
Deletion of Mitochondrial Translocator Protein (TSPO) Gene Decreases Oxidative Retinal Pigment Epithelial Cell Death via Modulation of TRPM2 Channel



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Experimental Design
2.3. Obtaining TSPO Knockout (ARPE19-KO) Cells
2.4. The Determination of Intracellular Free Ca2+ Concentration ([Ca2+]c)
2.5. Patch-Clamp Electrophysiology Analyses
2.6. The Assay of Cell Viability and Apoptosis
2.7. The Assays of Caspase -3 (Casp -3) and -9 (Casp -3)
2.8. The Analyses of Mitochondrial fROS Generations
2.9. The Assay of the Mitochondrial Membrane Depolarization (mMDP)
2.10. The Analyses of Western Blot
2.11. The Analysis of Reduced Glutathione (rGSH), Glutathione Peroxidase (GSHPx) and Lipid Peroxidation (Malondialdehyde, MDA)
2.12. Statistical Analysis
3. Results
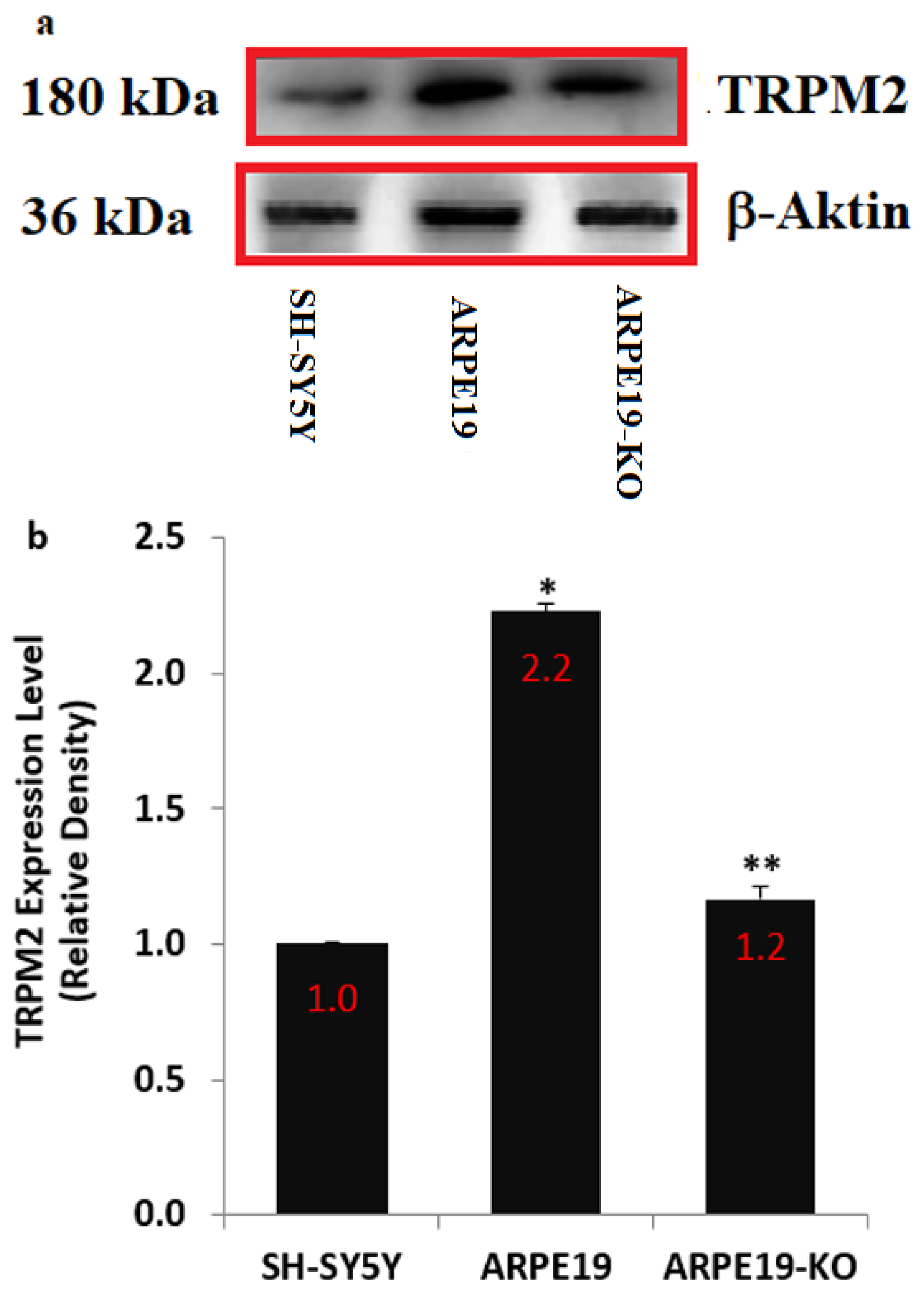
3.1. The Presence of Nature TRPM2 in the ARPE19 Cells Was Confirmed by Using the Positive Control Cells (SH-SY5Y)
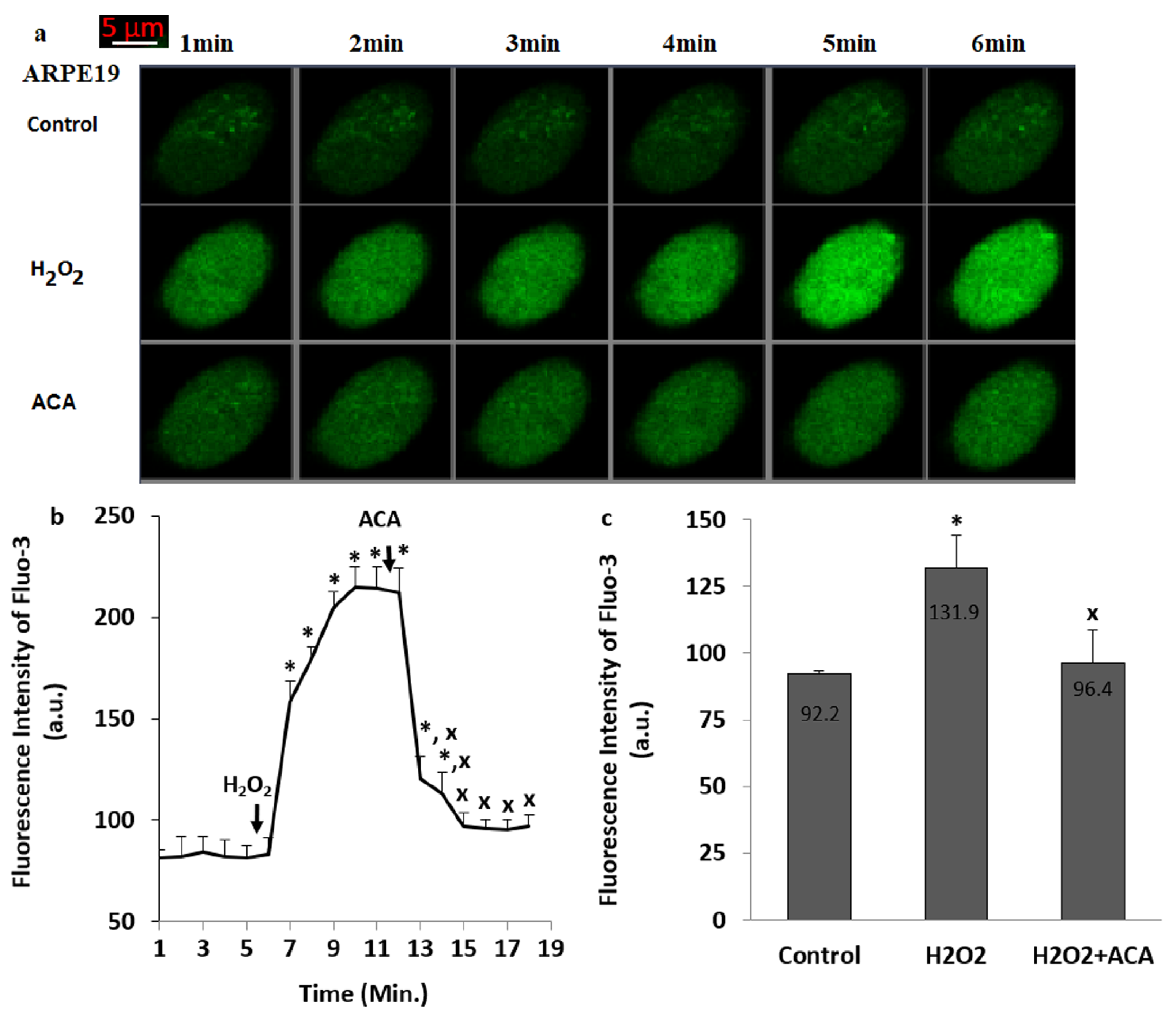
3.2. The TRPM2 Was Activated in the ARPE19 by the Stimulation of H2O2
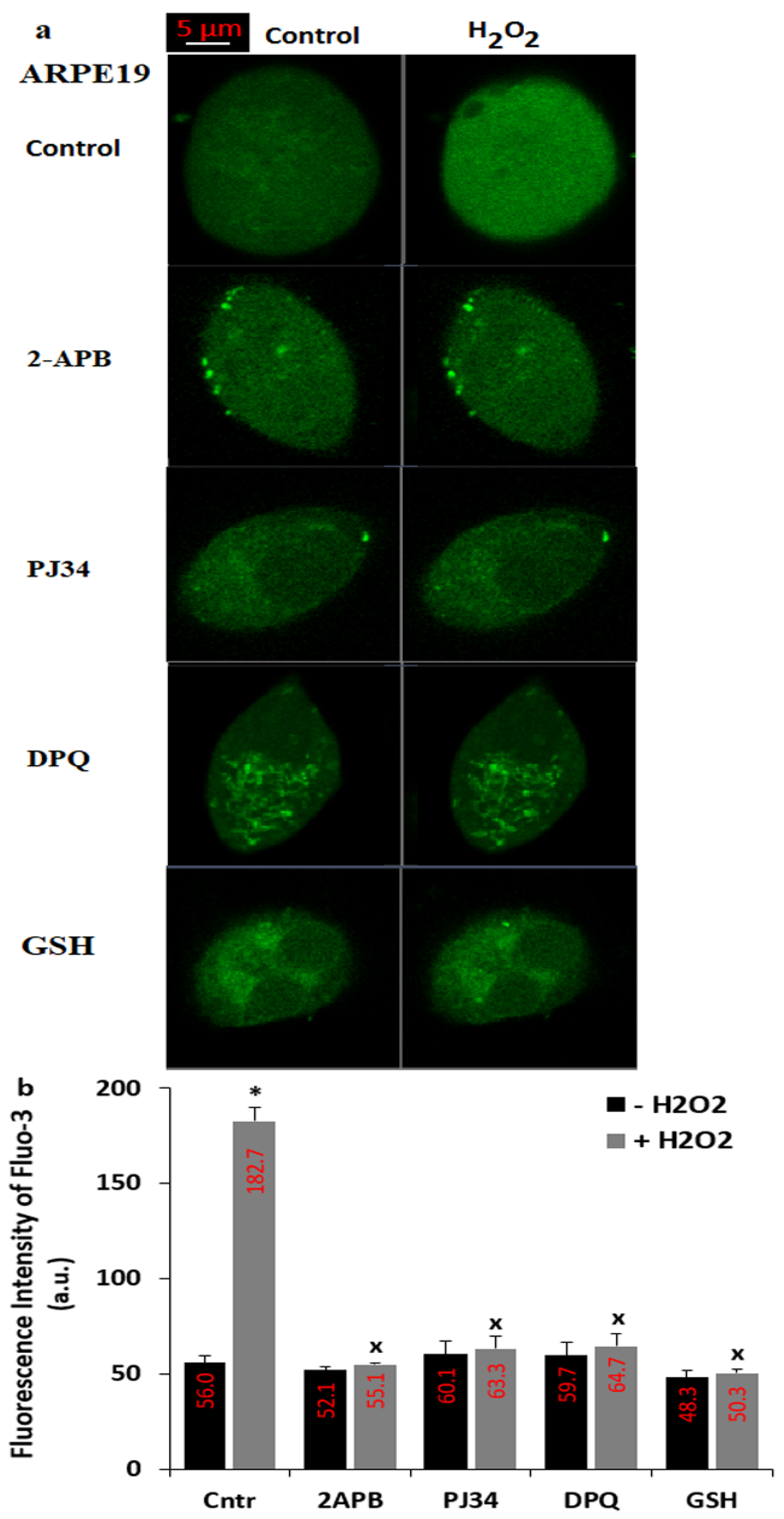
3.3. The H2O2-Mediated Upregulation of [Ca2+]c in the ARPE19 Was Diminished by the Treatments of PARP-1 Inhibitors (PJ34 and DPQ), TRPM2 Blocker (2APB) and GSH
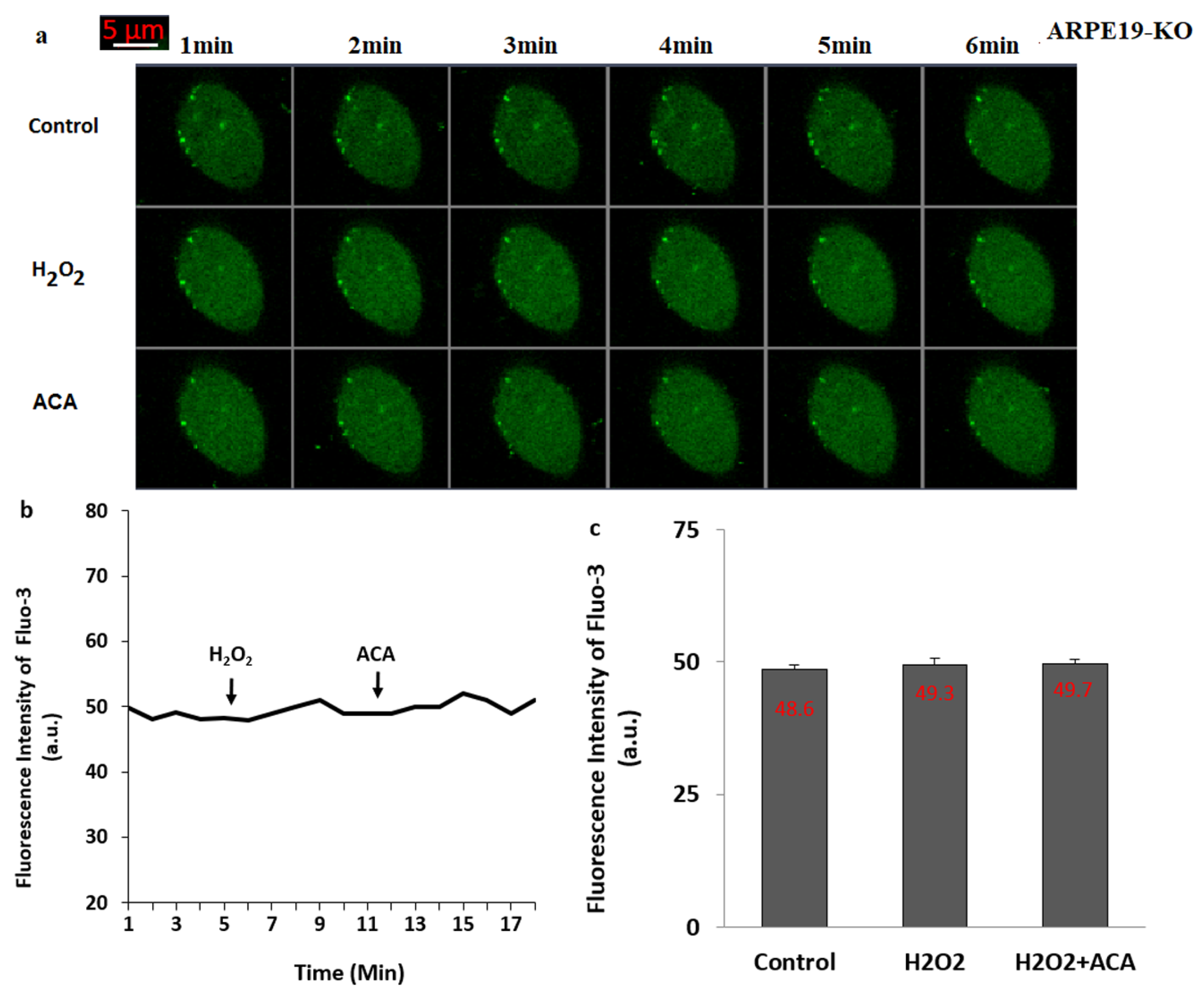
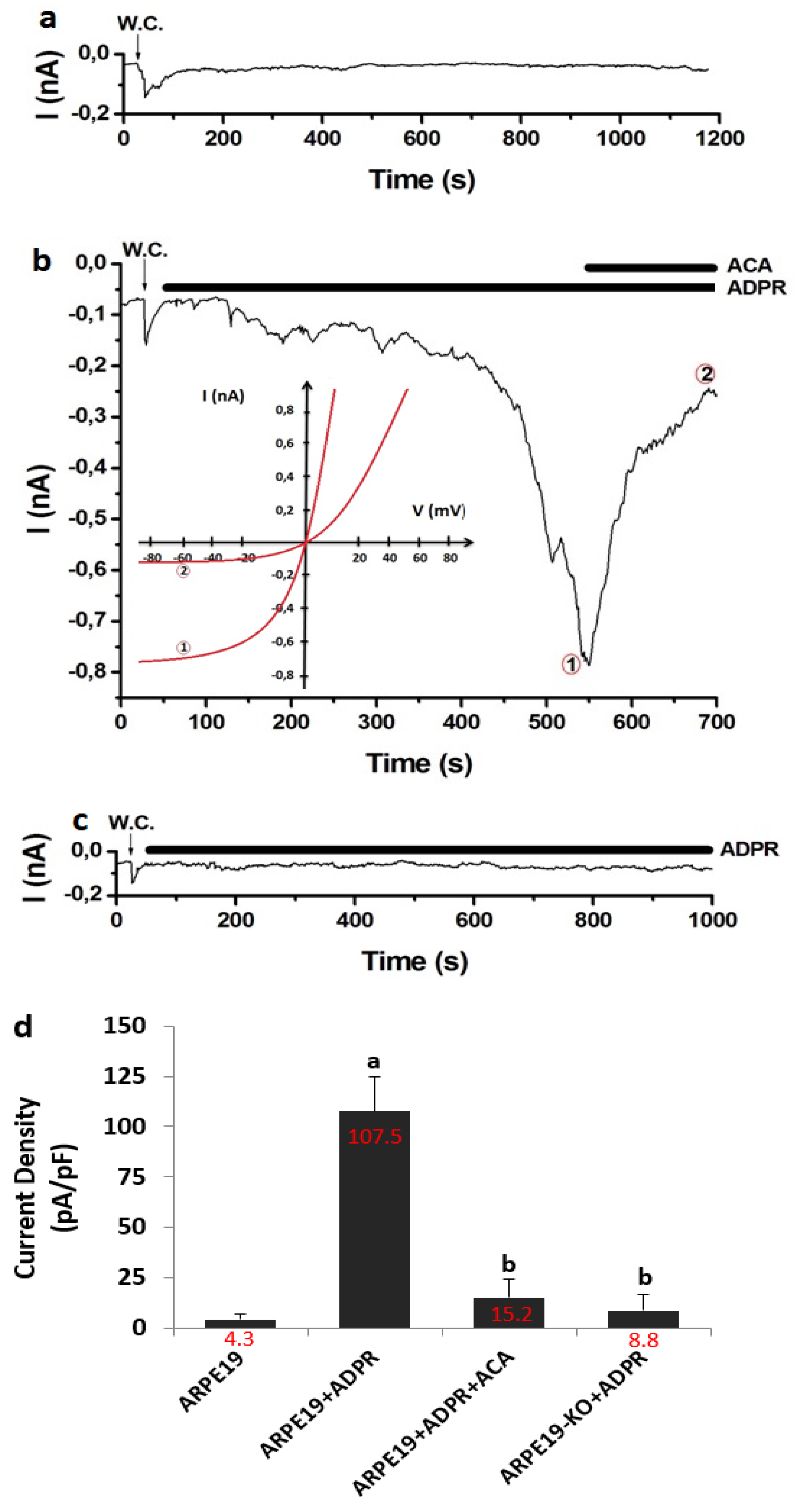
3.4. The Treatment of ADPR Induced TRPM2 Activation in the ARPE19, But Not in the ARPE19-KO
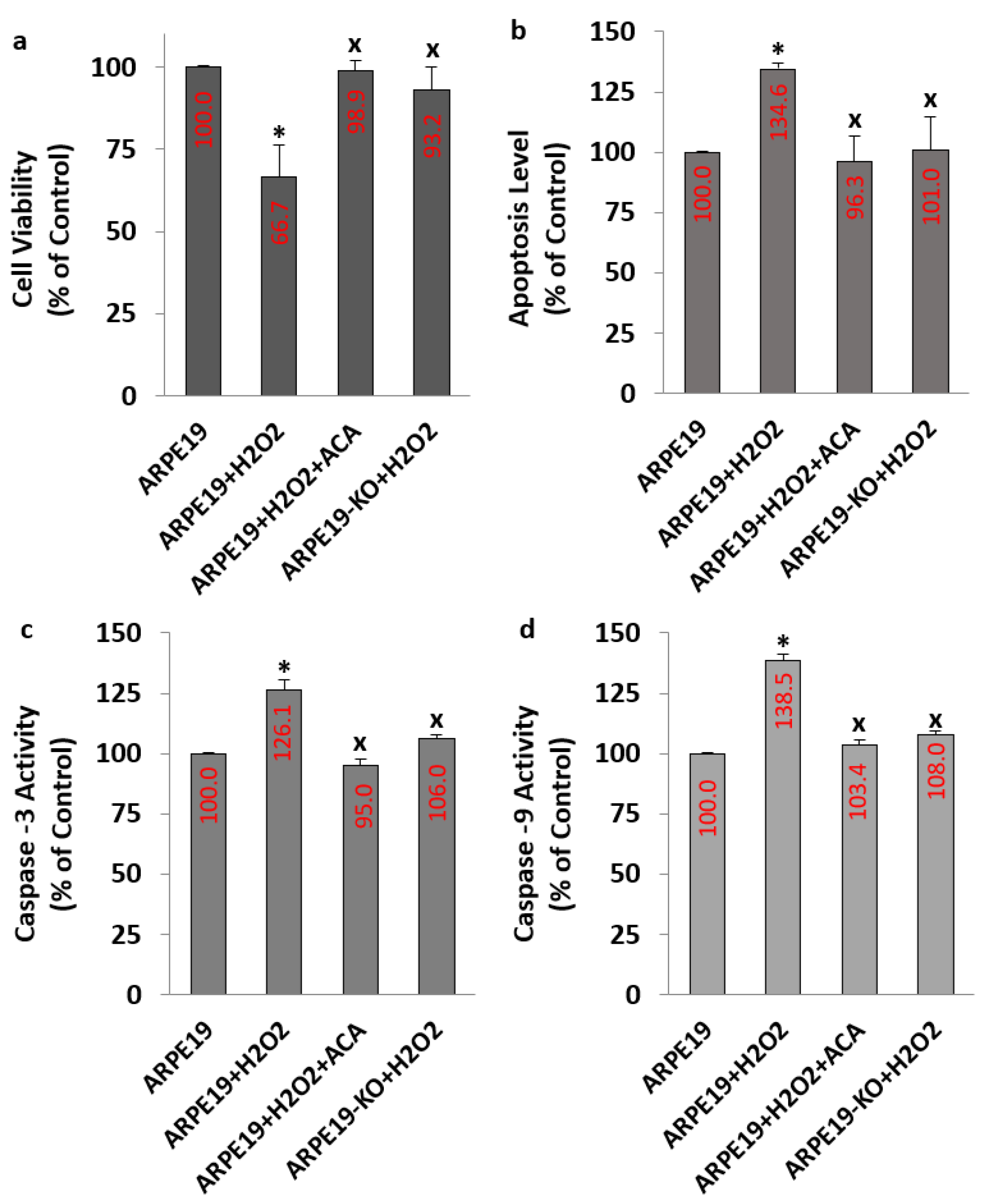
3.5. The Levels of Apoptosis, Casp -3, Casp -9 and Cell Viability Were Affected in the ARPE19 But Not in the ARPE19-KO Cells by the H2O2 Stimulation: Modulator Role of ACA
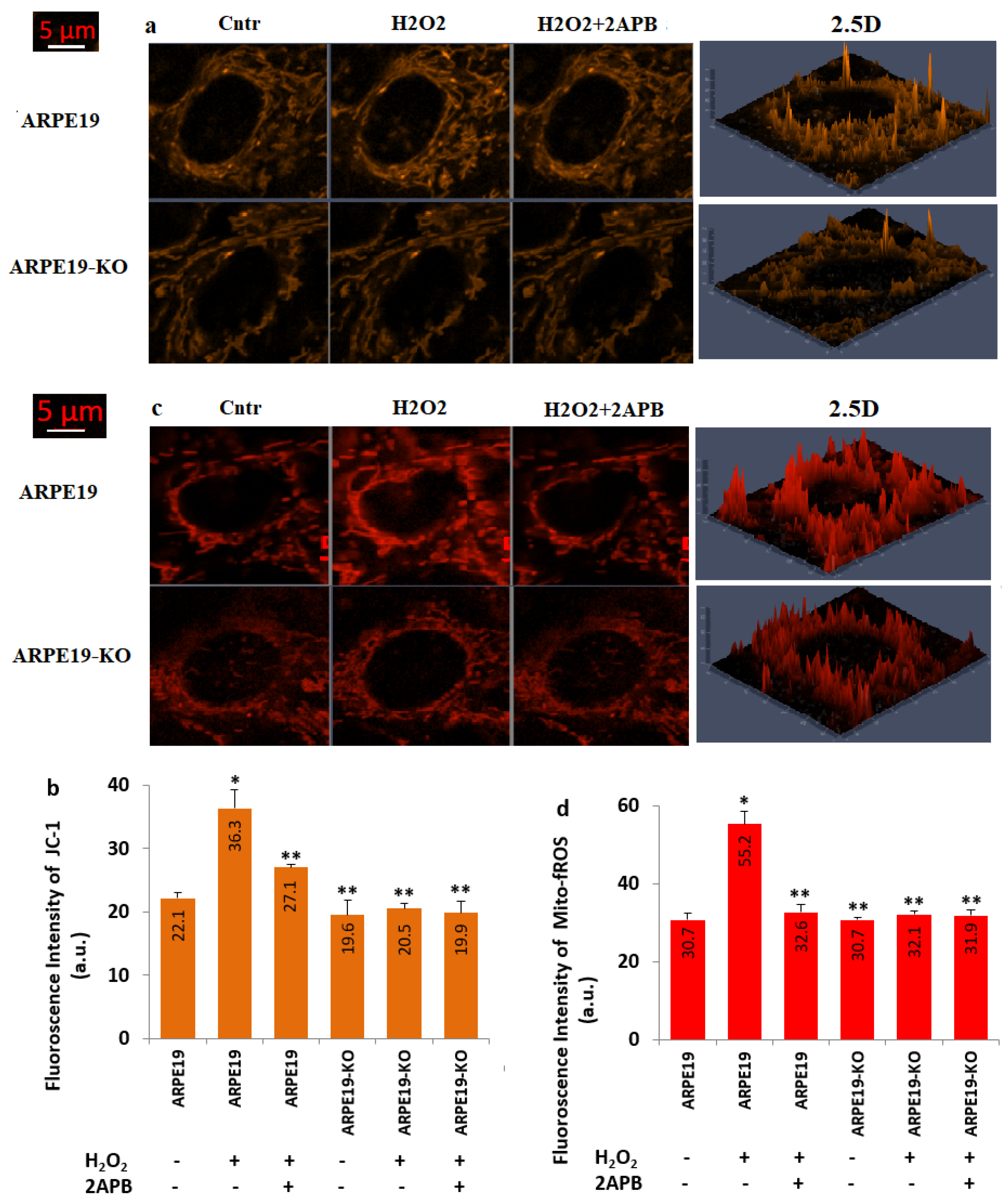
3.6. H2O2-Induced Increase of mMDP and Mito-fROS Levels Were Diminished in the ARPE19 Cells by the Treatment of 2APB
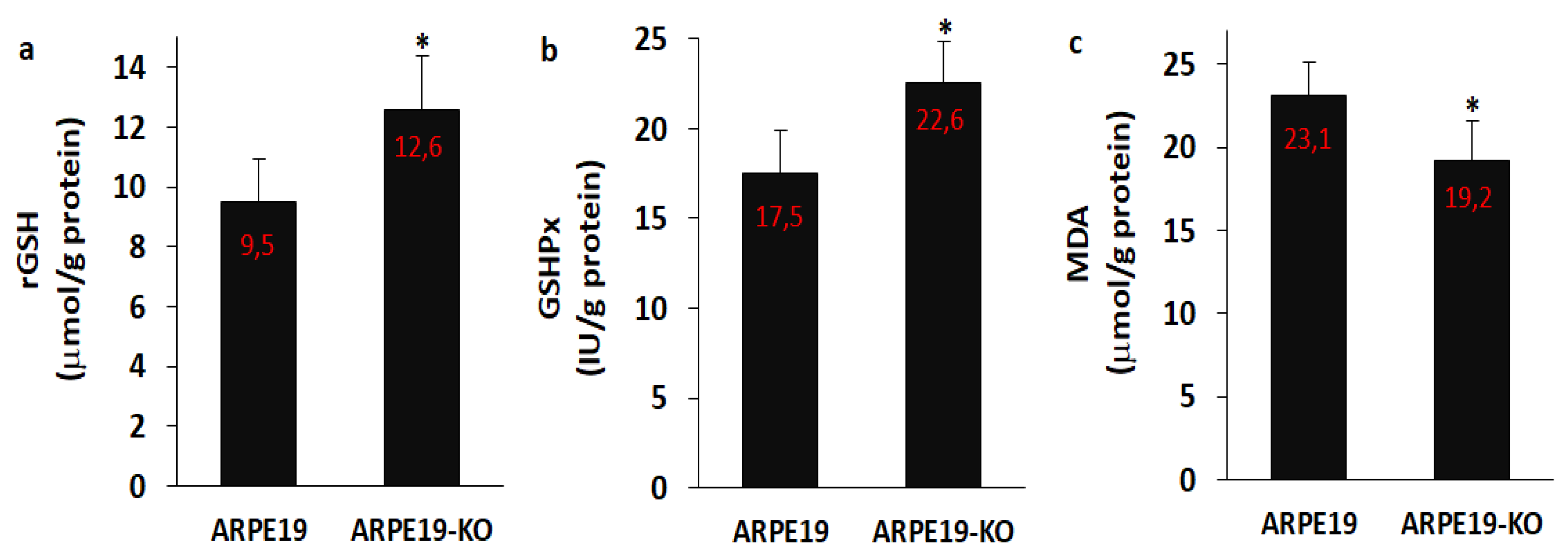
3.7. The Levels of rGSH, GSHPx and MDA Were Modulated in the ARPE19 by the Deletion of TSPO
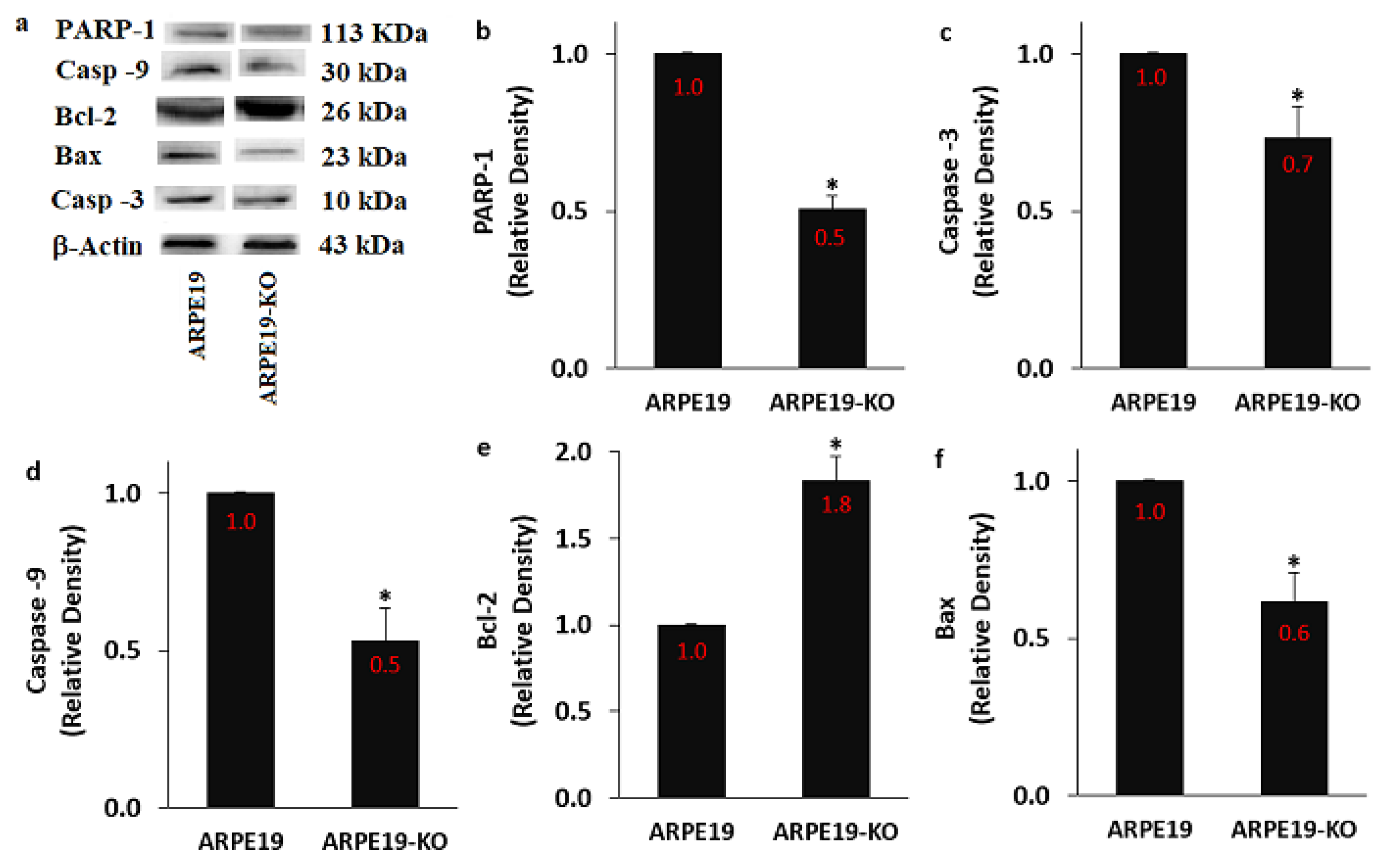
3.8. The Expression Levels of PARP-1, Bax, Bcl-2, Casp -3, and Casp -9 Were Modulated in the ARPE19 by the Deletion of TSPO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| [Ca2+]c | concentration of intracellular free calcium ion |
| 2APB | 2-aminoethoxydiphenyl borate |
| ACA | N-(p-amylcinnamoyl)anthranilic acid |
| ADPR | ADP-ribose |
| AMD | age-related macular degeneration |
| ARPE19 | adult retinal pigment epithelial 19 |
| Casp -3 | caspase -3 |
| Casp -9 | caspase -9 |
| Ca2+ | calcium ion |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| fROS | free reactive oxygen species |
| GSH | glutathione |
| GSHPx | glutathione peroxidase |
| h-RPE | human retinal pigment epithelium |
| MDA | malondialdehyde |
| mMDP | mitochondrial membrane depolarization |
| mPTP | mitochondrial permeability transition pore |
| rGSH | reduced glutathione |
| TRP | transient receptor potential |
| TRPM2 | transient receptor potential melastatin 2 |
| TSPO | 18 kDa mitochondrial translocator protein |
| VDCC | anion protein of voltage-dependent channel |
References
- Sreekumar, P.G.; Kannan, R. Mechanisms of protection of retinal pigment epithelial cells from oxidant injury by humanin and other mitochondrial-derived peptides: Implications for age-related macular degeneration. Redox Biol. 2020, 37, 101663. [Google Scholar] [CrossRef]
- Pennington, K.L.; DeAngelis, M.M. Epidemiology of age-related macular degeneration (AMD): Associations with cardiovascular disease phenotypes and lipid factors. Eye Vis. 2016, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Koc, F.; Erden, V.; Yurdakul, N.S. Causes of low vision and blindness in a Turkish adult population: The Izmir eye study. East. Mediterr. Health J. 2018, 24, 161–168. [Google Scholar] [CrossRef]
- Alamri, A.; Biswas, L.; Watson, D.G.; Shu, X. Deletion of TSPO Resulted in Change of Metabolomic Profile in Retinal Pigment Epithelial Cells. Int. J. Mol. Sci. 2019, 20, 1387. [Google Scholar] [CrossRef] [Green Version]
- Cougnard-Grégoire, A.; Delyfer, M.-N.; Korobelnik, J.-F.; Rougier, M.-B.; Le Goff, M.; Dartigues, J.-F.; Barberger-Gateau, P.; Delcourt, C. Elevated High-Density Lipoprotein Cholesterol and Age-Related Macular Degeneration: The Alienor Study. PLoS ONE 2014, 9, e90973. [Google Scholar] [CrossRef] [Green Version]
- Cano, M.; Fijalkowski, N.; Kondo, N.; Dike, S.; Handa, J. Advanced Glycation Endproduct Changes to Bruch’s Membrane Promotes Lipoprotein Retention by Lipoprotein Lipase. Am. J. Pathol. 2011, 179, 850–859. [Google Scholar] [CrossRef]
- Tuzcu, M.; Orhan, C.; Muz, O.; Sahin, N.; Juturu, V.; Sahin, K. Lutein and zeaxanthin isomers modulates lipid metabolism and the inflammatory state of retina in obesity-induced high-fat diet rodent model. BMC Ophthalmol. 2017, 17, 129. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Kambhampati, S.P.; Bhutto, I.A.; McLeod, D.S.; Lutty, G.A.; Kannan, R.M. Evolution of oxidative stress, inflammation and neovascularization in the choroid and retina in a subretinal lipid induced age-related macular degeneration model. Exp. Eye Res. 2021, 203, 108391. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M. New Molecular Mechanisms on the Activation of TRPM2 Channels by Oxidative Stress and ADP-Ribose. Neurochem. Res. 2007, 32, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Diagnostic and Therapeutic Potential of TSPO Studies Regarding Neurodegenerative Diseases, Psychiatric Disorders, Alcohol Use Disorders, Traumatic Brain Injury, and Stroke: An Update. Cells 2020, 9, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; You, L.; Ni, B.; Sai, N.; Wang, W.; Sun, M.; Xu, R.; Yao, Y.; Zhang, Z.; Qu, C.; et al. Phillyrin Mitigates Apoptosis and Oxidative Stress in Hydrogen Peroxide-Treated RPE Cells through Activation of the Nrf2 Signaling Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, V.M.; Slim, D.; Bader, S.; Koch, V.; Heinl, E.-S.; Alvarez-Carbonell, D.; Nothdurfter, C.; Rupprecht, R.; Wetzel, C.H. CRISPR-Cas9 Mediated TSPO Gene Knockout alters Respiration and Cellular Metabolism in Human Primary Microglia Cells. Int. J. Mol. Sci. 2019, 20, 3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, K.; Verhoyen, M.; Taiwo, M.; Langmann, T. Translocator protein (18 kDa) (TSPO) ligands activate Nrf2 signaling and attenuate inflammatory responses and oxidative stress in human retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 2020, 528, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Pittala, S.; Mizrachi, D. VDAC1 and the TSPO: Expression, Interactions, and Associated Functions in Health and Disease States. Int. J. Mol. Sci. 2019, 20, 3348. [Google Scholar] [CrossRef] [Green Version]
- Zeineh, N.; Denora, N.; Laquintana, V.; Franco, M.; Weizman, A.; Gavish, M. Efficaciousness of Low Affinity Compared to High Affinity TSPO Ligands in the Inhibition of Hypoxic Mitochondrial Cellular Damage Induced by Cobalt Chloride in Human Lung H1299 Cells. Biomedicines 2020, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Scholz, R.; Caramoy, A.; Bhuckory, M.B.; Rashid, K.; Chen, M.; Xu, H.; Grimm, C.; Langmann, T. Targeting translocator protein (18 kDa) (TSPO) dampens pro-inflammatory microglia reactivity in the retina and protects from degeneration. J. Neuroinflamm. 2015, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatliff, J.; East, A.D.; Singh, A.; Alvarez, M.S.; Frison, M.; Matic, I.; Ferraina, C.; Sampson, N.; Turkheimer, F.; Campanella, M. A role for TSPO in mitochondrial Ca2+ homeostasis and redox stress signaling. Cell Death Dis. 2017, 8, e2896. [Google Scholar] [CrossRef] [Green Version]
- Naziroğlu, M. Molecular role of catalase on oxidative stress-induced Ca2+signaling and TRP cation channel activation in nervous system. J. Recept. Signal Transduct. 2012, 32, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Nazıroğlu, M. TRPM2 Cation Channels, Oxidative Stress and Neurological Diseases: Where Are We Now? Neurochem. Res. 2010, 36, 355–366. [Google Scholar] [CrossRef]
- García, R.M.; Zamarripa, D.A.; Arnold, E.; Ruiz-Herrera, X.; Imm, R.N.; Cruz, G.B.; Adán, N.; Binart, N.; Riesgo-Escovar, J.; Goffin, V.; et al. Prolactin protects retinal pigment epithelium by inhibiting sirtuin 2-dependent cell death. EBioMedicine 2016, 7, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Özkaya, D.; Nazıroğlu, M.; Vanyorek, L.; Muhamad, S. Involvement of TRPM2 Channel on Hypoxia-Induced Oxidative Injury, Inflammation, and Cell Death in Retinal Pigment Epithelial Cells: Modulator Action of Selenium Nanoparticles. Biol. Trace Element Res. 2021, 199, 1356–1369. [Google Scholar] [CrossRef]
- Nazıroğlu, M. Activation of TRPM2 and TRPV1 Channels in Dorsal Root Ganglion by NADPH Oxidase and Protein Kinase C Molecular Pathways: A Patch Clamp Study. J. Mol. Neurosci. 2017, 61, 425–435. [Google Scholar] [CrossRef]
- Özkaya, D.; Nazıroğlu, M. Bevacizumab induces oxidative cytotoxicity and apoptosis via TRPM2 channel activation in retinal pigment epithelial cells: Protective role of glutathione. Graefe Arch. Clin. Exp. Ophthalmol. 2021, 1–16. [Google Scholar] [CrossRef]
- Wolf, A.; Herb, M.; Schramm, M.; Langmann, T. The TSPO-NOX1 axis controls phagocyte-triggered pathological angiogenesis in the eye. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Choi, J.; Ifuku, M.; Noda, M.; Guilarte, T.R. Translocator protein (18 kDa)/peripheral benzodiazepine receptor specific ligands induce microglia functions consistent with an activated state. Glia 2010, 59, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, K.; Aotaki-Keen, A.; Putkey, F.; Hjelmeland, L. ARPE-19, A Human Retinal Pigment Epithelial Cell Line with Differentiated Properties. Exp. Eye Res. 1996, 62, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Zhang, C.; Liu, D.; Wu, J.; Tian, H.; Lu, L.; Xu, G.-T.; Liu, F.; Zhang, J. Metformin Protects ARPE-19 Cells from Glyoxal-Induced Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 1740943. [Google Scholar]
- Chen, S.-J.; Hoffman, N.E.; Shanmughapriya, S.; Bao, L.; Keefer, K.; Conrad, K.; Merali, S.; Takahashi, Y.; Abraham, T.; Hirschler-Laszkiewicz, I.; et al. A Splice Variant of the Human Ion Channel TRPM2 Modulates Neuroblastoma Tumor Growth through Hypoxia-inducible Factor (HIF)-1/2α. J. Biol. Chem. 2014, 289, 36284–36302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Sukumaran, P.; Selvaraj, S.; Cilz, N.I.; Schaar, A.; Lei, S.; Singh, B.B. TRPM2 Promotes Neurotoxin MPP+/MPTP-Induced Cell Death. Mol. Neurobiol. 2018, 55, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Akyuva, Y.; Nazıroğlu, M. Resveratrol attenuates hypoxia-induced neuronal cell death, inflammation and mitochondrial oxidative stress by modulation of TRPM2 channel. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Armağan, H.H.; Nazıroğlu, M. Curcumin Attenuates Hypoxia-Induced Oxidative Neurotoxicity, Apoptosis, Calcium, and Zinc Ion Influxes in a Neuronal Cell Line: Involvement of TRPM2 Channel. Neurotox. Res. 2020. [Google Scholar] [CrossRef]
- Öz, A.; Çelik, Ö. Curcumin inhibits oxidative stress-induced TRPM2 channel activation, calcium ion entry and apoptosis values in SH-SY5Y neuroblastoma cells: Involvement of transfection procedure. Mol. Membr. Biol. 2016, 33, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Biswas, L.; Zhou, X.; Dhillon, B.; Graham, A.; Shu, X. Retinal pigment epithelium cholesterol efflux mediated by the 18 kDa translocator protein, TSPO, a potential target for treating age-related macular degeneration. Hum. Mol. Genet. 2017, 26, 4327–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, D.; Flemming, R.; Xu, S.-Z.; Perraud, A.-L.; Beech, D.J. Critical Intracellular Ca2+ Dependence of Transient Receptor Potential Melastatin 2 (TRPM2) Cation Channel Activation. J. Biol. Chem. 2003, 278, 11002–11006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perraud, A.-L.; Fleig, A.; Dunn, C.A.; Bagley, L.A.; Launay, P.; Schmitz, C.; Stokes, A.J.; Zhu, Q.; Bessman, M.J.; Penner, R.; et al. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nat. Cell Biol. 2001, 411, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M.; Lückhoff, A. A Calcium Influx Pathway Regulated Separately by Oxidative Stress and ADP-Ribose in TRPM2 Channels: Single Channel Events. Neurochem. Res. 2008, 33, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Mortadza, S.S.; Sim, J.A.; Stacey, M.; Jiang, L.-H. Signalling mechanisms mediating Zn2+-induced TRPM2 channel activation and cell death in microglial cells. Sci. Rep. 2017, 7, srep45032. [Google Scholar] [CrossRef]
- Han, D.; Wu, X.; Liu, L.; Shu, W.; Huang, Z. Sodium tanshinone IIA sulfonate protects ARPE-19 cells against oxidative stress by inhibiting autophagy and apoptosis. Sci. Rep. 2018, 8, 15137. [Google Scholar] [CrossRef] [Green Version]
- Bardak, H.; Uğuz, A.C.; Bardak, Y. Curcumin regulates intracellular calcium release and inhibits oxidative stress parameters, VEGF, and caspase-3/-9 levels in human retinal pigment epithelium cells. Physiol. Int. 2017, 104, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Papadopoulos, V. Mitochondrial TSPO Deficiency Triggers Retrograde Signaling in MA-10 Mouse Tumor Leydig Cells. Int. J. Mol. Sci. 2020, 22, 252. [Google Scholar] [CrossRef]
- Fan, J.; Wang, K.; Zirkin, B.; Papadopoulos, V. CRISPR/Cas9‒Mediated Tspo Gene Mutations Lead to Reduced Mitochondrial Membrane Potential and Steroid Formation in MA-10 Mouse Tumor Leydig Cells. Endocrinology 2018, 159, 1130–1146. [Google Scholar] [CrossRef] [Green Version]
- Zhao, A.H.; Tu, L.N.; Mukai, C.; Sirivelu, M.P.; Pillai, V.V.; Morohaku, K.; Cohen, R.; Selvaraj, V. Mitochondrial Translocator Protein (TSPO) Function Is Not Essential for Heme Biosynthesis. J. Biol. Chem. 2016, 291, 1591–1603. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Tian, M.; Yin, S.; Lai, S.; Zhou, Y.; Chen, J.; He, M.; Liao, Z. Downregulation of TSPO expression inhibits oxidative stress and maintains mitochondrial homeostasis in cardiomyocytes subjected to anoxia/reoxygenation injury. Biomed. Pharmacother. 2020, 121, 109588. [Google Scholar] [CrossRef]
- Carrasco, C.; Rodríguez, A.B.; Pariente, J.A. Melatonin as a stabilizer of mitochondrial function: Role in diseases and aging. Turk. J. Biol. 2015, 39, 822–831. [Google Scholar] [CrossRef]
- He, Y.; Tombran-Tink, J. Mitochondrial Decay and Impairment of Antioxidant Defenses in Aging RPE Cells. Adv. Exp. Med. Biol. 2009, 664, 165–183. [Google Scholar] [CrossRef]
- Rueda, C.B.; Llorente-Folch, I.; Traba, J.; Amigo, I.; Gonzalez-Sanchez, P.; Contreras, L.; Juaristi, I.; Martinez-Valero, P.; Pardo, B.; del Arco, A.; et al. Glutamate excitotoxicity and Ca2+-regulation of respiration: Role of the Ca2+ activated mitochondrial transporters (CaMCs). Biochim. Biophys. Acta (BBA) Bioenerg. 2016, 1857, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Schriewer, J.M.; Peek, C.B.; Bass, J.; Schumacker, P.T. ROS-Mediated PARP Activity Undermines Mitochondrial Function After Permeability Transition Pore Opening During Myocardial Ischemia–Reperfusion. J. Am. Hear. Assoc. 2013, 2, e000159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Sun, Y.; Wang, W.; Wang, B.; Zhang, Q.; Pan, C.; Yuan, Q.; Xu, F.; Wei, S.; Chen, Y. Poly (ADP-ribose) polymerase inhibition protects against myocardial ischaemia/reperfusion injury via suppressing mitophagy. J. Cell. Mol. Med. 2019, 23, 6897–6906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šileikytė, J.; Petronilli, V.; Zulian, A.; Dabbeni-Sala, F.; Tognon, G.; Nikolov, P.; Bernardi, P.; Ricchelli, F. Regulation of the Inner Membrane Mitochondrial Permeability Transition by the Outer Membrane Translocator Protein (Peripheral Benzodiazepine Receptor). J. Biol. Chem. 2011, 286, 1046–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klee, K.; Storti, F.; Barben, M.; Samardzija, M.; Langmann, T.; Dunaief, J.; Grimm, C. Systemic knockout of Tspo in mice does not affect retinal morphology, function and susceptibility to degeneration. Exp. Eye Res. 2019, 188, 107816. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özkaya, D.; Shu, X.; Nazıroğlu, M. Deletion of Mitochondrial Translocator Protein (TSPO) Gene Decreases Oxidative Retinal Pigment Epithelial Cell Death via Modulation of TRPM2 Channel. Biology 2021, 10, 382. https://doi.org/10.3390/biology10050382
Özkaya D, Shu X, Nazıroğlu M. Deletion of Mitochondrial Translocator Protein (TSPO) Gene Decreases Oxidative Retinal Pigment Epithelial Cell Death via Modulation of TRPM2 Channel. Biology. 2021; 10(5):382. https://doi.org/10.3390/biology10050382
Chicago/Turabian StyleÖzkaya, Dilek, Xinhua Shu, and Mustafa Nazıroğlu. 2021. "Deletion of Mitochondrial Translocator Protein (TSPO) Gene Decreases Oxidative Retinal Pigment Epithelial Cell Death via Modulation of TRPM2 Channel" Biology 10, no. 5: 382. https://doi.org/10.3390/biology10050382
APA StyleÖzkaya, D., Shu, X., & Nazıroğlu, M. (2021). Deletion of Mitochondrial Translocator Protein (TSPO) Gene Decreases Oxidative Retinal Pigment Epithelial Cell Death via Modulation of TRPM2 Channel. Biology, 10(5), 382. https://doi.org/10.3390/biology10050382