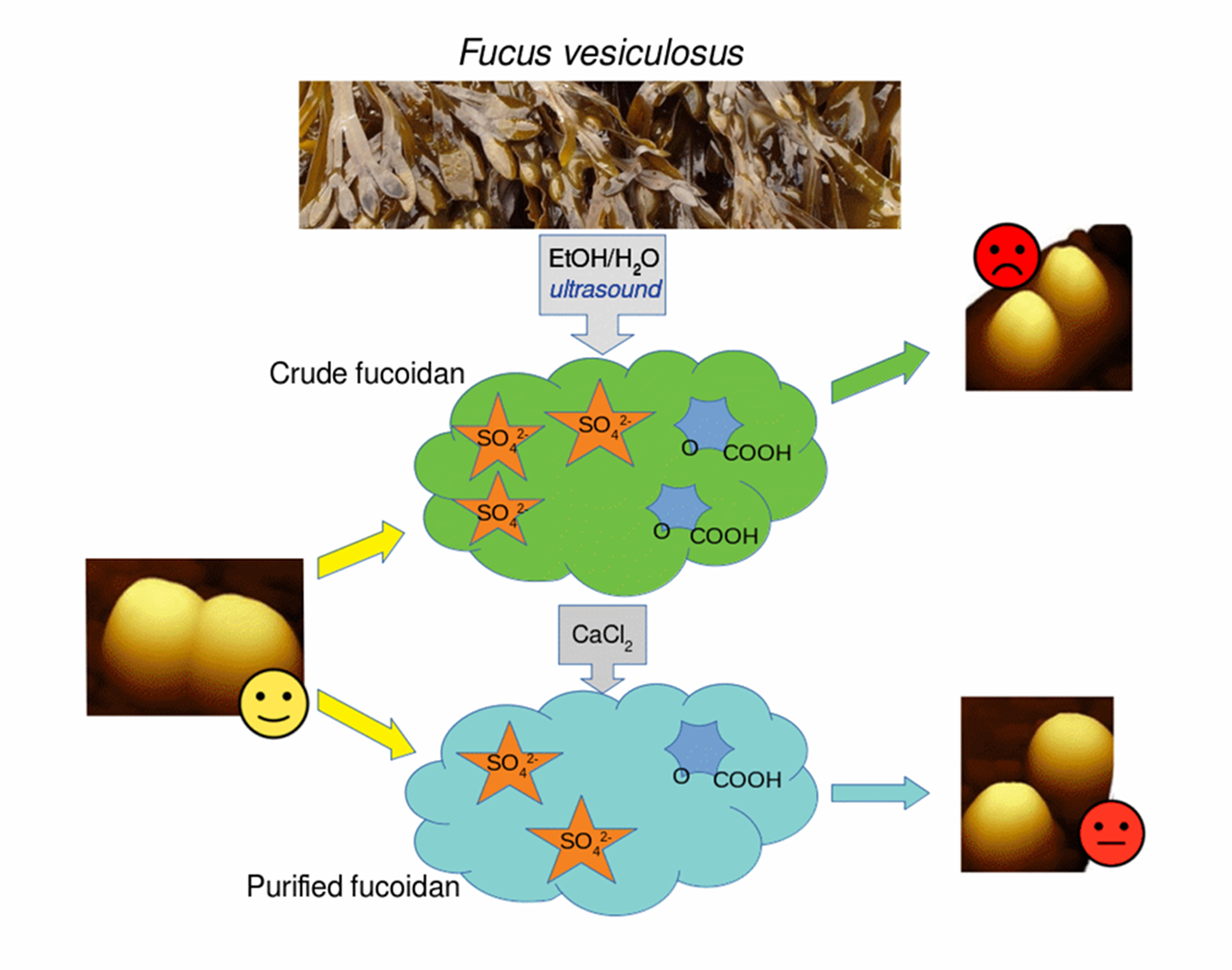
1. Introduction
2. Materials and Methods
3. Results
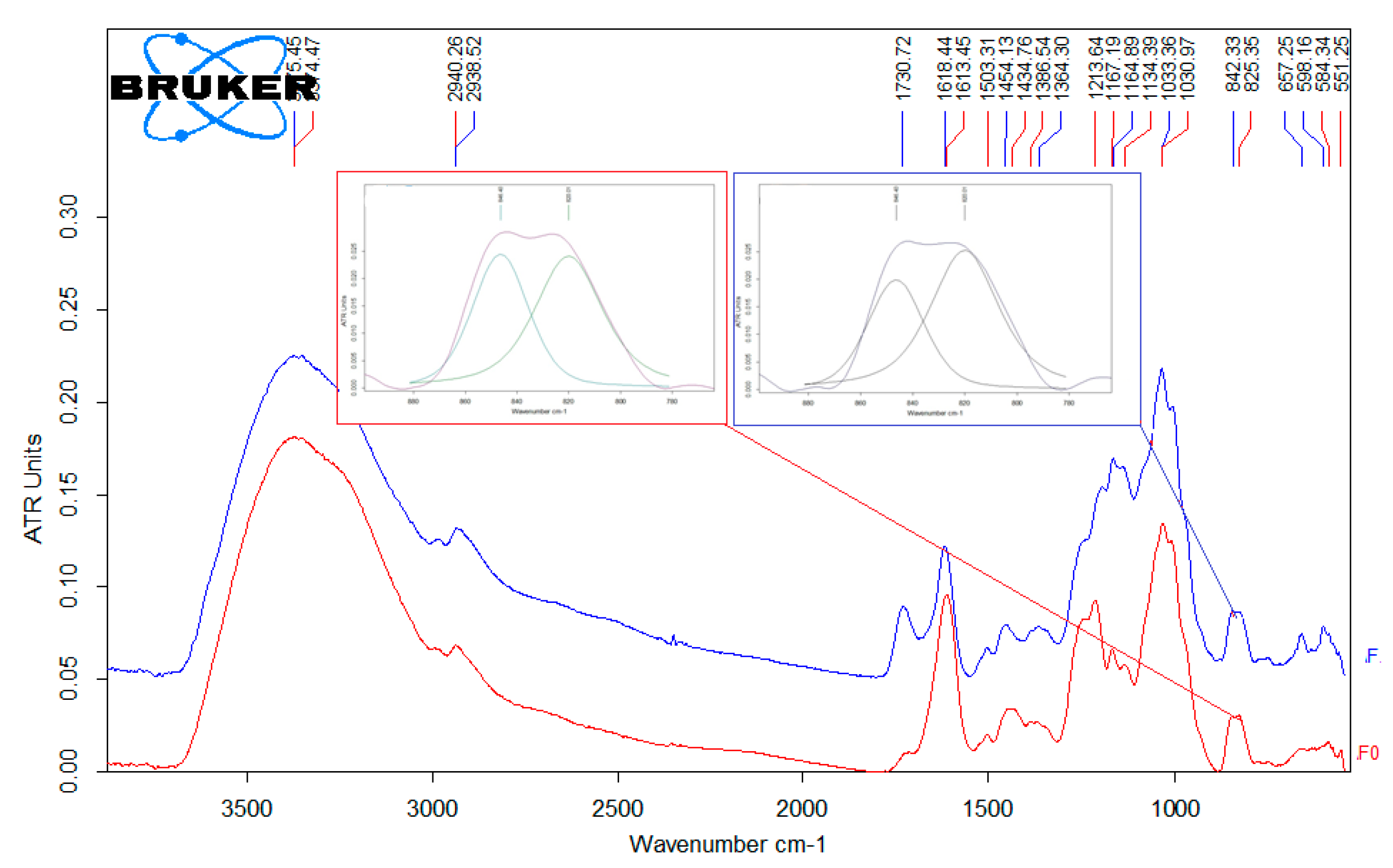
3.1. Characterization of Fucoidans
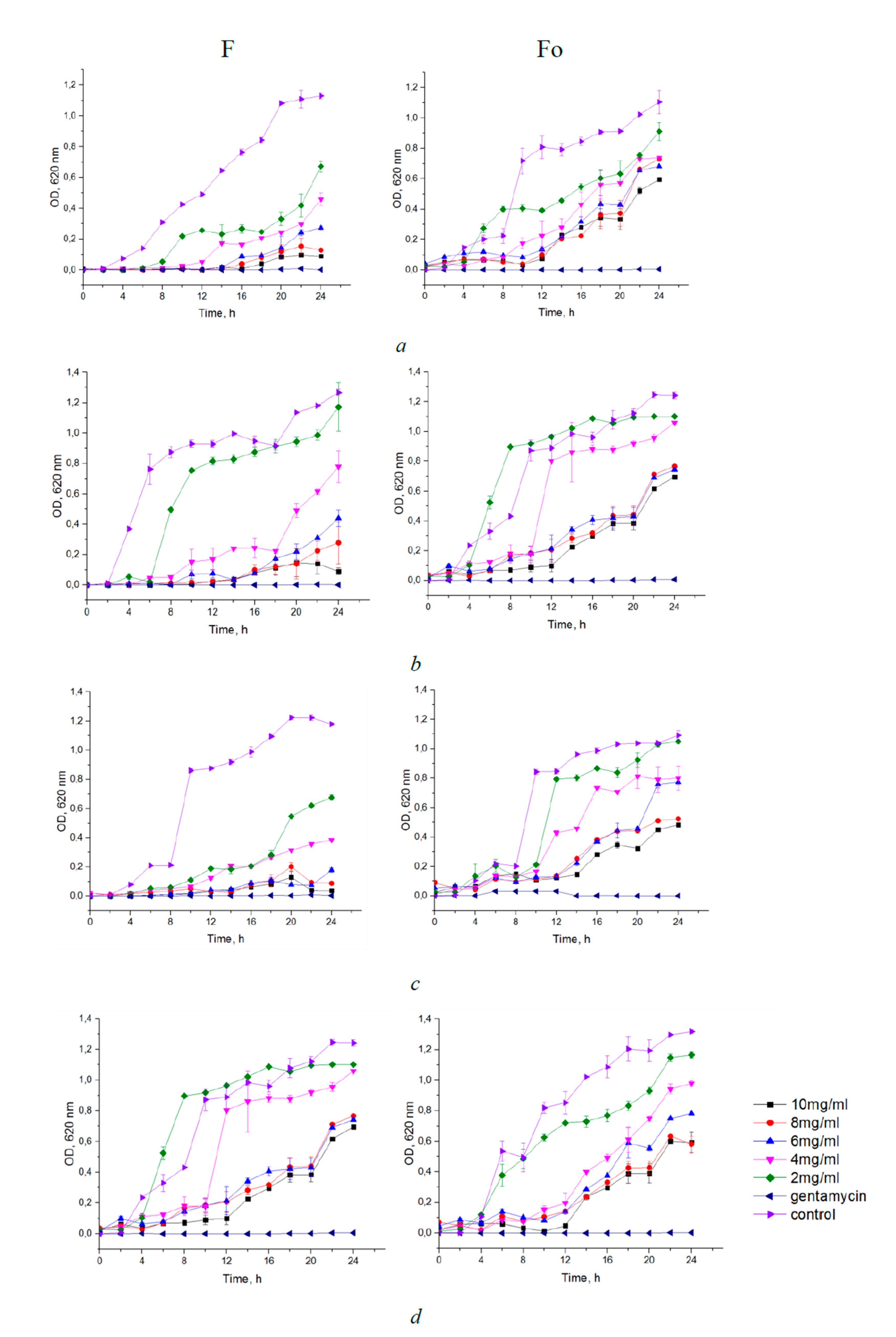
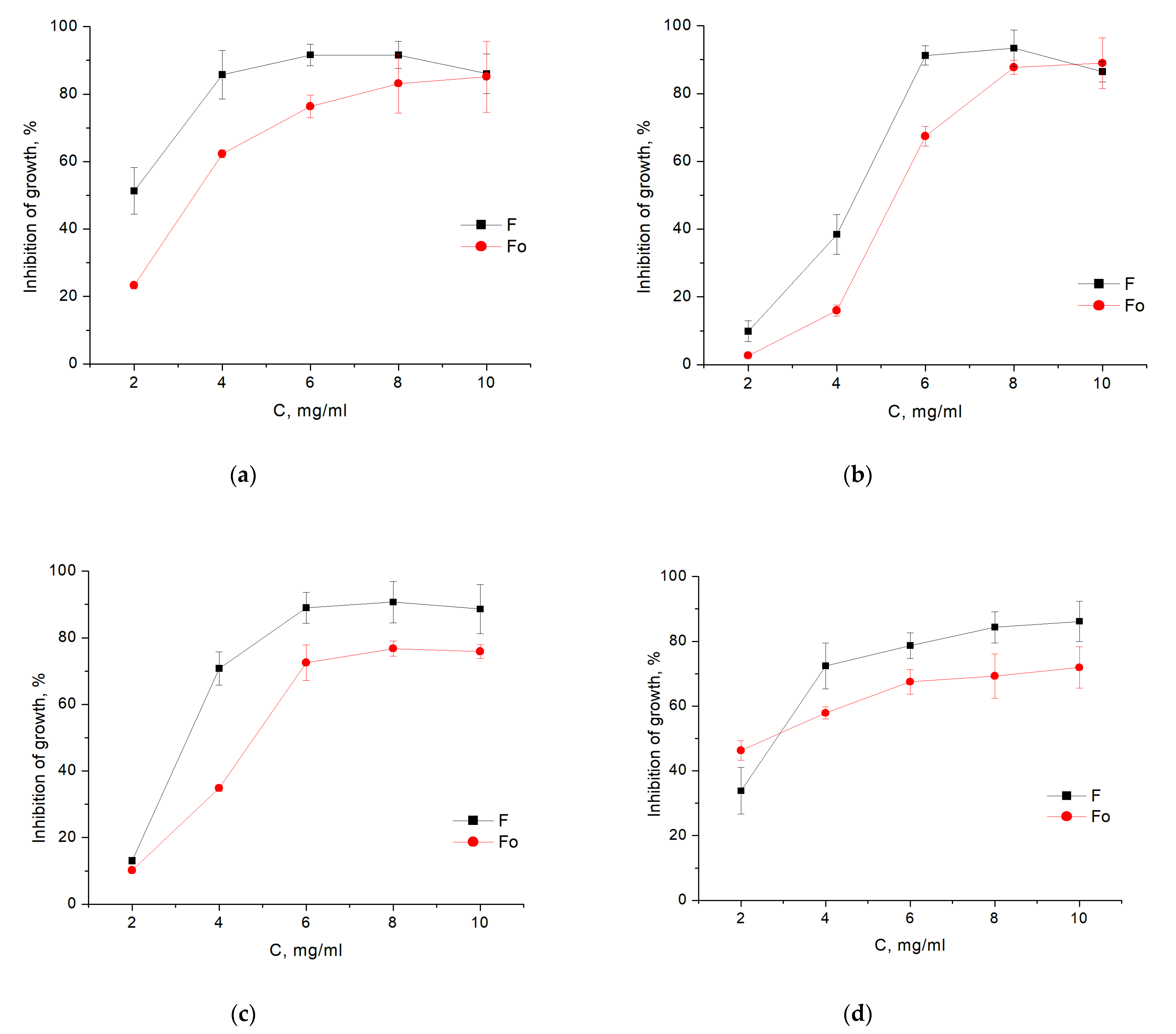
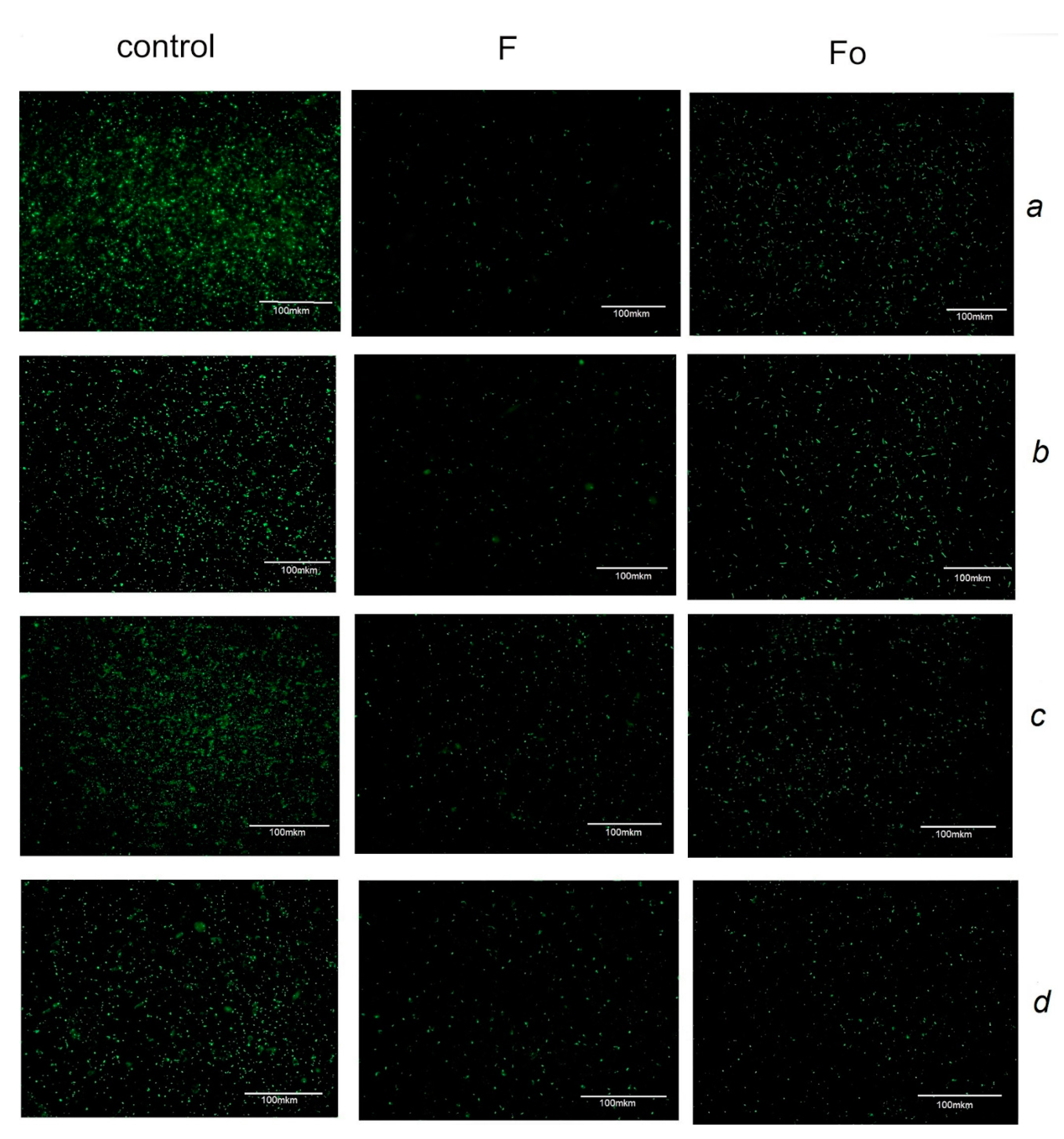
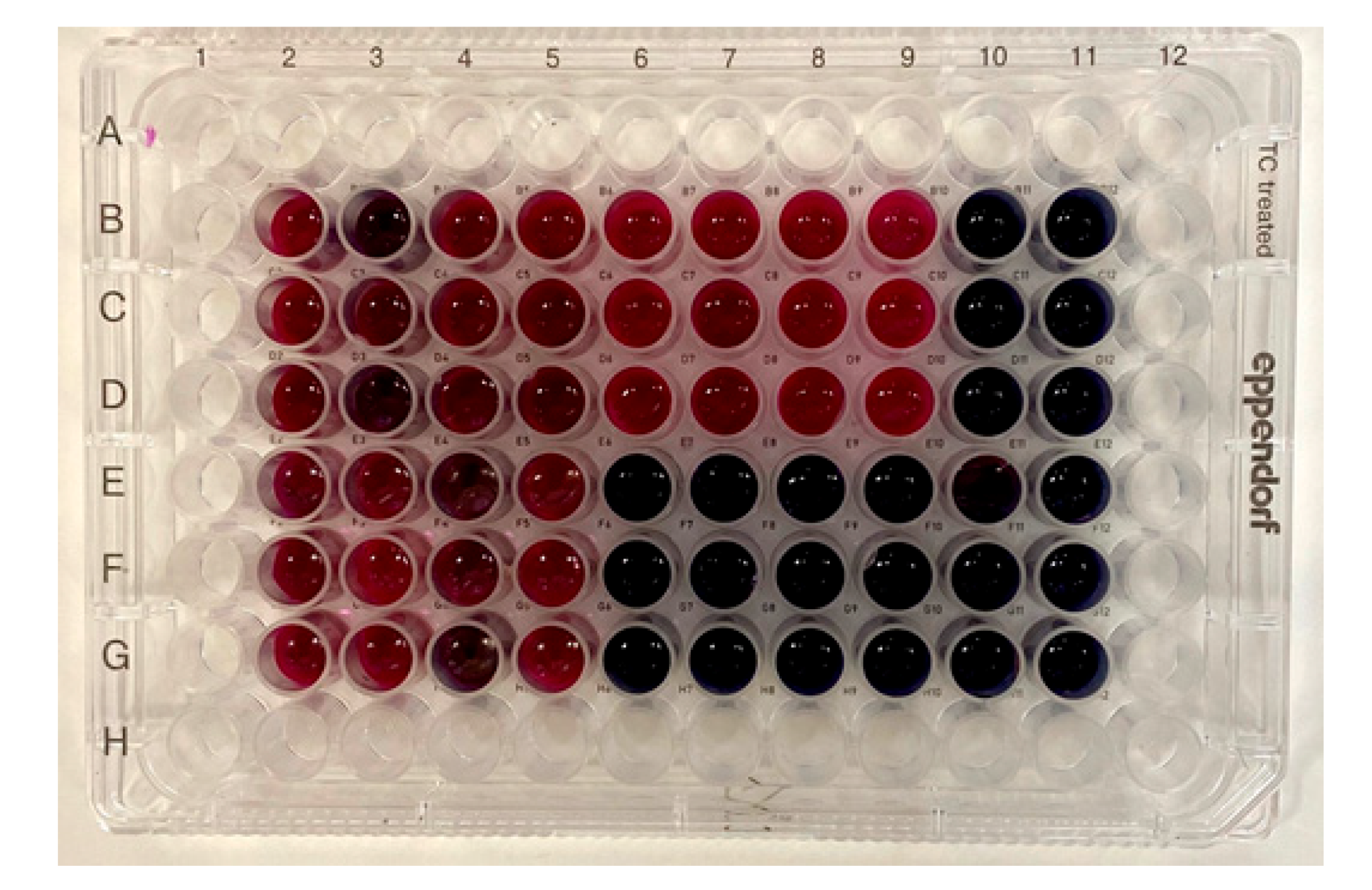
3.2. Antibacterial Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. Rev. Antimicrob. Resist. 2014, 20, 1–16. [Google Scholar]
- Who, Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 15 April 2020).
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morua, V.K.; Kim, J.; Kim, E.-K. Algal fucoidan: Structural and size-dependent bioactivities and their perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Zinoviev, E.V.; Lukyanov, S.A.; Kulminskaya, A.A.; Lapina, I.M.; Zhurishkina, E.V.; Lopatin, I.M.; Asadulaev, M.S.; Artsimovich, I.V.; Kostyakov, D.V.; Paneakh, M.B.; et al. Evaluation of the effectiveness of wound dressings based on bacterial cellulose with fucoidan for skin burns. Bull. Russ. Mil. Med. Acad. 2019, 1, 148–152. (In Russian) [Google Scholar]
- Patent RU 2 706 726 C1. Available online: https://yandex.ru/patents/doc/RU2706726C1_20191120 (accessed on 14 November 2020).
- Potoroko, I.Y.; Uskova, D.G.; Pajmulina, A.V.; Uday, B. Ultrasound micronization of fucoidan vegetable ingredient for the use in food production technology. Bull. South Ural. Univ. Ser. Food Biotechnol. 2019, 7, 58–70. [Google Scholar] [CrossRef]
- Malavenda, S.V.; Voskoboinikov, G.M. Influence of abiotic factors on the structure of the population of the brown alga Fucus vesiculosus in East Murman (Barents Sea). Russ. J. Mar. Biol. 2009, 35, 132–137. [Google Scholar] [CrossRef]
- Kılınç, B.; Cirik, S.; Turan, G. Seaweeds for Food and Industrial Applications. Food Ind. 2013, 31, 735–748. [Google Scholar]
- Luning, K. Seaweeds, Their Environment, Biogeography, and Ecophysiology, 1st ed.; Willey-Interscience: Cambridge, UK, 1990; p. 527. [Google Scholar]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Obluchinskaya, E.D. Comparative chemical composition of the Barents Sea brown algae. Appl. Biochem. Microbiol. 2008, 44, 305–309. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D. Influence of environmental factors on the content of polysaccharides in Fucus vesiculosus Fucus vesiculosus L. Chem. Plant Raw Mater. 2011, 3, 47–51. (In Russian) [Google Scholar]
- Tkach, A.V.; Obluchinskaya, E.D. Sterols and polyphenols of fucoids of the Murmansk coast of the Barents Sea. Bull. Murm. State Tech. Univ. 2017, 20, 326–335. [Google Scholar] [CrossRef]
- Parys, S.; Kehraus, S.; Pete, R.; Kupper, F.C.; Glombitza, K.W.; Konig, G.M. Seasonal variation of polyphenolics in Ascophyllum nodosum (Phaeophyceae). Eur. J. Phycol. 2009, 44, 331–338. (In Russian) [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of Bioactivities of Fucoidan from the Brown Seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Besednova, N.N.; Kuznetsova, T.A.; Zaporozhets, T.S.; Zvyagintseva, T.N. Sea brown algae is a source of new antibacterial pharmaceutical substances. Antibiot. Chemother. 2015, 60, 3–4. (In Russian) [Google Scholar]
- Patent RU 2 337 571(13) C2. Available online: https://patents.s3.yandex.net/RU2337571C2_20081110.pdf (accessed on 14 November 2020).
- Patent RU 2 506 089 C1. Available online: https://patentinform.ru/inventions/reg-2506089.html (accessed on 14 November 2020).
- Obluchinsksaya, E.D.; Makarova, M.N.; Pozharitskaya, O.N.; Shikov, A.N. Effects of ultrasound treatment on the chemical composition and anticoagulant properties of dry fucus extract. Pharm. Chem. J. 2015, 49, 183–186. [Google Scholar] [CrossRef]
- Zayed, A.; Muffler, K.; Hahn, T.; Rupp, S.; Finkelmeier, D.; Burger-Kentischer, A.; Ulber, R. Physicochemical and biological characterization of fucoidan from Fucus vesiculosus purified by dye affinity chromatography. Mar. Drugs 2016, 14, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Extraction of sulfated polysaccharides by autohydrolysis of brown seaweed Fucus vesiculosus. J. Appl. Phycol. 2013, 21, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dodgson, K.S. Determination of inorganic sulphate in studies on the enzymatic and non-enzymatic hydrolysis of carbohydrate and other sulphate esters. Biochem. J. 1961, 78, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Dische, Z.; Shettles, L.B.; Osnos, M. New specific color reactions of hexoses and spectrophotometric micro-metods for their determination. Arch. Biochem. 1949, 22, 169–184. [Google Scholar]
- Usov, A.I.; Smirnova, G.P.; Klochkova, N.G. Polysaccharide Composition of Several Brown Algae from Kamchatka. Russ. J. Bioorganic Chem. 2001, 27, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Gall, E. Phenolic Compounds in the Brown Seaweed Ascophyllum nodosum: Distribution and Radical-scavenging Activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Golovchenko, V.V.; Bushneva, O.A.; Ovodova, R.G.; Shashkov, A.S.; Chizhov, A.O.; Ovodov, Y.S. Structural study of bergenan, a pectin from Bergenia crassifolia. Russ. J. Bioorganic Chem. 2007, 33, 47–56. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://clsi.org/media/1469/m100s27_sample.pdf (accessed on 30 October 2020).
- Palanisamy, S.; Vinosha, M.; Rajasekar, P.; Anjali, R.; Sathiyaraj, G.; Marudhupandi, T.; Selvam, S.; Prabhu, N.M.; You, S. Antibacterial efficacy of a fucoidan fraction (Fu-F2) extracted from Sargassum polycystum. Int. J. Biol. Macromol. 2019, 125, 485–495. [Google Scholar] [CrossRef]
- Satyajit, D.; Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar]
- Rossetto, G.; Bergese, P.; Colombi, P.; Depero, L.; Giuliani, A.; Nicoletto, S.F.; Pirri, G. Atomic force microscopy evaluation of the effects of a novel antimicrobial multimeric peptide on Pseudomonas aeruginosa. Nanomed. Nanotechnol. Biol. Med. 2007, 3, 198–207. [Google Scholar] [CrossRef]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, H.R.; Biller, P.; Ross, A.B.; Adams, J.M.M. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Obluchinskaya, E.D.; Voskoboinikov, G.M.; Galynkin, V.A. Contents of alginic acid and fucoidan in Fucus algae of the Barents sea. Appl. Biochem. Microbiol. 2002, 38, 186–188. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Zaharova, L.V. Comparative study of polyphenols of brown algae of the Barents sea and the White sea, as well as the waters of the North Atlantic. Chem. Plant Raw Mater. 2020, 4. (In Russian) [Google Scholar] [CrossRef]
- Urvantseva, A.M.; Bakunina, I.Y.; Kim, N.Y.; Isakov, V.V.; Glazunov, V.P.; Zvyagintseva, T.N. Isolation of purified fucoidan from the natural complex with polyphenols and its characterisics. Chem. Veg. Raw Mater. 2004, 3, 15–24. [Google Scholar]
- Zhurishkina, E.V.; Stepanov, S.I.; Ayrapetyan, O.N.; Skorik, Y.A.; Vlasova, E.N.; Kruchina-Bogdanov, I.V.; Lebedev, D.V.; Kulminskaya, A.A.; Lapina, I.M. The effect of polydisperse fucoidans from Fucus vesiculosus on Hep G2 and Chang liver cells. Bioact. Carbohydr. Diet. Fibre 2020, 21, 100209. [Google Scholar] [CrossRef]
- Jun, J.Y.; Jung, M.J.; Jeong, I.H.; Yamazaki, K.; Kawai, Y.; Kim, B.M. Antimicrobial and Antibiofilm Activities of Sulfated Polysaccharides from Marine Algae against Dental Plaque Bacteria. Mar Drugs. 2018, 16, 301. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Liu, Y.; Cao, M.J.; Liu, G.M.; Chen, Q.; Sun, L.; Chen, H. Antibacterial activity and mechanisms of depolymerized fucoidans isolated from Laminaria japonica. Carbohydr Polym. 2017, 172, 294–305. [Google Scholar] [CrossRef]
- Ashayerizadeh, O.; Dastar, B.; Pourashouri, P. Study of antioxidant and antibacterial activities of depolymerized fucoidans extracted from Sargassum tenerrimum. Int. J. Biol. Macromol. 2019, 151, 1259–1266. [Google Scholar] [CrossRef] [PubMed]
- Gushchina, Y.Y.; Olyunina, L.N.; Goncharova, T.A.; Veselov, A.P.; Maikova, Y.A.; Ezhevskaya, M.A. Study of cell surface morphology azotobacter chrococcum in hyperthermia by method of atomic force microscopy Surface. X-RaySynchrotronNeutron Stud. 2005, 5, 87–92. (In Russian) [Google Scholar]
- Eaton, P.; Fernandes, J.C.; Pereira, E.; Pintado, M.E.; Malcata, F.X. Atomic force microscopy study of the antibacterial effects of chitosans on Escherichia coli and Staphylococcus aureus. Ultramicroscopy 2008, 108, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Donnenberg, M. Escherichia coli, 1st ed.; Academic Press: Cambridge, MA, USA, 2002; pp. 3–53. [Google Scholar]
- Yamashita, S.; Sugita-Konishi, Y.; Shimizu, M. In vitro Bacteriostatic Effects of Dietary Polysaccharides. Food Sci. Technol. Res. 2001, 7, 262–264. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fucoidan | Neutral Monosaccharides, mol% | Proteins, % | Sulfates, % | Uronic Acids, % | Fucose, % | Polyphenols, % | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fucose | Xylose | Mannose | Glucose | Galactose | ||||||
| F | 59.9 | 11.1 | 4.3 | 13.6 | 11.1 | 0 | 27.6 ± 0.2 | 9.2 ± 0.5 | 28.7 ± 0.4 | 6.8 ± 0.4 |
| Fo | 43.1 | 24.3 | 1.7 | 8.9 | 22.0 | 0 | 18.3 ± 0.3 | 5.1 ± 0.6 | 38.6 ± 0.7 | 6.5 ± 0.6 |
| Fucoidan | Peak | Mw, kDa | Area, % |
|---|---|---|---|
| F | 1 | 2500–2600 | 3.0 |
| 2 | 750–850 | 21.0 | |
| 3 | 200–250 | 50.2 | |
| 4 | 70–80 | 14.8 | |
| 5 | 10–15 | 11.0 | |
| Fo | 1 | 1150–1250 | 11.5 |
| 2 | 175–185 | 65.6 | |
| 3 | 10–20 | 22.0 | |
| 4 | 3.5–8 | 0.9 |
| Fraction | E. coli | B. licheniformis | S. aureus | S. epidermidis |
|---|---|---|---|---|
| F | 4 | 6 | 4 | 4 |
| Fo | 4 | 6 | 6 | 6 |
| Microorganisms | Control Ra, nm | F Ra, nm | Fo Ra, nm |
|---|---|---|---|
| E. coli | 1.5 ± 0.3 | 2.1 ± 0.3 | 1.4 ± 0.2 † |
| B. licheniformis | 1.2 ± 0.1 | 1.6 ± 0.1 ** | 2.0 ± 0.2 ** |
| S. aureus | 1.3 ± 0.3 | 2.2 ± 0.2 * | 2.5 ± 0.3 ** |
| S. epidermidis | 2.3 ± 0.4 | 4.1 ± 0.3 ** | 2.6 ± 0.5 † |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayrapetyan, O.N.; Obluchinskaya, E.D.; Zhurishkina, E.V.; Skorik, Y.A.; Lebedev, D.V.; Kulminskaya, A.A.; Lapina, I.M. Antibacterial Properties of Fucoidans from the Brown Algae Fucus vesiculosus L. of the Barents Sea. Biology 2021, 10, 67. https://doi.org/10.3390/biology10010067
Ayrapetyan ON, Obluchinskaya ED, Zhurishkina EV, Skorik YA, Lebedev DV, Kulminskaya AA, Lapina IM. Antibacterial Properties of Fucoidans from the Brown Algae Fucus vesiculosus L. of the Barents Sea. Biology. 2021; 10(1):67. https://doi.org/10.3390/biology10010067
Chicago/Turabian StyleAyrapetyan, Olga N., Ekaterina D. Obluchinskaya, Elena V. Zhurishkina, Yury A. Skorik, Dmitry V. Lebedev, Anna A. Kulminskaya, and Irina M. Lapina. 2021. "Antibacterial Properties of Fucoidans from the Brown Algae Fucus vesiculosus L. of the Barents Sea" Biology 10, no. 1: 67. https://doi.org/10.3390/biology10010067
APA StyleAyrapetyan, O. N., Obluchinskaya, E. D., Zhurishkina, E. V., Skorik, Y. A., Lebedev, D. V., Kulminskaya, A. A., & Lapina, I. M. (2021). Antibacterial Properties of Fucoidans from the Brown Algae Fucus vesiculosus L. of the Barents Sea. Biology, 10(1), 67. https://doi.org/10.3390/biology10010067