
Drinking Molecular Hydrogen Water Is Beneficial to Cardiovascular Function in Diet-Induced Obesity Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
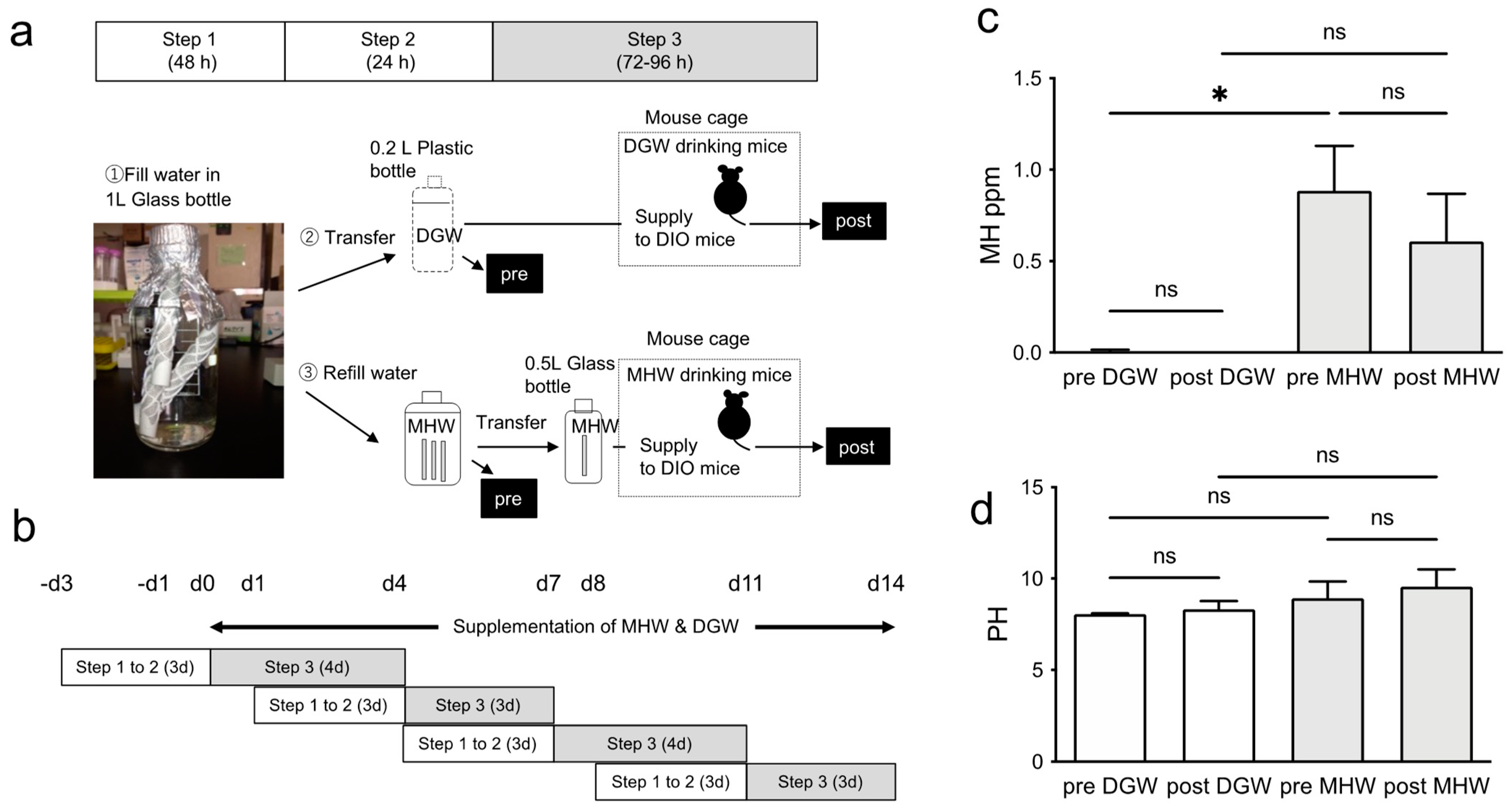
2.1. MHW and DIO Mice
2.2. Hydrogen Concentration and pH of MHW
2.3. Blood Glucose and Body Weight (BW) Measurements
2.4. Measurement of Cardiac Function by Echocardiography
2.5. Sample Preparation for Immunohistochemical Analysis, Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) and Endothelial Progenitor Cell (EPC) Colony Formation Assay
2.6. Immunohistochemical Analysis
2.7. qRT-PCR
2.8. Isolation of Peripheral Blood Mononuclear Cells (PBMCs) and Bone Marrow (BM)-Derived c-Kit + /Sca-1 + Lineage-Negative Cells (BM-KSL Cells)
2.9. EPC Colony Formation Assay
2.10. Statistical Analysis
3. Results
3.1. MH Concentration in Drinking Water
3.2. MHW Had no Effect on BW or Postprandial Blood Sugar (PPBS) of DIO Mice
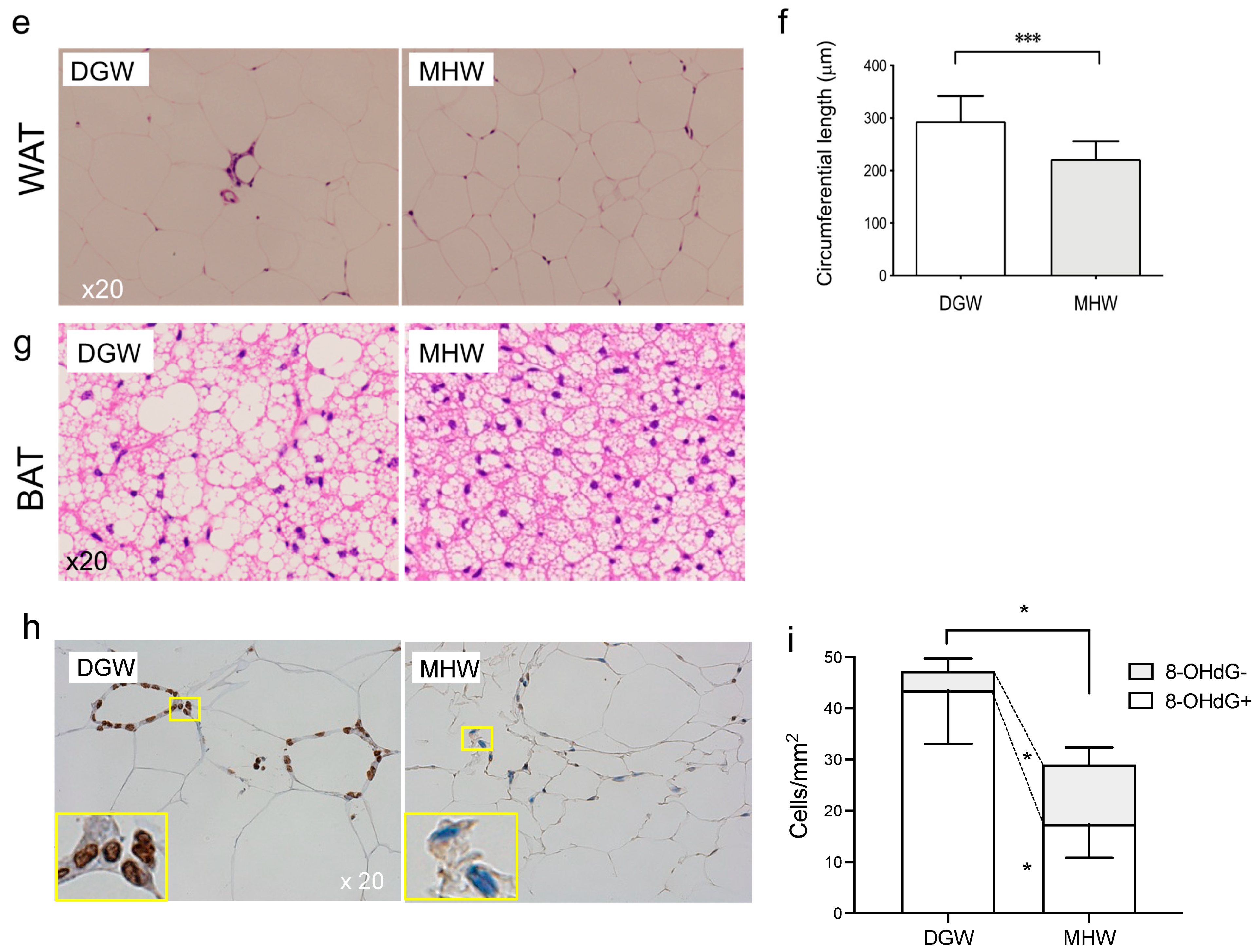
3.3. MHW Morphometrically Changed the Anti-Metabolic Phenotype of Adipose Tissue in DIO Mice
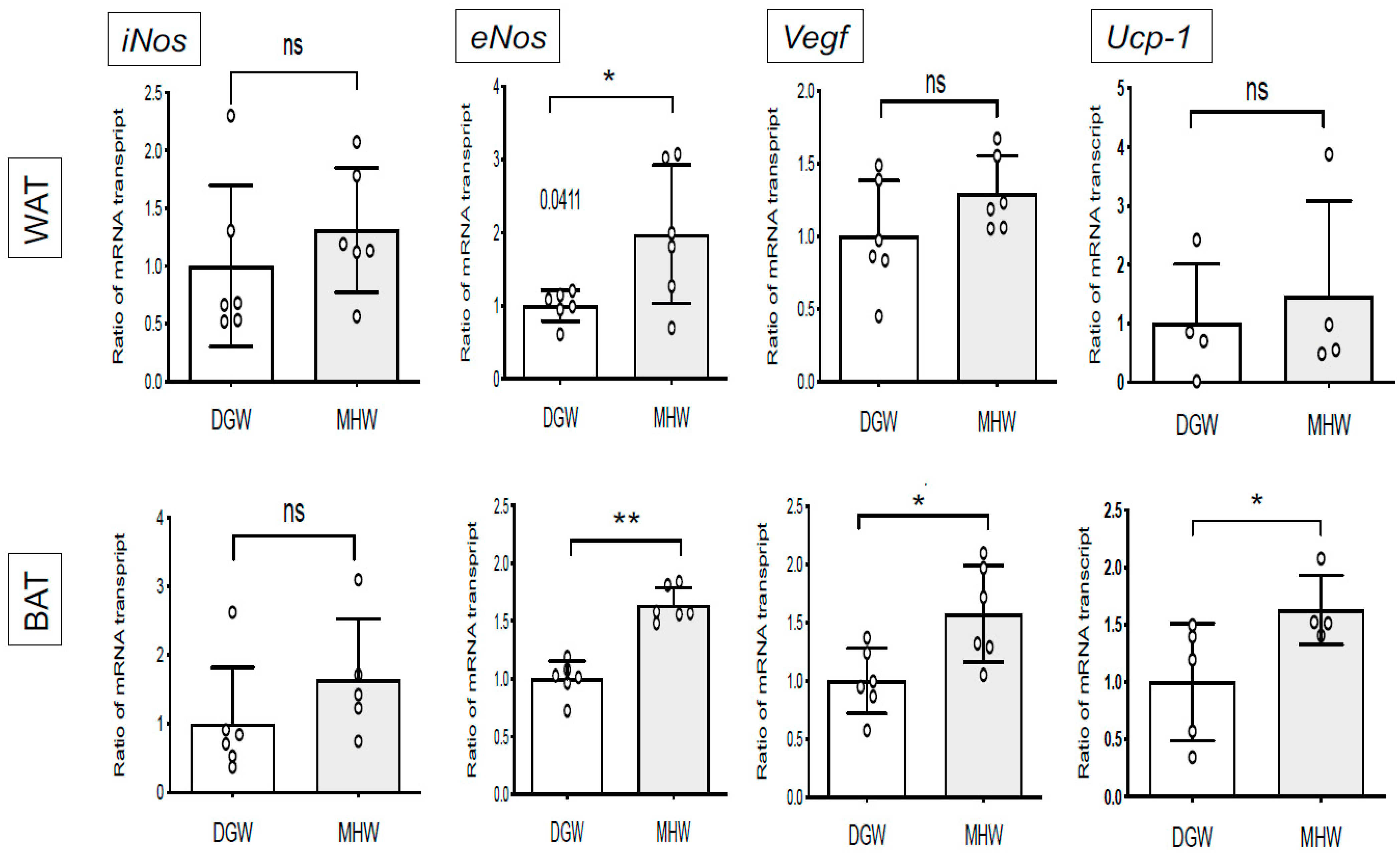
3.4. MHW Intake Upregulated the Expression of Thermogenic- and Angiogenic-Related Genes in the BAT of DIO Mice
3.5. MHW Intake Alleviated Metabolic Cardiomyopathy in DIO Mice
- (1)
- Morphometric improvement in cardiac hypertrophy
- (2)
- MHW intake promoted eNOS phosphorylation in cardiomyocytes of DIO mice
- (3)
- Cardiac arteriolar and capillary dilatation in DIO mice in the MHW group
- (4)
- MHW intake alleviated cardiac oxidative stress in DIO mice in the MHW group
- (5)
- MHW intake improved cardiac function in DIO mice
3.6. EPC Bioactivity Improved in the MHW Group
4. Discussion
4.1. BAT Activation and WAT Browning
4.2. Cardiovascular Protective Effects
4.3. Activation of EPC Bioactivity
4.4. The Unlikely Alkaline Effect of MHW
4.5. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Iuchi, K.; Imoto, A.; Kamimura, N.; Nishimaki, K.; Ichimiya, H.; Yokota, T.; Ohta, S. Molecular hydrogen regulates gene expression by modifying the free radical chain reaction-dependent generation of oxidized phospholipid mediators. Sci. Rep. 2016, 6, 18971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, S. Direct Targets and Subsequent Pathways for Molecular Hydrogen to Exert Multiple Functions: Focusing on Interventions in Radical Reactions. Curr. Pharm. Des. 2021, 27, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, K.; Sano, M.; Ohsawa, I.; Shinmura, K.; Tamaki, K.; Kimura, K.; Endo, J.; Katayama, T.; Kawamura, A.; Kohsaka, S.; et al. Inhalation of hydrogen gas reduces infarct size in the rat model of myocardial ischemia-reperfusion injury. Biochem. Biophys. Res. Commun. 2008, 373, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Qiu, Y.; Ye, G.; Luo, H.; Jiang, J.; Yu, F.; Zhou, W.; Zhang, S.; Feng, J. Treatment with hydrogen molecule attenuates cardiac dysfunction in streptozotocin-induced diabetic mice. Cardiovasc. Pathol. 2015, 24, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, N.; Nishimaki, K.; Ohsawa, I.; Ohta, S. Molecular hydrogen improves obesity and diabetes by inducing hepatic FGF21 and stimulating energy metabolism in db/db mice. Obesity (Silver Spring) 2011, 19, 1396–1403. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Tian, H.; Qin, S.; Sun, X.; Yao, S.; Zong, C.; Luo, Y.; Liu, J.; Yu, Y.; Sang, H.; et al. Hydrogen decreases athero-susceptibility in apolipoprotein B-containing lipoproteins and aorta of apolipoprotein E knockout mice. Atherosclerosis 2012, 221, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Taura, K.; Hatano, E.; Tanabe, K.; Yamamoto, G.; Nakamura, K.; Yamanaka, K.; Kitamura, K.; Narita, M.; Nagata, H.; et al. Effects of oral intake of hydrogen water on liver fibrogenesis in mice. Hepatol. Res. 2014, 44, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.G.; Zhang, B.B.; Wu, Z.Q.; Hang, X.F.; Xu, W.S.; Ni, W.; Zhang, R.Q.; Miao, X.H. Consumption of hydrogen-rich water alleviates renal injury in spontaneous hypertensive rats. Mol. Cell Biochem. 2014, 392, 117–124. [Google Scholar] [CrossRef]
- Asada, R.; Saitoh, Y.; Miwa, N. Effects of hydrogen-rich water bath on visceral fat and skin blotch, with boiling-resistant hydrogen bubbles. Med. Gas. Res. 2019, 9, 68–73. [Google Scholar] [CrossRef]
- LeBaron, T.W.; Singh, R.B.; Fatima, G.; Kartikey, K.; Sharma, J.P.; Ostojic, S.M.; Gvozdjakova, A.; Kura, B.; Noda, M.; Mojto, V.; et al. The Effects of 24-Week, High-Concentration Hydrogen-Rich Water on Body Composition, Blood Lipid Profiles and Inflammation Biomarkers in Men and Women with Metabolic Syndrome: A Randomized Controlled Trial. Diabetes Metab. Syndr. Obes. 2020, 13, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Ji, G.; Pan, R.; Zhao, Y.; Yan, P. Protective effect of hydrogen-rich water on liver function of colorectal cancer patients treated with mFOLFOX6 chemotherapy. Mol. Clin. Oncol. 2017, 7, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.M.; Kang, Y.N.; Choi, I.B.; Gu, Y.; Kawamura, T.; Toyoda, Y.; Nakao, A. Effects of drinking hydrogen-rich water on the quality of life of patients treated with radiotherapy for liver tumors. Med. Gas. Res. 2011, 1, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.M.; Maurovich-Horvat, P.; Liu, C.Y.; Vasan, R.S.; Murabito, J.M.; Meigs, J.B.; Cupples, L.A.; et al. Abdominal visceral and subcutaneous adipose tissue compartments: Association with metabolic risk factors in the Framingham Heart Study. Circulation 2007, 116, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezin, A.E.; Berezin, A.A.; Lichtenauer, M. Emerging Role of Adipocyte Dysfunction in Inducing Heart Failure Among Obese Patients with Prediabetes and Known Diabetes Mellitus. Front. Cardiovasc. Med. 2020, 7, 583175. [Google Scholar] [CrossRef] [PubMed]
- Kajiyama, S.; Hasegawa, G.; Asano, M.; Hosoda, H.; Fukui, M.; Nakamura, N.; Kitawaki, J.; Imai, S.; Nakano, K.; Ohta, M.; et al. Supplementation of hydrogen-rich water improves lipid and glucose metabolism in patients with type 2 diabetes or impaired glucose tolerance. Nutr. Res. 2008, 28, 137–143. [Google Scholar] [CrossRef]
- Nakao, A.; Toyoda, Y.; Sharma, P.; Evans, M.; Guthrie, N. Effectiveness of hydrogen rich water on antioxidant status of subjects with potential metabolic syndrome-an open label pilot study. J. Clin. Biochem. Nutr. 2010, 46, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Li, M.; Sang, H.; Zhang, L.; Li, X.; Yao, S.; Yu, Y.; Zong, C.; Xue, Y.; Qin, S. Hydrogen-rich water decreases serum LDL-cholesterol levels and improves HDL function in patients with potential metabolic syndrome. J. Lipid Res. 2013, 54, 1884–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, R.; Tazawa, K.; Sato, S.; Miwa, N. Effects of hydrogen-rich water prepared by alternating-current-electrolysis on antioxidant activity, DNA oxidative injuries, and diabetes-related markers. Med. Gas. Res. 2020, 10, 114–121. [Google Scholar] [CrossRef]
- Kamimura, N.; Ichimiya, H.; Iuchi, K.; Ohta, S. Molecular hydrogen stimulates the gene expression of transcriptional coactivator PGC-1alpha to enhance fatty acid metabolism. NPJ Aging Mech. Dis. 2016, 2, 16008. [Google Scholar] [CrossRef] [Green Version]
- Reitman, M.L. FGF21: A missing link in the biology of fasting. Cell Metab. 2007, 5, 405–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Invest. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.; Kahn, C.R. Transplantation of adipose tissue and stem cells: Role in metabolism and disease. Nat. Rev. Endocrinol. 2010, 6, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. (Lausanne) 2016, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, T.; Wang, B.; Yang, Q.; de Avila, J.M.; Zhu, M.J.; You, J.; Chen, D.; Du, M. Raspberry promotes brown and beige adipocyte development in mice fed high-fat diet through activation of AMP-activated protein kinase (AMPK) alpha1. J. Nutr. Biochem. 2018, 55, 157–164. [Google Scholar] [CrossRef]
- Gao, S.; Ho, D.; Vatner, D.E.; Vatner, S.F. Echocardiography in Mice. Curr. Protoc. Mouse Biol. 2011, 1, 71–83. [Google Scholar] [CrossRef]
- Tsukada, S.; Kwon, S.M.; Matsuda, T.; Jung, S.Y.; Lee, J.H.; Lee, S.H.; Masuda, H.; Asahara, T. Identification of mouse colony-forming endothelial progenitor cells for postnatal neovascularization: A novel insight highlighted by new mouse colony-forming assay. Stem Cell Res. Ther. 2013, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Cai, L. Diabetes/obesity-related inflammation, cardiac cell death and cardiomyopathy. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2006, 31, 814–818. [Google Scholar]
- Palomer, X.; Salvado, L.; Barroso, E.; Vazquez-Carrera, M. An overview of the crosstalk between inflammatory processes and metabolic dysregulation during diabetic cardiomyopathy. Int. J. Cardiol. 2013, 168, 3160–3172. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Ndisang, J.F. The role of obesity in cardiomyopathy and nephropathy. Curr. Pharm. Des. 2014, 20, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, J.; Wan, Y.; Song, S.; Zhang, Y.; Zhang, G.; Liu, C.; Fu, J. Hydrogen water alleviates lung injury induced by one-lung ventilation. J. Surg. Res. 2015, 199, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Yamafuji, M.; Tachibana, T.; Nakabeppu, Y.; Noda, M.; Nakaya, H. Oral ‘hydrogen water’ induces neuroprotective ghrelin secretion in mice. Sci. Rep. 2013, 3, 3273. [Google Scholar] [CrossRef] [Green Version]
- Francois, M.; Barde, S.; Legrand, R.; Lucas, N.; Azhar, S.; El Dhaybi, M.; Guerin, C.; Hokfelt, T.; Dechelotte, P.; Coeffier, M.; et al. High-fat diet increases ghrelin-expressing cells in stomach, contributing to obesity. Nutrition 2016, 32, 709–715. [Google Scholar] [CrossRef]
- Petrovic, V.; Buzadzic, B.; Korac, A.; Vasilijevic, A.; Jankovic, A.; Korac, B. NO modulates the molecular basis of rat interscapular brown adipose tissue thermogenesis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 147–159. [Google Scholar] [CrossRef]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Picoli, C.C.; Gilio, G.R.; Henriques, F.; Leal, L.G.; Besson, J.C.; Lopes, M.A.; de Moraes, S.M.F.; Hernandes, L.; Batista Junior, M.L.; Peres, S.B. Resistance exercise training induces subcutaneous and visceral adipose tissue browning in Swiss mice. J. Appl. Physiol. 2020, 129, 66–74. [Google Scholar] [CrossRef]
- Boon, M.R.; Bakker, L.E.; van der Linden, R.A.; van Ouwerkerk, A.F.; de Goeje, P.L.; Counotte, J.; Jazet, I.M.; Rensen, P.C. High prevalence of cardiovascular disease in South Asians: Central role for brown adipose tissue? Crit. Rev. Clin. Lab. Sci. 2015, 52, 150–157. [Google Scholar] [CrossRef]
- Thoonen, R.; Hindle, A.G.; Scherrer-Crosbie, M. Brown adipose tissue: The heat is on the heart. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1592–H1605. [Google Scholar] [CrossRef] [Green Version]
- Thoonen, R.; Ernande, L.; Cheng, J.; Nagasaka, Y.; Yao, V.; Miranda-Bezerra, A.; Chen, C.; Chao, W.; Panagia, M.; Sosnovik, D.E.; et al. Functional brown adipose tissue limits cardiomyocyte injury and adverse remodeling in catecholamine-induced cardiomyopathy. J. Mol. Cell Cardiol. 2015, 84, 202–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z.; Chen, M.; Tao, X.; Liu, Y.; He, J.; Wang, T.; Li, X. Synergistic Treatment of Obesity via Locally Promoting Beige Adipogenesis and Antioxidative Defense in Adipose Tissues. ACS Biomater. Sci. Eng. 2021, 7, 727–738. [Google Scholar] [CrossRef]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.; Oates, P.J.; Hammes, H.P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Dlaskova, A.; Clarke, K.J.; Porter, R.K. The role of UCP 1 in production of reactive oxygen species by mitochondria isolated from brown adipose tissue. Biochim. Biophys. Acta 2010, 1797, 1470–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceicao, G.; Matos, J.; Miranda-Silva, D.; Goncalves, N.; Sousa-Mendes, C.; Goncalves, A.; Ferreira, R.; Leite-Moreira, A.F.; Vitorino, R.; Falcao-Pires, I. Fat Quality Matters: Distinct Proteomic Signatures Between Lean and Obese Cardiac Visceral Adipose Tissue Underlie its Differential Myocardial Impact. Cell Physiol. Biochem. 2020, 54, 384–400. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Chen, R.; Wu, Y.; Huang, B.; Tang, C.; Chen, J.; Wang, Q.; Wu, Q.; Yang, J.; Qiu, H.; et al. PPARgamma-PI3K/AKT-NO signal pathway is involved in cardiomyocyte hypertrophy induced by high glucose and insulin. J. Diabetes Complicat. 2015, 29, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Qi, Y.; Gao, C. Cardiac myocyte-protective effect of microRNA-22 during ischemia and reperfusion through disrupting the caveolin-3/eNOS signaling. Int. J. Clin. Exp. Pathol. 2015, 8, 4614–4626. [Google Scholar] [PubMed]
- MacDougall, D.A.; Pugh, S.D.; Bassi, H.S.; Lotteau, S.; Porter, K.E.; Calaghan, S. Simvastatin Promotes Cardiac Myocyte Relaxation in Association with Phosphorylation of Troponin I. Front. Pharmacol. 2017, 8, 203. [Google Scholar] [CrossRef] [Green Version]
- Fadini, G.P.; de Kreutzenberg, S.; Agostini, C.; Boscaro, E.; Tiengo, A.; Dimmeler, S.; Avogaro, A. Low CD34+ cell count and metabolic syndrome synergistically increase the risk of adverse outcomes. Atherosclerosis 2009, 207, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Nana, Y.; Peng, J.; Jianlin, Z.; Xiangjian, Z.; Shutong, Y.; Enxin, Z.; Bin, L.; Chuanlong, Z.; Hua, T.; Yanhong, S.; et al. Reverse-D-4F Increases the Number of Endothelial Progenitor Cells and Improves Endothelial Progenitor Cell Dysfunctions in High Fat Diet Mice. PLoS ONE 2015, 10, e0138832. [Google Scholar] [CrossRef]
- Segal, M.S.; Shah, R.; Afzal, A.; Perrault, C.M.; Chang, K.; Schuler, A.; Beem, E.; Shaw, L.C.; Li Calzi, S.; Harrison, J.K.; et al. Nitric oxide cytoskeletal-induced alterations reverse the endothelial progenitor cell migratory defect associated with diabetes. Diabetes 2006, 55, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.; Dressler, N.; Ben-Shushan, R.S.; Meerson, A.; LeBaron, T.W.; Tamir, S. Effects of alkaline-electrolyzed and hydrogen-rich water, in a high-fat-diet nonalcoholic fatty liver disease mouse model. World J. Gastroenterol. 2018, 24, 5095–5108. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda, H.; Sato, A.; Miyata, K.; Shizuno, T.; Oyamada, A.; Ishiwata, K.; Nakagawa, Y.; Asahara, T. Drinking Molecular Hydrogen Water Is Beneficial to Cardiovascular Function in Diet-Induced Obesity Mice. Biology 2021, 10, 364. https://doi.org/10.3390/biology10050364
Masuda H, Sato A, Miyata K, Shizuno T, Oyamada A, Ishiwata K, Nakagawa Y, Asahara T. Drinking Molecular Hydrogen Water Is Beneficial to Cardiovascular Function in Diet-Induced Obesity Mice. Biology. 2021; 10(5):364. https://doi.org/10.3390/biology10050364
Chicago/Turabian StyleMasuda, Haruchika, Atsuko Sato, Kumiko Miyata, Tomoko Shizuno, Akira Oyamada, Kazuo Ishiwata, Yoshihiro Nakagawa, and Takayuki Asahara. 2021. "Drinking Molecular Hydrogen Water Is Beneficial to Cardiovascular Function in Diet-Induced Obesity Mice" Biology 10, no. 5: 364. https://doi.org/10.3390/biology10050364
APA StyleMasuda, H., Sato, A., Miyata, K., Shizuno, T., Oyamada, A., Ishiwata, K., Nakagawa, Y., & Asahara, T. (2021). Drinking Molecular Hydrogen Water Is Beneficial to Cardiovascular Function in Diet-Induced Obesity Mice. Biology, 10(5), 364. https://doi.org/10.3390/biology10050364