
Sheep β-Defensin 2 Regulates Escherichia coli F17 Resistance via NF-κB and MAPK Signaling Pathways in Ovine Intestinal Epithelial Cells


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sample and Ethical Statement
2.2. Cell Culture and E. coli F17 Stimulation in OIECs
2.3. Plasmid Construction and RNA Oligonucleotides
2.4. Cell Transfection
2.5. Pathway Inhibitors Blocking Signaling Pathway Test
2.6. Total RNA Extraction and Real-Time PCR (RT-PCR)
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Plate Counting Method for Bacteria Enumeration
2.9. Statistical Analysis
3. Results
3.1. Effect of E. coli F17 Different Infection Concentration and Time on SBD-2 Expression
3.2. Expression Level of SBD-2 Regulates E. coli F17 Resistance in OIECs
3.3. Effect of E. coli F17 Stimulation in OIECs on NF-κB and MAPK Pathways
3.4. NF-κB and MAPK Pathways Influence SBD-2 Expression at the mRNA and Protein Levels
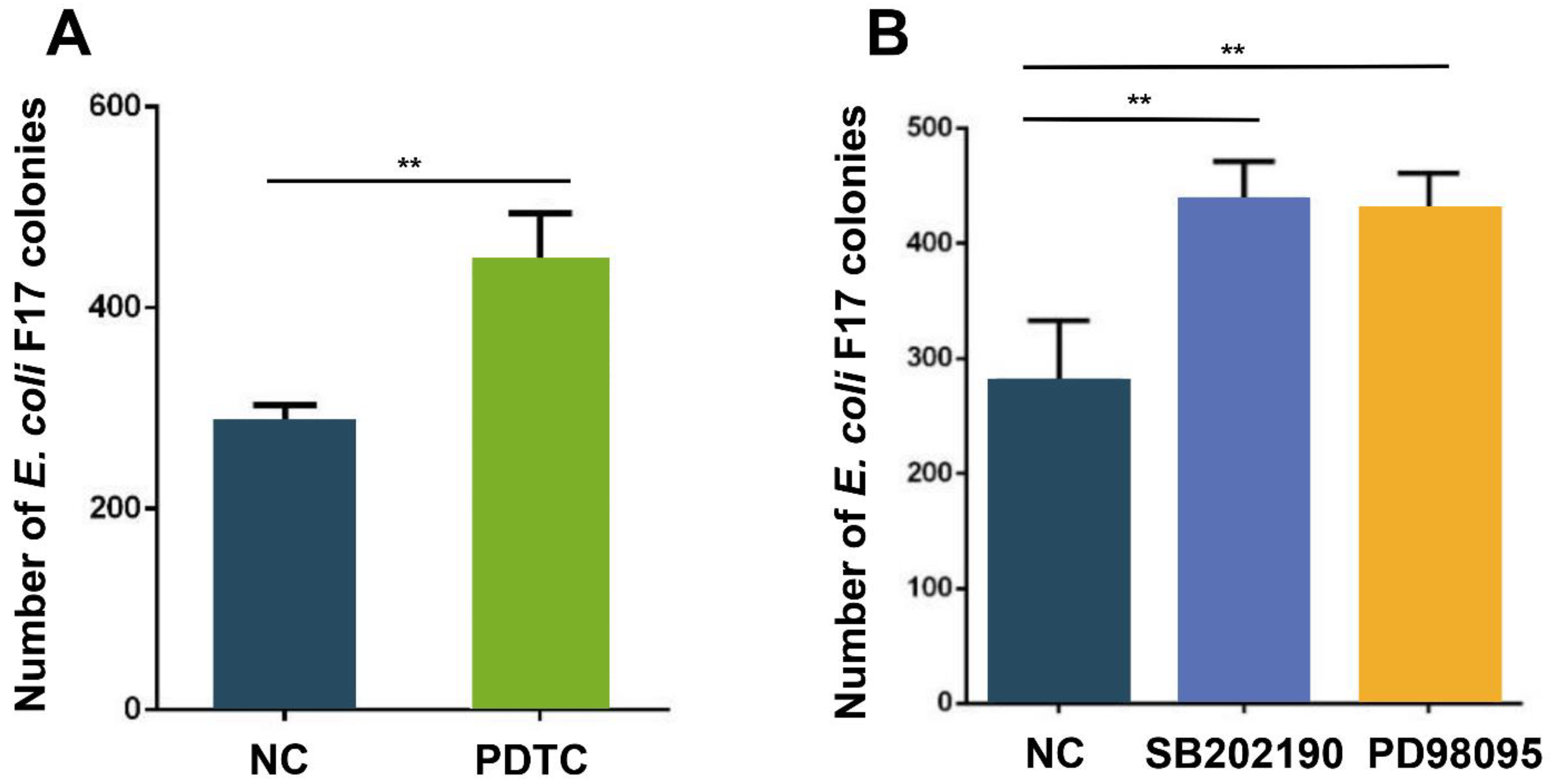
3.5. NF-κB and MAPK Pathways Regulate Adhesion of E. coli F17 to OIECs
4. Discussion
4.1. Effect of SBD-2 Expression on E. coli F17 Adhesion to OIECs
4.2. Effect of NF-κB and MAPK Pathways on SBD-2 Expression and E. coli F17 Adhesion to OIECs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagy, B.; Fekete, P.Z. Enterotoxigenic Escherichia coli in veterinary medicine. Int. J. Med. Microbiol. 2005, 295, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Shabana, I.I.; Bouqellah, N.A.; Zaraket, H. Investigation of viral and bacterial enteropathogens of diarrheic sheep and goats in Medina, Saudi Arabia. Trop. Biomed. 2017, 34, 944–955. [Google Scholar] [PubMed]
- Kopic, S.; Geibel, J.P. Toxin mediated diarrhea in the 21 century: The pathophysiology of intestinal ion transport in the course of ETEC, V. cholerae and rotavirus infection. Toxins 2010, 2, 2132–2157. [Google Scholar] [CrossRef] [Green Version]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Lintermans, P.F.; Pohl, P.; Bertels, A.; Charlier, G.; Vandekerckhove, J.; Van Damme, J.; Schoup, J.; Schlicker, C.; Korhonen, T.; De Greve, H.; et al. Characterization and purification of the F17 adhesin on the surface of bovine enteropathogenic and septicemic Escherichia coli. Am. J. Vet. Res. 1988, 49, 1794–1799. [Google Scholar] [PubMed]
- Holly, M.K.; Diaz, K.; Smith, J.G. Defensins in Viral Infection and Pathogenesis. Annu. Rev. Virol. 2017, 4, 369–391. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A.; Ackermann, M.; McCray, P.B., Jr.; Tack, B.F. Antimicrobial peptides in animals and their role in host defences. Int. J. Antimicrob. Agents 2003, 22, 465–478. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Lehrer, R.I. Antimicrobial peptides of vertebrates. Curr. Opin. Immunol. 1998, 10, 41–44. [Google Scholar] [CrossRef]
- Huttner, K.M.; Lambeth, M.R.; Burkin, H.R.; Burkin, D.J.; Broad, T.E. Localization and genomic organization of sheep antimicrobial peptide genes. Gene 1998, 206, 85–91. [Google Scholar] [CrossRef]
- Hou, J.; Liu, H.Y.; Diao, H.; Yu, H. The truncated human beta-defensin 118 can modulate lipopolysaccharide mediated inflammatory response in RAW264.7 macrophages. Peptides 2021, 136, 170438. [Google Scholar] [CrossRef]
- Ostuni, R.; Zanoni, I.; Granucci, F. Deciphering the complexity of Toll-like receptor signaling. Cell. Mol. Life Sci. CMLS 2010, 67, 4109–4134. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bian, T.; Lyu, J.; Cui, D.; Lei, L.; Yan, F. Human β-defensin-3 alleviates the progression of atherosclerosis accelerated by Porphyromonas gingivalis lipopolysaccharide. Int. Immunopharmacol. 2016, 38, 204–213. [Google Scholar] [CrossRef]
- Han, F.; Zhang, H.; Xia, X.; Xiong, H.; Song, D.; Zong, X.; Wang, Y. Porcine β-defensin 2 attenuates inflammation and mucosal lesions in dextran sodium sulfate-induced colitis. J. Immunol. 2015, 194, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jouve, M.; Garcia, M.I.; Courcoux, P.; Labigne, A.; Gounon, P.; Le Bouguénec, C. Adhesion to and invasion of HeLa cells by pathogenic Escherichia coli carrying the afa-3 gene cluster are mediated by the AfaE and AfaD proteins, respectively. Infect. Immun. 1997, 65, 4082–4089. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef] [Green Version]
- Souza, B.B.; Barbosa, E.M.; Azevedo, J.S.; Campelo, J.E.; Rodrigues, L.F.; Pinheiro, L.M.; Silva, S.C.; Schierholt, A.S.; Souza, P.H.; Gonçalves, E.C.; et al. Genetic polymorphisms in β-defensin II gene in Amazon sheep from Brazil. Genet. Mol. Res. GMR 2015, 14, 12805–12810. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, M.; Zhu, X.M.; Fan, Y.R.; Du, C.G.; Bao, H.E.; Xu, S.G.; Tian, Q.Z.; Wang, Y.H.; Yang, Y.F. Modulation of ovine SBD-1 expression by Saccharomyces cerevisiae in ovine ruminal epithelial cells. BMC Vet. Res. 2018, 14, 134. [Google Scholar] [CrossRef]
- Zhang, M.; Jin, X.; Yang, Y.F. β-Glucan from Saccharomyces cerevisiae induces SBD-1 production in ovine ruminal epithelial cells via the Dectin-1-Syk-NF-κB signaling pathway. Cell. Signal. 2019, 53, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Zhang, M.; Yang, Y.F. Saccharomyces cerevisiae β-glucan-induced SBD-1 expression in ovine ruminal epithelial cells is mediated through the TLR-2-MyD88-NF-κB/MAPK pathway. Vet. Res. Commun. 2019, 43, 77–89. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, M.; Cao, G.F.; Yang, Y.F. Saccharomyces cerevisiae mannan induces sheep beta-defensin-1 expression via Dectin-2-Syk-p38 pathways in ovine ruminal epithelial cells. Vet. Res. 2019, 50, 8. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Tian, Q.Z.; Zhang, M.; Cao, G.F.; Yang, Y.F. Modulation of SBD-1 expression by Saccharomyces cerevisiae cell wall components in ovine ruminal epithelial cells. Res. Vet. Sci. 2020, 129, 28–38. [Google Scholar] [CrossRef]
- Huttner, K.M.; Brezinski-Caliguri, D.J.; Mahoney, M.M.; Diamond, G. Antimicrobial peptide expression is developmentally regulated in the ovine gastrointestinal tract. J. Nutr. 1998, 128, 297s–299s. [Google Scholar] [CrossRef] [Green Version]
- Meyerholz, D.K.; Gallup, J.M.; Grubor, B.M.; Evans, R.B.; Tack, B.F.; McCray, P.B., Jr.; Ackermann, M.R. Developmental expression and distribution of sheep beta-defensin-2. Dev. Comp. Immunol. 2004, 28, 171–178. [Google Scholar] [CrossRef]
- Shi, N.; Jin, F.; Zhang, X.; Clinton, S.K.; Pan, Z.; Chen, T. Overexpression of human β-defensin 2 promotes growth and invasion during esophageal carcinogenesis. Oncotarget 2014, 5, 11333–11344. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, C.; Di, T.; Yang, J.; Wang, K.; Zhu, Y.; Zhu, R.; Zhou, A.; Qian, Y. Human β-Defensin-2 Improves Hyperoxia-Induced Lung Structural and Functional Injury in Neonatal Rats. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 6074–6084. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cheng, Y.T.; Tan, M.F.; Zhang, H.W.; Liu, W.Q.; Zou, G.; Zhang, L.S.; Zhang, C.Y.; Deng, S.M.; Yu, L.; et al. Overexpression of Porcine Beta-Defensin 2 Enhances Resistance to Actinobacillus pleuropneumoniae Infection in Pigs. Infect. Immun. 2015, 83, 2836–2843. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yang, X.; Wang, A.; Huang, C.; Tang, H.; Zhang, Q.; Fang, Q.; Yu, Z.; Liu, X.; Huang, Q.; et al. Pigs Overexpressing Porcine β-Defensin 2 Display Increased Resilience to Glaesserella parasuis Infection. Antibiotics 2020, 9, 903. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Nagase, T.; Tomita, T.; Nakamura, K.; Fukuhara, S.; Amano, T.; Yamamoto, H.; Ide, Y.; Suzuki, M.; Teramoto, S.; et al. Beta-defensin overexpression induces progressive muscle degeneration in mice. Am. J. Physiol. Cell Physiol. 2007, 292, C2141–C2149. [Google Scholar] [CrossRef] [PubMed]
- Röhrl, J.; Huber, B.; Koehl, G.E.; Geissler, E.K.; Hehlgans, T. Mouse β-defensin 14 (Defb14) promotes tumor growth by inducing angiogenesis in a CCR6-dependent manner. J. Immunol. 2012, 188, 4931–4939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziv, C.; Yarden, O. Gene silencing for functional analysis: Assessing RNAi as a tool for manipulation of gene expression. Methods Mol. Biol. 2010, 638, 77–100. [Google Scholar] [CrossRef]
- Sang, Y.; Ramanathan, B.; Ross, C.R.; Blecha, F. Gene silencing and overexpression of porcine peptidoglycan recognition protein long isoforms: Involvement in beta-defensin-1 expression. Infect. Immun. 2005, 73, 7133–7141. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, D.A.; Cole, S.P.; Martin-Porter, E.; Housley, M.P.; Liu, L.; Ganz, T.; Kagnoff, M.F. Regulation of human beta-defensins by gastric epithelial cells in response to infection with Helicobacter pylori or stimulation with interleukin-1. Infect. Immun. 2000, 68, 5412–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Xie, K.; Chen, D.; Yu, B.; Mao, X.; Yu, J.; Luo, J.; Zheng, P.; Luo, Y.; Yan, H.; et al. Expression and Functional Characterization of a Novel Antimicrobial Peptide: Human Beta-Defensin 118. BioMed Res. Int. 2020, 2020, 1395304. [Google Scholar] [CrossRef]
- Su, G.; Xie, K.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; et al. Differential expression, molecular cloning, and characterization of porcine beta defensin 114. J. Anim. Sci. Biotechnol. 2019, 10, 60. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, H.; Karino, M.; Tobe, T. Modulation of the Inflammasome Signaling Pathway by Enteropathogenic and Enterohemorrhagic Escherichia coli. Front. Cell. Infect. Microbiol. 2016, 6, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.; Han, J.; Ma, Q.; Cheng, B.; Shan, A.; Gao, N.; Yang, Y. TLR2/4-mediated NF-κB pathway combined with the histone modification regulates β-defensins and interleukins expression by sodium phenyl butyrate in porcine intestinal epithelial cells. Food Nutr. Res. 2018, 62, 1493. [Google Scholar] [CrossRef]
- Lan, J.; Dou, X.; Li, J.; Yang, Y.; Xue, C.; Wang, C.; Gao, N.; Shan, A. l-Arginine Ameliorates Lipopolysaccharide-Induced Intestinal Inflammation through Inhibiting the TLR4/NF-κB and MAPK Pathways and Stimulating β-Defensin Expression in Vivo and in Vitro. J. Agric. Food Chem. 2020, 68, 2648–2663. [Google Scholar] [CrossRef]
- Liu, H.; Yu, H.; Gu, Y.; Xin, A.; Zhang, Y.; Diao, H.; Lin, D. Human beta-defensin DEFB126 is capable of inhibiting LPS-mediated inflammation. Appl. Microbiol. Biotechnol. 2013, 97, 3395–3408. [Google Scholar] [CrossRef]
- Cui, L.; Wang, H.; Lin, J.; Wang, Y.; Dong, J.; Li, J.; Li, J. Progesterone inhibits inflammatory response in E.coli- or LPS-Stimulated bovine endometrial epithelial cells by NF-κB and MAPK pathways. Dev. Comp. Immunol. 2020, 105, 103568. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zheng, Y.; Wang, H.; Dong, J.; Li, J.; Song, Q.; Qian, C.; Li, J. Cortisol inhibits the Escherichia coli-induced endometrial inflammatory response through NF-κB and MAPK pathways in postpartum goats. Anim. Reprod. Sci. 2020, 215, 106333. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cao, Y.; Deng, S.; Jiang, X.; Wang, J.; Zhang, X.; Zhang, J.; Liu, G.; Lian, Z. Overexpression of Toll-like Receptor 4-linked Mitogen-activated Protein Kinase Signaling Contributes to Internalization of Escherichia coli in Sheep. Int. J. Biol. Sci. 2018, 14, 1022–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, C.H.; Choi, Y.S.; Song, S.Y.; Kim, Y.K.; Kim, Y.D. Escherichia coli-derived and Staphylococcus aureus-derived extracellular vesicles induce MUC5AC expression via extracellular signal related kinase 1/2 and p38 mitogen-activated protein kinase in human airway epithelial cells. Int. Forum Allergy Rhinol. 2017, 7, 91–98. [Google Scholar] [CrossRef]
- Li, Q.; Bao, F.; Zhi, D.; Liu, M.; Yan, Q.; Zheng, X.; Ren, L.; Cong, S.; Li, Y.; Cao, G. Lipopolysaccharide induces SBD-1 expression via the P38 MAPK signaling pathway in ovine oviduct epithelial cells. Lipids Health Dis. 2016, 15, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.; Luo, Y.; Chen, D.; Yu, B.; He, J. NF-κB-dependent induction of porcine β-defensin 114 regulates intestinal epithelium homeostasis. Int. J. Biol. Macromol. 2021, 192, 241–249. [Google Scholar] [CrossRef]
- Lin, Q.; Fu, Q.; Li, X.; Luo, Y.; Luo, J.; Chen, D.; Mao, X.; Yu, B.; Zheng, P.; Huang, Z.; et al. Human β-Defensin 118 Attenuates Escherichia coli K88-Induced Inflammation and Intestinal Injury in Mice. Probiotics Antimicrob. Proteins 2021, 13, 586–597. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A Blank Control Group | B Positive Control Group | C Negative Control Group | D Experimental Group |
|---|---|---|---|
| No treatment | E. coli F17 only | NF-κB pathway inhibitor PDTC (25 μM), | NF-κB pathway inhibitor PDTC (25 μM) + E. coli F17, |
| p38 pathway inhibitor SB202190 (25 μM), | p38 pathway inhibitor SB202190 (25 μM) + E. coli F17, | ||
| ERK1/2 pathway inhibitor PD98059 (50 μM), | ERK1/2 pathway inhibitor PD98059 (50 μM) + E. coli F17, | ||
| (Add separately) | (Add separately) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, L.; Zou, S.; Yuan, Z.; Chen, W.; Wang, S.; Cao, X.; Lv, X.; Getachew, T.; Mwacharo, J.M.; Haile, A.; et al. Sheep β-Defensin 2 Regulates Escherichia coli F17 Resistance via NF-κB and MAPK Signaling Pathways in Ovine Intestinal Epithelial Cells. Biology 2021, 10, 1356. https://doi.org/10.3390/biology10121356
Ge L, Zou S, Yuan Z, Chen W, Wang S, Cao X, Lv X, Getachew T, Mwacharo JM, Haile A, et al. Sheep β-Defensin 2 Regulates Escherichia coli F17 Resistance via NF-κB and MAPK Signaling Pathways in Ovine Intestinal Epithelial Cells. Biology. 2021; 10(12):1356. https://doi.org/10.3390/biology10121356
Chicago/Turabian StyleGe, Ling, Shuangxia Zou, Zehu Yuan, Weihao Chen, Shanhe Wang, Xiukai Cao, Xiaoyang Lv, Tesfaye Getachew, Joram M. Mwacharo, Aynalem Haile, and et al. 2021. "Sheep β-Defensin 2 Regulates Escherichia coli F17 Resistance via NF-κB and MAPK Signaling Pathways in Ovine Intestinal Epithelial Cells" Biology 10, no. 12: 1356. https://doi.org/10.3390/biology10121356
APA StyleGe, L., Zou, S., Yuan, Z., Chen, W., Wang, S., Cao, X., Lv, X., Getachew, T., Mwacharo, J. M., Haile, A., & Sun, W. (2021). Sheep β-Defensin 2 Regulates Escherichia coli F17 Resistance via NF-κB and MAPK Signaling Pathways in Ovine Intestinal Epithelial Cells. Biology, 10(12), 1356. https://doi.org/10.3390/biology10121356