
Zinc Chloride: Time-Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of ZnCl2 Stock Solution
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Proliferation Assay
2.6. Gene Expression
2.7. Statistical Analysis
3. Results and Discussion
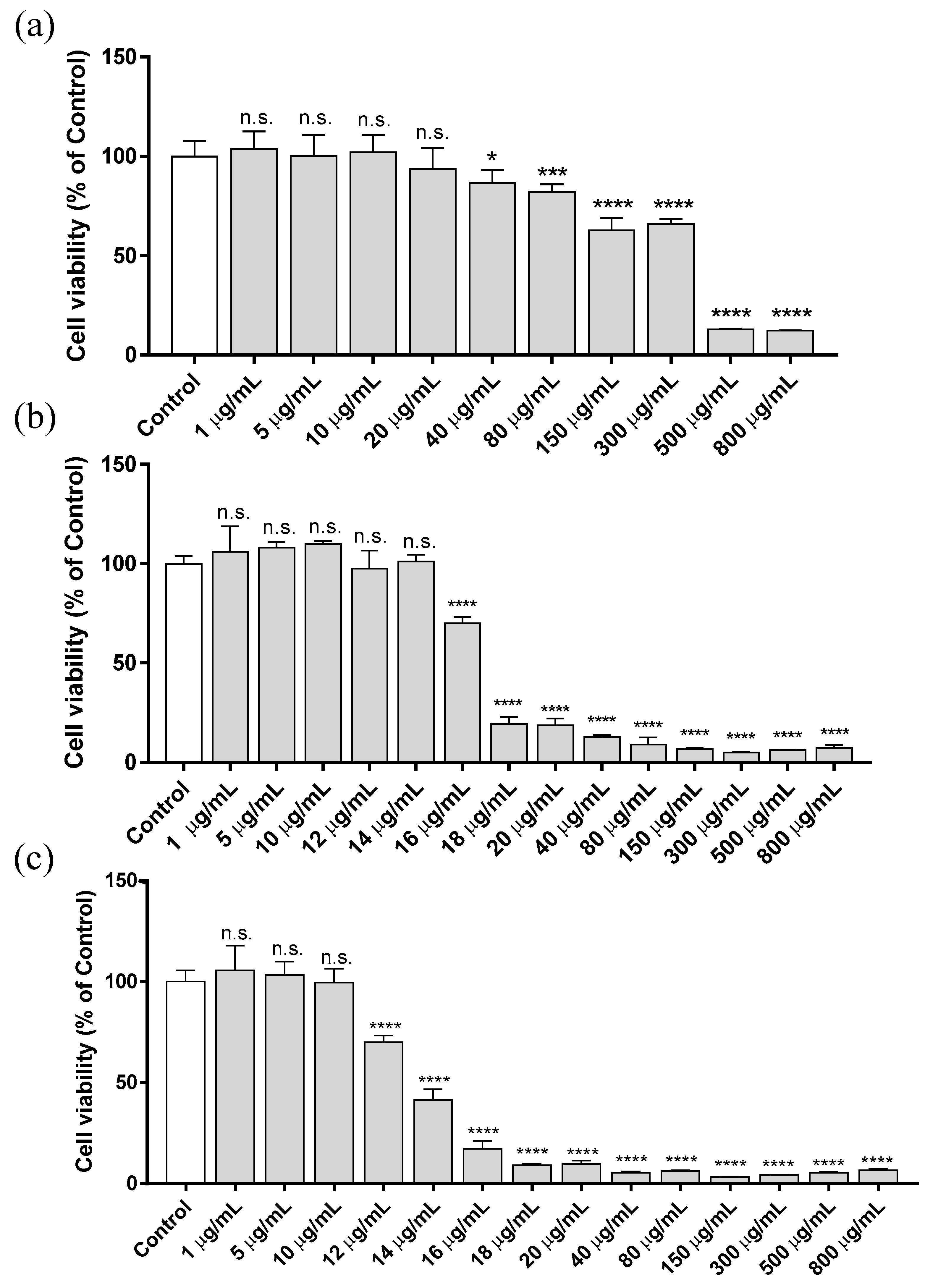
3.1. Cytotoxicity Assay
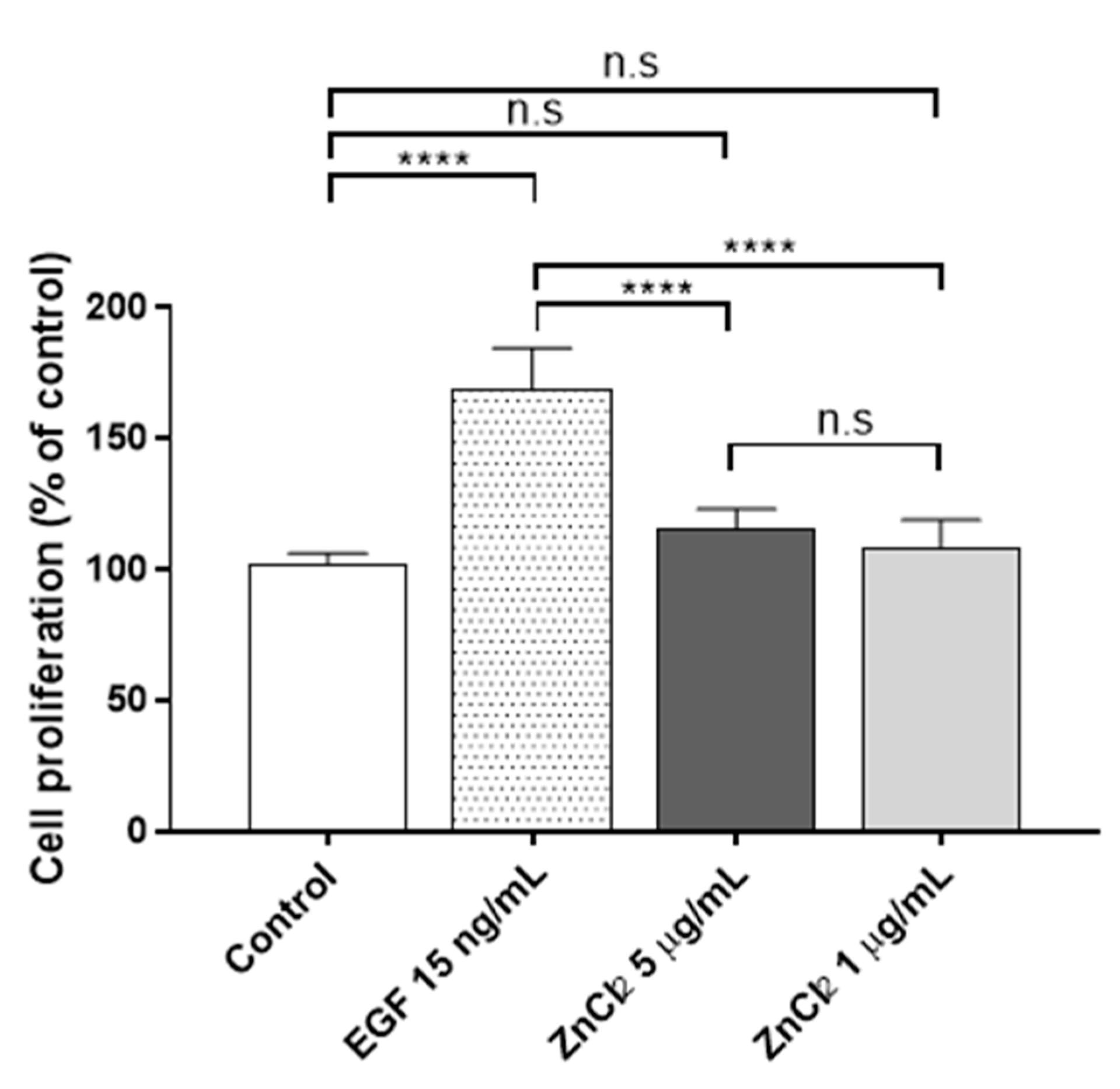
3.2. Proliferation Assay
3.3. Gene Expression
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beyersmann, D.; Haase, H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. BioMetals 2001, 14, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Kambe, T. (Eds.) Zinc Signaling; Springer Nature Singapore Pte Ltd.: Singapore, 2019; ISBN 9789811505577. [Google Scholar]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: Molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. 2017, 67, 283–301. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kawamura, T.; Shimada, S. Zinc and skin biology. Arch. Biochem. Biophys. 2016, 611, 113–119. [Google Scholar] [CrossRef]
- Jackson, M.J. (Ed.) Physiology of Zinc: General Aspects; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kinoshita, M.; Shimada, S.; Kawamura, T. Zinc and skin disorders. Nutrients 2018, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani, D.; Moztarzadeh, F.; Bizari, D. Alginate-bioactive glass containing Zn and Mg composite scaffolds for bone tissue engineering. Int. J. Biol. Macromol. 2019, 137, 1256–1267. [Google Scholar] [CrossRef] [PubMed]
- Neščáková, Z.; Zheng, K.; Liverani, L.; Nawaz, Q.; Galusková, D.; Kaňková, H.; Michálek, M.; Galusek, D.; Boccaccini, A.R. Multifunctional zinc ion doped sol–gel derived mesoporous bioactive glass nanoparticles for biomedical applications. Bioact. Mater. 2019, 4, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salcedo, S.; Shruti, S.; Salinas, A.J.; Malavasi, G.; Menabue, L.; Vallet-Regí, M. In vitro antibacterial capacity and cytocompatibility of SiO 2-CaO-P2O5 meso-macroporous glass scaffolds enriched with ZnO. J. Mater. Chem. B 2014, 2, 4836–4847. [Google Scholar] [CrossRef]
- Paramita, P.; Ramachandran, M.; Narashiman, S.; Nagarajan, S.; Sukumar, D.K.; Chung, T.W.; Ambigapathi, M. Sol–gel based synthesis and biological properties of zinc integrated nano bioglass ceramics for bone tissue regeneration. J. Mater. Sci. Mater. Med. 2021, 32, 5. [Google Scholar] [CrossRef]
- Mouriño, V.; Cattalini, J.P.; Boccaccini, A.R. Metallic ions as therapeutic agents in tissue engineering scaffolds: An overview of their biological applications and strategies for new developments. J. R. Soc. Interface 2012, 9, 401–419. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Jia, B.; Zhang, Z.; Qu, X.; Li, G.; Lin, W.; Zhu, D.; Dai, K.; Zheng, Y. Alloying design of biodegradable zinc as promising bone implants for load-bearing applications. Nat. Commun. 2020, 11, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, G.K.; Goldman, J.; Aghion, E. The prospects of zinc as a structural material for biodegradable implants—A review paper. Metals 2017, 7, 402. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.H.; Sermersheim, M.; Li, H.; Lee, P.H.U.; Steinberg, S.M.; Ma, J. Zinc in wound healing modulation. Nutrients 2018, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surf. A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Frígols, B.; Martí, M.; Salesa, B.; Hernández-Oliver, C.; Aarstad, O.; Ulset, A.-S.T.; Sætrom, G.I.; Aachmann, F.L.; Serrano-Aroca, Á. Graphene oxide in zinc alginate films: Antibacterial activity, cytotoxicity, zinc release, water sorption/diffusion, wettability and opacity. PLoS ONE 2019, 14, e0212819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rembe, J.D.; Boehm, J.K.; Fromm-Dornieden, C.; Hauer, N.; Stuermer, E.K. Comprehensive analysis of zinc derivatives pro-proliferative, anti-apoptotic and antimicrobial effect on human fibroblasts and keratinocytes in a simulated, nutrient-deficient environment in vitro. Int. J. Mol. Cell. Med. 2020, 9, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Tabata, Y. Biomaterial technology for tissue engineering applications. J. R. Soc. Interface 2009, 6, S311–S324. [Google Scholar] [CrossRef] [Green Version]
- Laurenti, M.; Cauda, V. ZnO nanostructures for tissue engineering applications. Nanomaterials 2017, 7, 374. [Google Scholar] [CrossRef] [Green Version]
- Sabater i Serra, R.; Molina-mateo, J.; Torregrosa-cabanilles, C.; Andrio-Balado, A.; Meseguer Dueñas, J.; Serrano-Aroca, A. Bio-Nanocomposite Hydrogel Based on Zinc Conformation, Thermal Behavior/Degradation, and Dielectric Properties. Polymers 2020, 12, 702. [Google Scholar] [CrossRef] [Green Version]
- Tiffany, A.S.; Gray, D.L.; Woods, T.J.; Subedi, K.; Harley, B.A.C. The inclusion of zinc into mineralized collagen scaffolds for craniofacial bone repair applications. Acta Biomater. 2019, 93, 86–96. [Google Scholar] [CrossRef]
- De Aragão Tavares, E.; De Medeiros, W.M.T.Q.; De Assis Pontes, T.P.; Barbosa, M.M.; De Araújo, A.A.; De Araújo, R.F.; Figueiredo, J.G.; Leitão, R.C.; Da Silva Martins, C.; Da Silva, F.O.N.; et al. Chitosan membrane modified with a new zinc(II)-vanillin complex improves skin wound healing in diabetic rats. Front. Pharmacol. 2019, 9, 1511. [Google Scholar] [CrossRef]
- Priyadarshi, R.; Kumar, B.; Rhim, J.W. Green and facile synthesis of carboxymethylcellulose/ZnO nanocomposite hydrogels crosslinked with Zn2+ ions. Int. J. Biol. Macromol. 2020, 162, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Ciofani, G.; Genchi, G.G.; Mattoli, V. ZnO nanowire arrays as substrates for cell proliferation and differentiation. Mater. Sci. Eng. C 2012, 32, 341–347. [Google Scholar] [CrossRef]
- Lee, J.; Kang, B.S.; Hicks, B.; Chancellor, T.F.; Chu, B.H.; Wang, H.T.; Keselowsky, B.G.; Ren, F.; Lele, T.P. The control of cell adhesion and viability by zinc oxide nanorods. Biomaterials 2008, 29, 3743–3749. [Google Scholar] [CrossRef]
- Jiménez, M.; Abradelo, C.; San Román, J.; Rojo, L. Bibliographic review on the state of the art of strontium and zinc based regenerative therapies. Recent developments and clinical applications. J. Mater. Chem. B 2019, 7, 1974–1985. [Google Scholar] [CrossRef]
- Bae, Y.S.; Hill, N.D.; Bibi, Y.; Dreiher, J.; Cohen, A.D. Innovative uses for zinc in dermatology. Dermatol. Clin. 2010, 28, 587–597. [Google Scholar] [CrossRef]
- Azgın, İ.; Arbağ, H.; Eryılmaz, M.A.; Çelik, Z.E. The effects of local and intraperitoneal zinc treatments on maxillofacial fracture healing in rabbits. J. Cranio-Maxillofac. Surg. 2020, 48, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Sharir, H.; Zinger, A.; Nevo, A.; Sekler, I.; Hershfinkel, M. Zinc released from injured cells is acting via the Zn2+-sensing receptor, ZnR, to trigger signaling leading to epithelial repair. J. Biol. Chem. 2010, 285, 26097–26106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.; Varigos, G.; Ackland, M.L. Apoptosis may underlie the pathology of zinc-deficient skin. Immunol. Cell Biol. 2006, 84, 28–37. [Google Scholar] [CrossRef]
- Michaelsson, G.; Ljunghall, K.; Danielson, B.G. Zinc in epidermis and dermis in healthy subjects. Acta Derm. Venereol. 1980, 60, 295–299. [Google Scholar]
- Pastar, I.; Stojadinovic, O.; Tomic-Canic, M. Role of keratinocytes in healing of chronic wounds. Surg. Technol. Int. 2008, 17, 105–112. [Google Scholar] [PubMed]
- Lansdown, A.B.G.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Ågren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef]
- Holmes, A.M.; Mackenzie, L.; Roberts, M.S. Disposition and measured toxicity of zinc oxide nanoparticles and zinc ions against keratinocytes in cell culture and viable human epidermis. Nanotoxicology 2020, 14, 263–274. [Google Scholar] [CrossRef]
- Li, Y.; Maret, W. Transient fluctuations of intracellular zinc ions in cell proliferation. Exp. Cell Res. 2009, 315, 2463–2470. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Tsuruoka, S.; Ioka, T.; Ando, H.; Ito, C.; Akimoto, T.; Fujimura, A.; Asano, Y.; Kusano, E. Indoxyl sulfate stimulates proliferation of rat vascular smooth muscle cells. Kidney Int. 2006, 69, 1780–1785. [Google Scholar] [CrossRef] [Green Version]
- Bozym, R.A.; Chimienti, F.; Giblin, L.J.; Gross, G.W.; Korichneva, I.; Li, Y.; Libert, S.; Maret, W.; Parviz, M.; Frederickson, C.J.; et al. Free zinc ions outside a narrow concentration range are toxic to a variety of cells in vitro. Exp. Biol. Med. Maywood 2010, 235, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Guo, D.; Wu, Q.; Liu, D.; Bi, H. Zinc chloride inhibits human lens epithelial cell migration and proliferation involved in TGF-β1 and TNF-α signaling pathways in HLE B-3 cells. Biol. Trace Elem. Res. 2014, 159, 425–433. [Google Scholar] [CrossRef]
- Salazar-García, S.; García-Rodrigo, J.F.; Martínez-Castañón, G.A.; Ruiz-Rodríguez, V.M.; Portales-Pérez, D.P.; Gonzalez, C. Silver nanoparticles (AgNPs) and zinc chloride (ZnCl2) exposure order determines the toxicity in C6 rat glioma cells. J. Nanopart. Res. 2020, 22, 253. [Google Scholar] [CrossRef]
- Tekuri, S.K.; Bassaiahgari, P.; Gali, Y.; Amuru, S.R.; Pabbaraju, N. Determination of Median Lethal Dose of Zinc chloride in Wistar Rat. Adv. Anim. Vet. Sci. 2021, 9, 393–399. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Guo, Y.; Lu, J.; Veeraraghavan, V.P.; Mohan, S.K.; Wang, C.; Yu, X. Synthesis of Zinc oxide nanoparticles from Marsdenia tenacissima inhibits the cell proliferation and induces apoptosis in laryngeal cancer cells (Hep-2). J. Photochem. Photobiol. B Biol. 2019, 201, 111624. [Google Scholar] [CrossRef] [PubMed]
- Emri, E.; Miko, E.; Bai, P.; Boros, G.; Nagy, G.; Rózsa, D.; Juhász, T.; Hegeds, C.; Horkay, I.; Remenyik, É.; et al. Effects of non-toxic zinc exposure on human epidermal keratinocytes. Metallomics 2015, 7, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamore, S.D.; Cabello, C.M.; Wondrak, G.T. The topical antimicrobial zinc pyrithione is a heat shock response inducer that causes DNA damage and PARP-dependent energy crisis in human skin cells. Cell Stress Chaperones 2010, 15, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Pie, J.E.; Kim, Y.R.; Lee, H.R.; Son, S.W.; Kim, M.K. Effects of zinc oxide nanoparticles on gene expression profile in human keratinocytes. Mol. Cell. Toxicol. 2012, 8, 113–118. [Google Scholar] [CrossRef]
- Sato, R.; Yamamoto, H.; Kasai, K.; Yamauchi, M. Distribution pattern of versican, link protein and hyaluronic acid in the rat periodontal ligament during experimental tooth movement. J. Periodontal Res. 2002, 37, 15–22. [Google Scholar] [CrossRef]
- Zimmermann, D.R.; Dours-Zimmermann, M.T.; Schubert, M.; Bruckner-Tuderman, L. Versican is expressed in the proliferating zone in the epidermis and in association with the elastic network of the dermis. J. Cell Biol. 1994, 124, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Beikler, T.; Peters, U.; Prior, K.; Eisenacher, M.; Flemmig, T.F. Gene expression in periodontal tissues following treatment. BMC Med. Genom. 2008, 1, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.J.; Harris, T.J.C. Cadherin Trafficking for Tissue Morphogenesis: Control and Consequences. Traffic 2016, 17, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.F.; Chiang, H.S.; Lo, H.M.; Jian, J.S.; Wu, W. Bin E-cadherin and its downstream catenins are proteolytically cleaved in human HaCaT keratinocytes exposed to UVB. Exp. Dermatol. 2006, 15, 315–321. [Google Scholar] [CrossRef]
- Yeh, J.T.; Yeh, L.K.; Jung, S.M.; Chang, T.J.; Wu, H.H.; Shiu, T.F.; Liu, C.Y.; Kao, W.W.Y.; Chu, P.H. Impaired skin wound healing in lumican-null mice. Br. J. Dermatol. 2010, 163, 1174–1180. [Google Scholar] [CrossRef]
- Niewiarowska, J.; Brézillon, S.; Sacewicz-Hofman, I.; Bednarek, R.; Maquart, F.X.; Malinowski, M.; Wiktorska, M.; Wegrowski, Y.; Cierniewski, C.S. Lumican inhibits angiogenesis by interfering with α2β1 receptor activity and downregulating MMP-14 expression. Thromb. Res. 2011, 128, 452–457. [Google Scholar] [CrossRef]
- Liu, X.J.; Kong, F.Z.; Wang, Y.H.; Zheng, J.H.; Wan, W.D.; Deng, C.L.; Mao, G.Y.; Li, J.; Yang, X.M.; Zhang, Y.L.; et al. Lumican Accelerates Wound Healing by Enhancing α2β1 Integrin-Mediated Fibroblast Contractility. PLoS ONE 2013, 8, e67124. [Google Scholar] [CrossRef] [PubMed]
- Handford, P.A. Fibrillin-1, a calcium binding protein of extracellular matrix. Biochim. Biophys. Acta BBA Mol. Cell Res. 2000, 1498, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Adamo, C.S.; Zuk, A.V.; Sengle, G. The fibrillin microfibril/elastic fibre network: A critical extracellular supramolecular scaffold to balance skin homoeostasis. Exp. Dermatol. 2021, 30, 25–37. [Google Scholar] [CrossRef]
- Philips, N.; Samuel, M.; Arena, R.; Chen, Y.J.; Conte, J.; Natrajan, P.; Haas, G.; Gonzalez, S. Direct inhibition of elastase and matrixmetalloproteinases and stimulation of biosynthesis of fibrillar collagens, elastin, and fibrillins by xanthohumol. J. Cosmet. Sci. 2010, 61, 125–132. [Google Scholar] [CrossRef]
- Dzamba, B.J.; Keene, D.R.; Isogai, Z.; Charbonneau, N.L.; Karaman-Jurukovska, N.; Simon, M.; Sakai, L.Y. Assembly of epithelial cell fibrillins. J. Investig. Dermatol. 2001, 117, 1612–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Kruk, J.; Duchnik, E. Oxidative stress and skin diseases: Possible role of physical activity. Asian Pac. J. Cancer Prev. 2014, 15, 561–568. [Google Scholar] [CrossRef]
- Butt, H.; Mehmood, A.; Ali, M.; Tasneem, S.; Tarar, M.N.; Riazuddin, S. Vitamin E preconditioning alleviates in vitro thermal stress in cultured human epidermal keratinocytes. Life Sci. 2019, 239, 116972. [Google Scholar] [CrossRef]
- Calcabrini, C.; De Bellis, R.; Mancini, U.; Cucchiarini, L.; Potenza, L.; De Sanctis, R.; Patrone, V.; Scesa, C.; Dachà, M. Rhodiola rosea ability to enrich cellular antioxidant defences of cultured human keratinocytes. Arch. Dermatol. Res. 2010, 302, 191–200. [Google Scholar] [CrossRef]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; de la Vega, L.; Muñoz, E. Cannabidiol induces antioxidant pathways in keratinocytes by targeting BACH1. Redox Biol. 2020, 28, 101321. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Buontempo, F.; Martelli, A.M.; Falcieri, E.; Battistelli, M. Protective effect of different antioxidant agents in UVB-irradiated keratinocytes. Eur. J. Histochem. 2017, 61, 215–221. [Google Scholar] [CrossRef]
- Son, D.H.; Yang, D.J.; Sun, J.S.; Kim, S.K.; Kang, N.; Kang, J.Y.; Choi, Y.H.; Lee, J.H.; Moh, S.H.; Shin, D.M.; et al. A novel peptide, nicotinyl–isoleucine–valine–histidine (Na–IVH), promotes antioxidant gene expression and wound healing in HaCaT cells. Mar. Drugs 2018, 16, 262. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Venkataraman, S.; Coleman, M.C.; Spitz, D.R.; Wertz, P.W.; Domann, F.E. Glutathione peroxidase-1 inhibits UVA-induced AP-2α expression in human keratinocytes. Biochem. Biophys. Res. Commun. 2006, 351, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, H.R.; Mazurier, F.; Cario-André, M.; Pain, C.; Ged, C.; Taïeb, A.; De Verneuil, H. Protective effects of catalase overexpression on UVB-induced apoptosis in normal human keratinocytes. J. Biol. Chem. 2006, 281, 17999–18007. [Google Scholar] [CrossRef] [Green Version]
- Miyachi, Y. Photoaging from an oxidative standpoint. J. Dermatol. Sci. 1995, 9, 79–86. [Google Scholar] [CrossRef]
- Dahle, J.; Kvam, E.; Stokke, T. Bystander effects in UV-induced genomic instability: Antioxidants inhibit delayed mutagenesis induced by ultraviolet A and B radiation. J. Carcinog. 2005, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H.; Izutsu, Y.; Yahagi, S.; Okano, Y. Reactive oxygen species in HaCaT keratinocytes after UVB irradiation are triggered by intracellular Ca2+ levels. J. Investig. Dermatol. Symp. Proc. 2009, 14, 50–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluemsamran, T.; Onkoksoong, T.; Panich, U. Caffeic acid and ferulic acid inhibit UVA-induced matrix metalloproteinase-1 through regulation of antioxidant defense system in keratinocyte HaCaT cells. In Proceedings of the Photochemistry and Photobiology. Photochem. Photobiol. 2012, 88, 961–968. [Google Scholar] [CrossRef]
- Wu, F.; Cui, L. Resveratrol suppresses melanoma by inhibiting NF-κB/miR-221 and inducing TFG expression. Arch. Dermatol. Res. 2017, 309, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K. Preservation effects of geniposidic acid on human keratinocytes (HaCaT) against UVB. Biomed. Dermatol. 2018, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.H.; Ku, J.-E.; Rhie, S.-J.; Ryu, J.Y.; Bae, S.; Kim, Y.-S. Anti-oxidant and Anti-inflammatory Effects of Sinapic Acid in UVB Irradiation-Damaged HaCaT Keratinocytes. Asian J. Beauty Cosmetol. 2017, 15, 513–522. [Google Scholar] [CrossRef]
- Salesa, B.; Serrano-Aroca, Á. Multi-Layer Graphene Oxide in Human Keratinocytes: Time-Dependent Cytotoxicity, Proliferation, and Gene Expression. Coatings 2021, 11, 414. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol (Access Number) | Gene Name | Oligo Sequences | Function |
|---|---|---|---|
| ACTB (NM_001101) | Actin beta | 5′-CCATGCCCACCATCACGC-3′ | Highly conserved protein that are involved in cell motility, structure and integrity. |
| 5′-CACAGAGCCTCGCCTTTG-3′ | |||
| CAT (NM_001752) | Catalase | 5′-TGAATGAGGAACAGAGGAAACG-3′ | Encodes a key antioxidant enzyme (catalase) in the body defense against oxidative stress. |
| 5′-AGATCCGGACTGCACAAAG-3′ | |||
| MMP1 (NM_001145938) | Matrix metallopeptidase 1 | 5′-GGACCATGCCATTGAGAAAG-3′ | Involved in the breakdown of extracellular matrix in normal physiological processes. |
| 5′-TCCTCCAGGTCCATCAAAAG-3′ | |||
| GPX1 (NM_000581) | Glutathione peroxidase 1 | 5′-TTTGGGCATCAGGAGAACGC-3′ | Catalyze the reduction of organic hydroperoxides and hydrogen peroxide by glutathione, and thereby protect cells against oxidative damage. |
| 5′-ACCGTTCACCTCGCACTTC-3′ | |||
| COL4A1 (NM_000088) | Collagen type I alpha 1 | 5′-CAAGGGCGACAGAGGTTTGC-3′ | Abundant in bone, cornea, dermis and tendon. Mutations in this gene are associated with osteogenesis imperfect types I–IV. |
| 5′-AAAACTCACCAGGCTCCCCC-3′ | |||
| TGFB1 (NM_000660) | Transforming growth factor beta 1 | 5′-AGCTGTACATTGACTTCCGCA-3′ | Regulates cell proliferation, differentiation and growth. |
| 5′-TGTCCAGGCTCCAAATGTAGG-3′ | |||
| HAS2 (NM_005328) | Hyaluronan synthase 2 | 5′-CCGAGAATGGCTGTACAATGC-3′ | Involved in a variety of functions, such as space filling, lubrication of joints and provision of a matrix through which cells can migrate. |
| 5′-AGAGCTGGATTACTGTGGCAA-3′ | |||
| LAMB1 (NM_002291) | Laminin subunit beta 1 | 5′-CAGGGTGTGCAGTCAGGGAA-3′ | Implicated in a wide variety of biological processes, including cell adhesion, differentiation, migration, signaling, neurite outgrowth and metastasis. |
| 5′-TGTGTCTGCGTTGAGGGTGT-3′ | |||
| LUM (NM_002345) | Lumican | 5′-ACTTGGGTAGCTTTCAGGGCA-3′ | Is the major keratan sulfate proteoglycan of the cornea, but is also distributed in interstitial collagenous matrices throughout the body. |
| 5′-TTCCTGGCATTGATTGGTGGT-3′ | |||
| FN1 (NM_001306129) | Fibronectin 1 | 5′-GGCCAGTCCTACAACCAGT-3′ | Involved in cell adhesion and migration processes, including embryogenesis, wound healing, blood coagulation, host defense and metastasis. |
| 5′-CGGGAATCTTCTCTGTCAGC-3′ | |||
| VCAN (NM_001126336) | Versican | 5′-CTGGTCTCCGCTGTATCCTG-3′ | Involved in cell adhesion, proliferation, migration and angiogenesis and plays a central role in tissue morphogenesis and maintenance. |
| 5′-ATCGCTGCAAAATGAACCCG-3′ | |||
| CDH1 (NM_001317184) | Cadherin 1 | 5′-AACAGCACGTACACAGCCCT-3′ | Loss of function of this gene is thought to contribute to cancer progression by increasing proliferation, invasion and/or metastasis. |
| 5′-TCTGGTATGGGGGCGTTGTC-3′ | |||
| FBN (NM_000138) | Fibrillin 1 | 5′-ATCCAACCACGTGCATCAGT-3′ | Extracellular matrix glycoprotein that serves as a structural component of calcium-binding microfibrils, providing force-bearing structural support in elastic and non-elastic connective tissue throughout the body. |
| 5′-AGAGCGGGTATCAACACAGC-3′ | |||
| SOD1 (NM_000454) | Superoxide dismutase 1 | 5′-GGTGTGGCCGATGTGTCT-3′ | The protein encoded by this gene binds copper and Zn2+ ions and is one of two isozymes responsible for destroying free superoxide radicals in the body. |
| 5′-TCCACCTTTGCCCAAGTCA-3′ |
| ZnCl2 Exposure | EC50 (µg/mL) | 95% CI | R Square |
|---|---|---|---|
| 3 h | 193.1 | 172.6–216.5 | 0.9524 |
| 12 h | 16.8 | 16.5–17.1 | 0.9598 |
| 24 h | 13.5 | 13.2–13.6 | 0.9799 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salesa, B.; Sabater i Serra, R.; Serrano-Aroca, Á. Zinc Chloride: Time-Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes. Biology 2021, 10, 1072. https://doi.org/10.3390/biology10111072
Salesa B, Sabater i Serra R, Serrano-Aroca Á. Zinc Chloride: Time-Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes. Biology. 2021; 10(11):1072. https://doi.org/10.3390/biology10111072
Chicago/Turabian StyleSalesa, Beatriz, Roser Sabater i Serra, and Ángel Serrano-Aroca. 2021. "Zinc Chloride: Time-Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes" Biology 10, no. 11: 1072. https://doi.org/10.3390/biology10111072
APA StyleSalesa, B., Sabater i Serra, R., & Serrano-Aroca, Á. (2021). Zinc Chloride: Time-Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes. Biology, 10(11), 1072. https://doi.org/10.3390/biology10111072