
Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Housing and Husbandry
2.2.1. Group-Housing
2.2.2. Pair-Housing
2.2.3. Relocation
2.3. Procedures
2.3.1. Hair Cortisol Analysis
2.3.2. Anthropometric Measurements
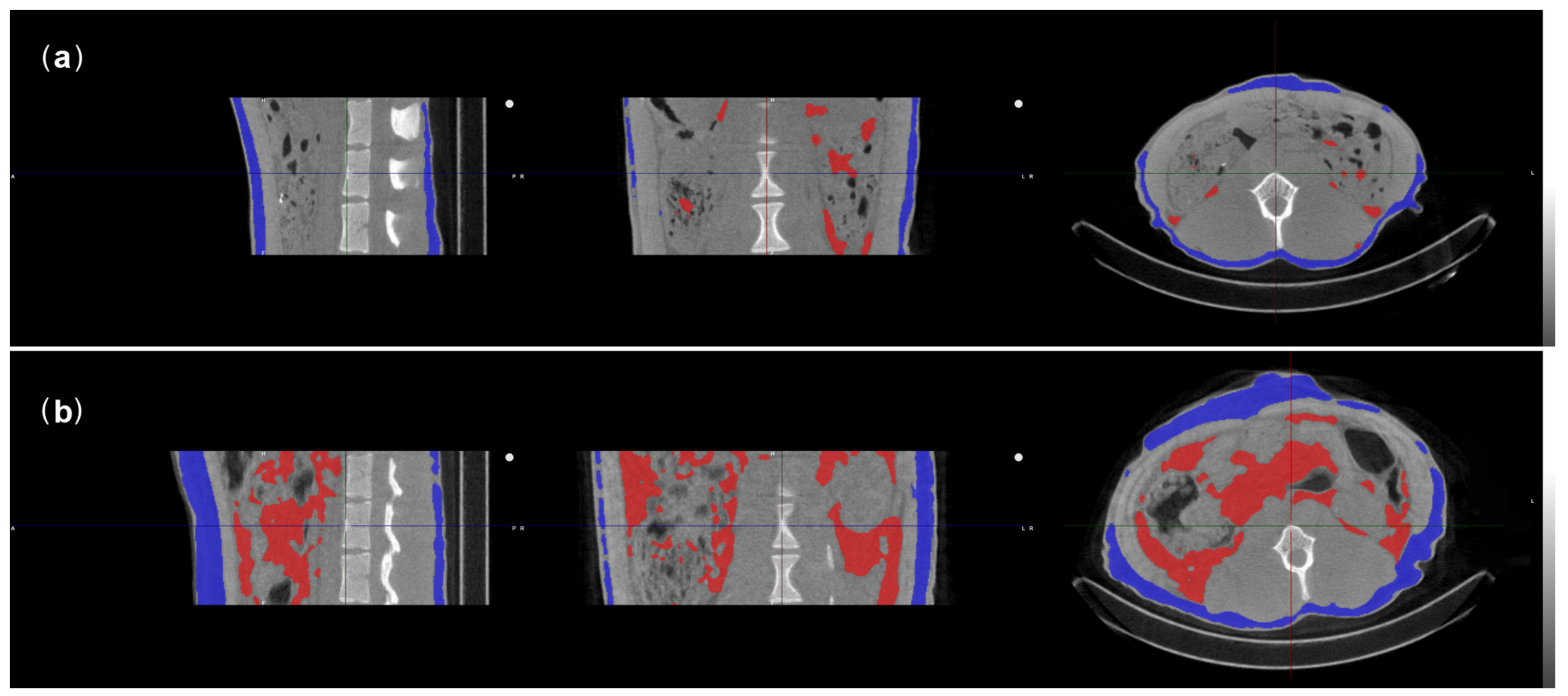
2.3.3. CTs
2.3.4. Dominance Rank
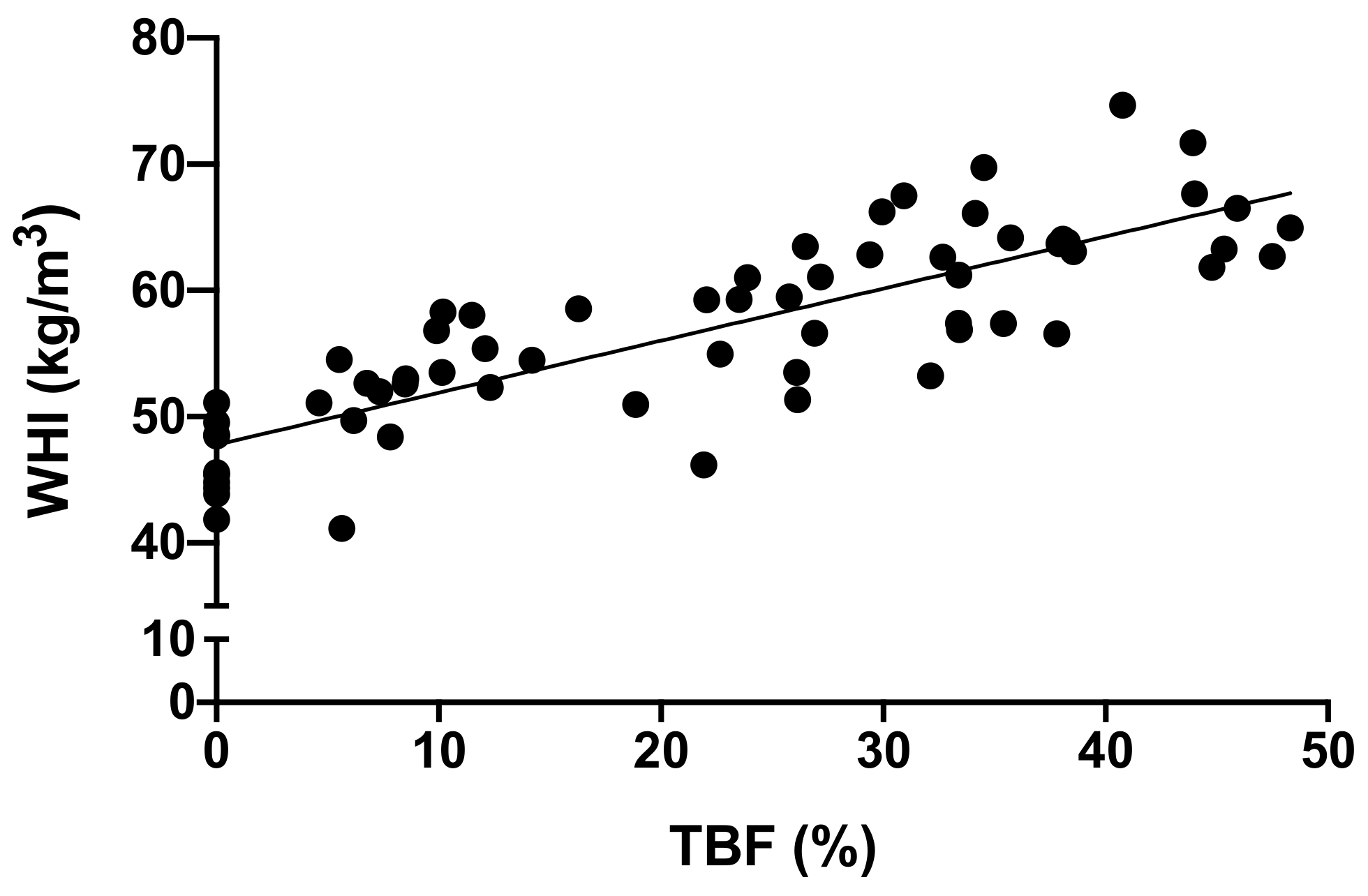
2.4. Data Analyses
3. Results
3.1. Group-Housing
3.2. Pair-Housing
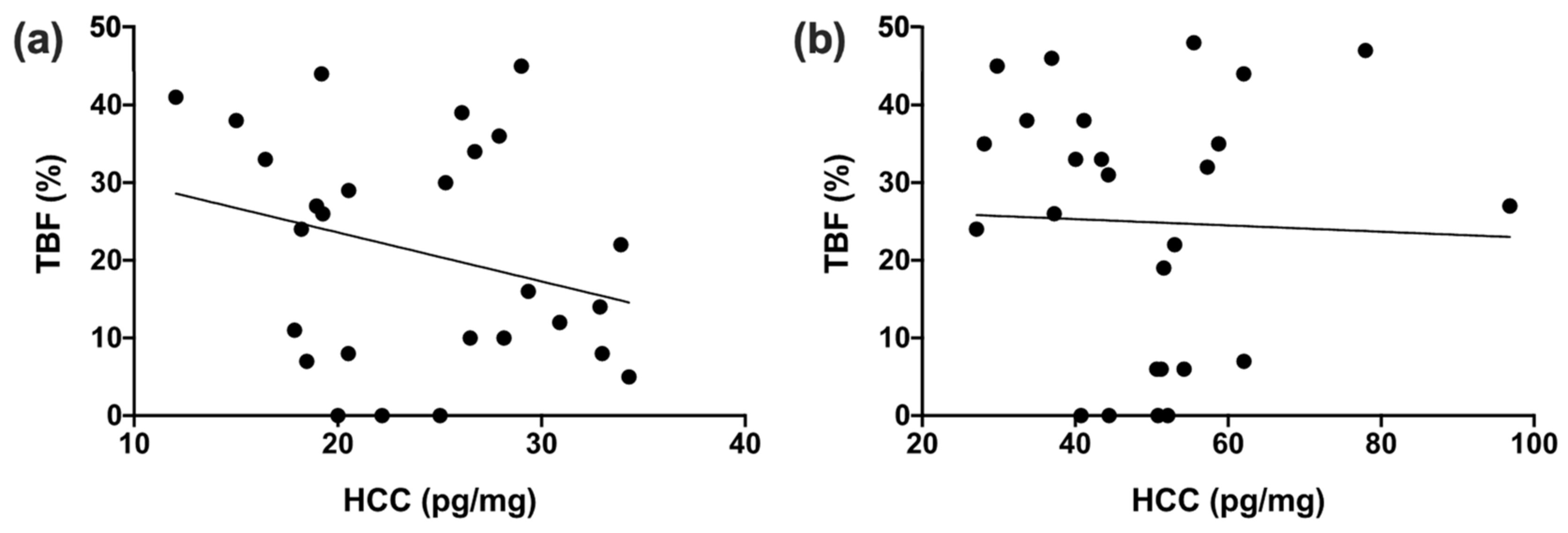
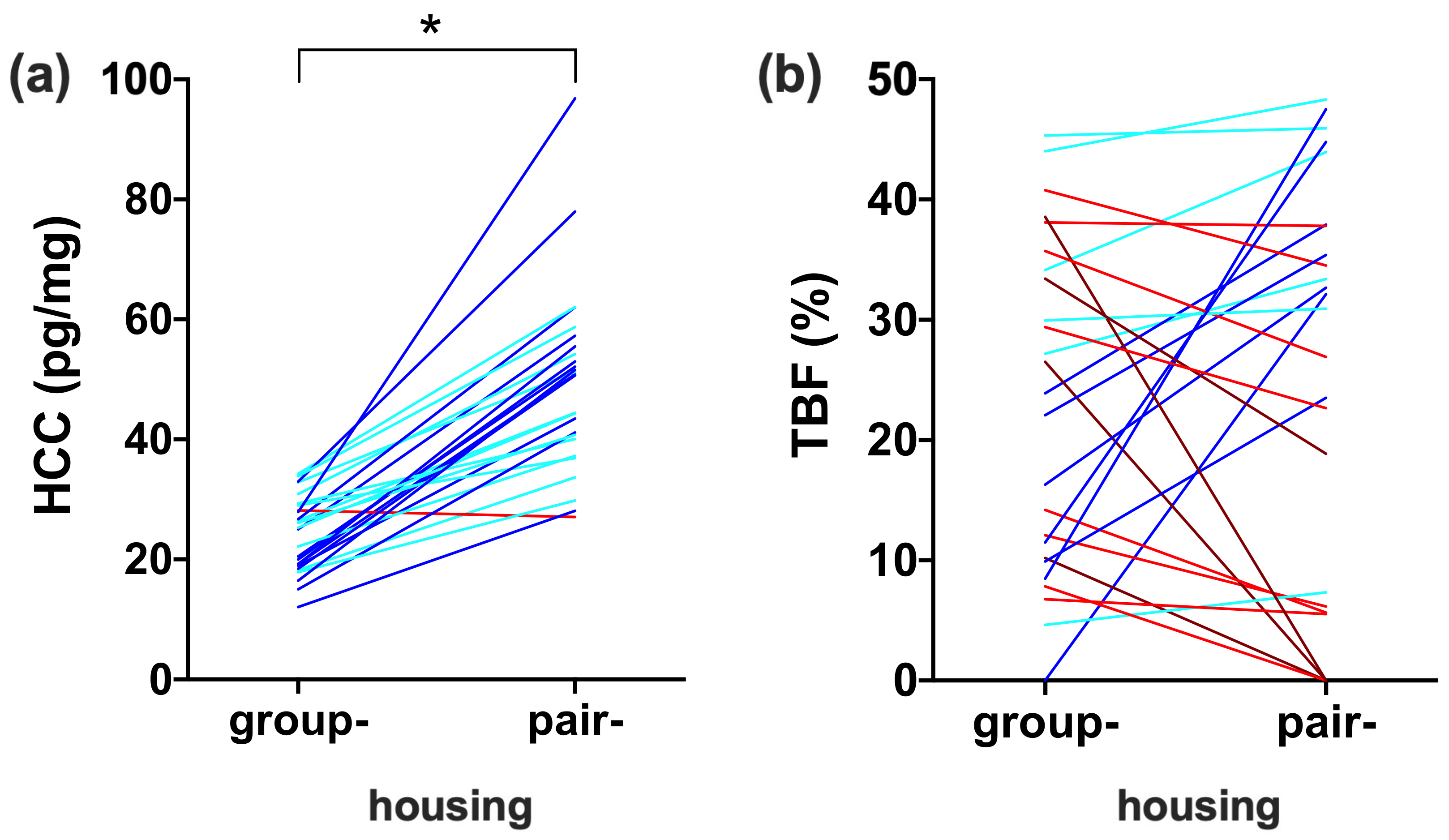
3.3. Comparison between Housing Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WHI | AC | AST | ABF | SBF | TBF | |
|---|---|---|---|---|---|---|
| BW | r = 0.857 # | r = 0.945 # | r = 0.847 # | r = 0.859 | r = 0.838 | r = 0.894 |
| WHI | X | r = 0.911 # | r = 0.861 # | r = 0.784 | r = 0.855 | r = 0.855 |
| AC | X | X | r = 0.905 # | r = 0.893 | r = 0.847 | r = 0.930 |
| ASF | X | X | X | r = 0.846 | r = 0.878 | r = 0.892 |
| ABF | X | X | X | X | r = 0.795 | r = 0.977 |
| SBF | X | X | X | X | X | r = 0.822 |
| Age | Group-Housed Dominance Rank | Pair-Housed Dominance Rank | Baseline Value | |
|---|---|---|---|---|
| BW | F = 0.965, p = 0.337 | F = 1.636, p = 0.214 | F = 0.219, p = 0.644 | F = 10.261, p = 0.004 ** |
| WHI | F = 0.367, p = 0.551 | F = 3.409, p = 0.078 * | F = 0.440, p = 0.514 | F = 4.487, p = 0.046 ** |
| AC | F = 0.535, p = 0.472 | F = 4.048, p = 0.057 * | F = 0.106, p = 0.748 | F = 8.764, p = 0.007 ** |
| ASF | F = 1.296, p = 0.267 | F = 3.526, p = 0.074 * | F = 0.044, p = 0.836 | F = 9.229, p = 0.006 ** |
| ABF | F = 1.697, p = 0.206 | F = 4.594, p = 0.043 ** | F = 0.005, p = 0.946 | F = 10.936, p = 0.003 ** |
| SBF | F = 0.019, p = 0.892 | F = 2.433, p = 0.133 | F = 0.144, p = 0.708 | F = 4.033, p = 0.057 * |
| TBF | F = 0.955, p = 0.339 | F = 4.282, p = 0.050 * | F = 0.010, p = 0.923 | F = 9.373, p = 0.006 ** |
| Delta WHI (N = 27) | Delta TBF (N = 27) | Delta HCC (N = 26) | |
|---|---|---|---|
| Number of procedures (0/5/8) | F = 0.397, p = 0.677 | F = 0.862, p = 0.435 | F = 0.365, p = 0.698 |
| Number of injuries (0/1/2/3) | F = 0.132, p = 0.940 | F = 0.288, p = 0.833 | F = 1.801, p = 0.176 |
| Exposure to humans (high vs. low) | t = 0.570, p = 0.575 | t = 0.365, p = 0.719 | t = −0.122, p = 0.912 |
| Cage size (single vs. double) | t = 0.547, p = 0.590 | t = 0.484, p = 0.633 | t = 1.867, p = 0.074 |
| Related pair (yes/no) | t = −0.181, p = 0.858 | t = −0.369, p = 0.715 | t = −0.950, p = 0.351 |
| Trainability (high vs. low) | t = −0.801, p = 0.431 | t = −1.988, p = 0.058 | t = 0.573, p = 0.572 |
| Measure | Group-Housing | Pair-Housing | Change |
|---|---|---|---|
| BW | 7.9 ± 0.2 (5.9–10.1) | 7.7 ± 0.2 (5.6–9.6) | −0.2 ± 0.2 (−2.5–1.7) |
| WHI | 58.3 ± 1.4 (43.9–74.7) | 56.3 ± 1.6 (41.1–71.7) | −2.0 ± 1.4 (−18.1–10.1) |
| AC | 35.3 ± 1.0 (26.2–43.7) | 35.0 ± 1.0 (26.9–44.4) | −0.3 ± 1.0 (−11.2–8.9) |
| ASF | 9.9 ± 0.8 (3.0–17.9) | 9.7 ± 1.0 (1.9–20.3) | −0.2 ± 1.0 (−12.7–9.5) |
| ABF | 11.9 ± 1.9 (0–32.2) | 14.8 ± 2.2 (0–40.5) | 3.0 ± 2.4 (−26.6–35.1) |
| SBF | 9.3 ± 1.1 (0–17.1) | 10.0 ± 1.2 (0–20.7) | 0.7 ± 1.2 (−13.9–11.5) |
| TBF | 21.1 ± 2.8 (0–45.3) | 24.8 ± 3.1 (0–48.3) | 3.7 ± 3.4 (−38.5–39.0) |
Appendix B

References
- Pieters, F. Use of primates. In Non-Human Primates in Biomedical Research—Reasons and Alternatives for Their Use, 1st ed.; Science shop for Biology: Utrecht, The Netherlands, 2007; pp. 13–28. [Google Scholar]
- Estes, J.D.; Wong, S.W.; Brenchley, J.M. Nonhuman primate models of human viral infections. Nat. Rev. Immunol. 2018, 18, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, J.; Bennett, B.T. Russell and Burch’s 3Rs then and now: The need for clarity in definition and purpose. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 120–132. [Google Scholar]
- Olsson, I.A.S.; Westlund, K. More than numbers matter: The effect of social factors on behaviour and welfare of laoratory rodents and non-human primates. Appl. Anim. Behav. Sci. 2007, 103, 229–254. [Google Scholar] [CrossRef]
- Cassidy, L.C.; Hannibal, D.L.; Semple, S.; McCowan, B. Improved behavioral indices of welfare in continuous compared to intermittent pair-housing in adult female rhesus macaques (Macaca mulatta). Am. J. Primatol. 2020, 82, e23189. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. The Psychological Well-Being of Nonhuman Primates; National Academies Collection: Reports funded by National Institutes of Health; National Academies Press: Washington, DC, USA, 1998. [Google Scholar]
- Jennings, M.; Prescott, M.J.; Buchanan-Smith, H.M.; Gamble, M.R.; Gore, M.; Hawkins, P.; Hubrecht, R.; Hudson, S.; Jennings, M.; Members of the Joint Working Group on Refinement (Primates); et al. Refinements in husbandry, care and common procedures for non-human primates: Ninth report of the BVAAWF/FRAME/RSPCA/UFAW Joint Working Group on Refinement. Lab. Anim. 2009, 43 (Suppl. 1), 1–47. [Google Scholar] [CrossRef]
- McCann, C.; Buchanan-Smith, H.M.; Jones-Engel, L.; Farmer, K.H.; Prescott, M.J.; Fitch-Snyder, H.; Taylor, S. IPS International Guidelines for the Acquisition, Care and Breeding of Nonhuman Primates; International Primatological Society: Chicago, IL, USA, 2007. [Google Scholar]
- Novak, M.A.; O’Neill, P.L.; Beckley, S.A.; Suomi, S.J. Naturalistic Environments for Captive Primates. In Naturalistic Environments in Captivity for Animal Behavior Research; Gibbons, E.F.J., Wyers, E.J., Waters, E., Menzel, E.W.J., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 236–258. [Google Scholar]
- O’Neill, P.L.; Novak, M.A.; Suomi, S.J. Normalizing laboratory-reared rhesus macaque (Macaca mulatta) behavior with exposure to complex outdoor enclosures. Zoo Biol. 1991, 10, 237–245. [Google Scholar] [CrossRef]
- Clarke, A.S.; Juno, C.J.; Maple, T.L. Behavioral Effects of a Change in the Physical Envirnoment: A Pilot Study of Captive Chimpanzees. Zoo Biol. 1982, 1, 371–380. [Google Scholar] [CrossRef]
- Ross, S.R.; Calcutt, S.; Schapiro, S.J.; Hau, J. Space use selectivity by chimpanzees and gorillas in an indoor-outdoor enclosure. Am. J. Primatol. 2011, 73, 197–208. [Google Scholar] [CrossRef]
- Hannibal, D.L.; Bliss-Moreau, E.; Vandeleest, J.; McCowan, B.; Capitanio, J. Laboratory rhesus macaque social housing and social changes: Implications for research. Am. J. Primatol. 2017, 79, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Schapiro, S.J.; Lambeth, S.P.; Jacobsen, K.R.; Williams, L.E.; Nehete, B.N.; Nehete, P.N. Physiological and Welfare Consequences of Transport, Relocation, and Acclimatization of Chimpanzees (Pan troglodytes). Appl. Anim. Behav. Sci. 2012, 137, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Snowdon, C.T. The significance od naturalistic environments for primate behavioral research. In Naturalistic Environments in Captivity for Animal Behavior ResearchI; Gibbons, E.F.J., Wyers, E.J., Waters, E., Menzel, E.W.J., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 217–235. [Google Scholar]
- Hau, J.; Shapiro, S.J. The welfare of non-human primates. In The Welfare of Laboratory Animals; Springer: Dordrecht, The Netherlands, 2007; pp. 291–314. [Google Scholar]
- Clarke, A.S.; Mason, W.A.; Mendoza, S.P. Heart rate patterns under stress in three species of macaques. Am. J. Primatol. 1994, 33, 133–148. [Google Scholar] [CrossRef]
- Clarke, A.S.; Mason, W.A.; Moberg, G.P. Differentiall behavioral and adrenocortical responses to stress among three macaque species. Am. J. Primatol. 1988, 14, 37–52. [Google Scholar] [CrossRef]
- Cohen, S.; Kaplan, J.R.; Cunnick, J.E.; Manuck, S.B.; Rabin, B.S. Chronic Social Stress, Affiliation, and Cellular Immune Response in Nonhuman Primates. Psychol. Sci. 1992, 3, 301–304. [Google Scholar] [CrossRef]
- Fernstrom, A.L.; Sutian, W.; Royo, F.; Westlund, K.; Nilsson, T.; Carlsson, H.E.; Paramastri, Y.; Pamungkas, J.; Sajuthi, D.; Schapiro, S.J.; et al. Stress in cynomolgus monkeys (Macaca fascicularis) subjected to long-distance transport and simulated transport housing conditions. Stress 2008, 11, 467–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gust, D.A.; Gordon, T.P.; Brodie, A.R.; McClure, H.M. Effect of a preferred companion in modulating stress in adult female rhesus monkeys. Physiol. Behav. 1994, 55, 681–684. [Google Scholar] [CrossRef]
- Cheney, D.L.; Seyfarth, R.M. Stress and Coping Mechanisms in Female Primates. Adv. Study Behav. 2009, 39, 1–44. [Google Scholar]
- Davenport, M.D.; Lutz, C.K.; Tiefenbacher, S.; Novak, M.A.; Meyer, J.S. A rhesus monkey model of self-injury: Effects of relocation stress on behavior and neuroendocrine function. Biol. Psychiatry 2008, 63, 990–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honess, P.E.; Johnson, P.J.; Wolfensohn, S.E. A study of behavioural responses of non-human primates to air transport and re-housing. Lab. Anim. 2004, 38, 119–132. [Google Scholar] [CrossRef]
- Bethea, C.L.; Centeno, M.L.; Cameron, J.L. Neurobiology of stress-induced reproductive dysfunction in female macaques. Mol. Neurobiol. 2008, 38, 199–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncharova, N.D.; Oganyan, T.E. Age-related differences in stress responsiveness of the hypothalamic-pituitary-adrenal axis of nonhuman primates with various types of adaptive behavior. Gen. Comp. Endocrinol. 2018, 258, 163–172. [Google Scholar] [CrossRef]
- Cohen, S.; Line, S.; Manuck, S.B.; Rabin, B.S.; Heise, E.R.; Kaplan, J.R. Chronic social stress, social status, and susceptibility to upper respiratory infections in nonhuman primates. Psychosom. Med. 1997, 59, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Davenport, M.D.; Tiefenbacher, S.; Lutz, C.K.; Novak, M.A.; Meyer, J.S. Analysis of endogenous cortisol concentrations in the hair of rhesus macaques. Gen. Comp. Endocrinol. 2006, 147, 255–261. [Google Scholar] [CrossRef]
- Bauer, S.A.; Arndt, T.P.; Leslie, K.E.; Pearl, D.L.; Turner, P.V. Obesity in rhesus and cynomolgus macaques: A comparative review of the condition and its implications for research. Comp. Med. 2011, 61, 514–526. [Google Scholar]
- Shively, C.A.; Register, T.C.; Clarkson, T.B. Social stress, visceral obesity, and coronary artery atherosclerosis: Product of a primate adaptation. Am. J. Primatol. 2009, 71, 742–751. [Google Scholar] [CrossRef] [Green Version]
- Jayo, J.M.; Shively, C.A.; Kaplan, J.R.; Manuck, S.B. Effects of exercise and stress on body fat distribution in male cynomolgus monkeys. Int. J. Obes. Relat. Metab. Disords 1993, 17, 597–604. [Google Scholar]
- Rhyu, I.J.; Bytheway, J.A.; Kohler, S.J.; Lange, H.; Lee, K.J.; Boklewski, J.; McCormick, K.; Williams, N.I.; Stanton, G.B.; Greenough, W.T.; et al. Effects of aerobic exercise training on cognitive function and cortical vascularity in monkeys. Neuroscience 2010, 167, 1239–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crumley, D.; Raleigh, M.J. Social status and fat distribution in male vervet monkeys. Am. J. Primatol. 1995, 36, 118–119. [Google Scholar]
- Vernes, M.K.; Louwerse, A.L. (Eds.) BPRC’s Enrichment Manual for Macaques & Marmosets; Biomedical Primate Research Centre: Rijswijk, The Netherlands, 2010; Available online: https://www.bprc.nl/sites/default/files/downloads/Enrichment%20Manual.pdf (accessed on 2 August 2021).
- Sterck, E.H.M.; Zijlmans, D.G.M.; de Vries, H.; van den Berg, L.M.; van Schaik, C.P.; Langermans, J.A.M. Determining overweight and underweight with a new weight-for-height index in captive group-housed macaques. Am. J. Primatol. 2019, 81, e22996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittus, W.P. Arboreal adaptations of body fat in wild toque macaques (Macaca sinica) and the evolution of adiposity in primates. Am. J. Phys. Anthropol. 2013, 152, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimburge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef]
- Behringer, V.; Deschner, T. Non-invasive monitoring of physiological markers in primates. Horm. Behav. 2017, 91, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Wooddell, L.J.; Hamel, A.F.; Murphy, A.M.; Byers, K.L.; Kaburu, S.S.K.; Meyer, J.S.; Suomi, S.J.; Dettmer, A.M. Relationships between affiliative social behavior and hair cortisol concentrations in semi-free ranging rhesus monkeys. Psychoneuroendocrinology 2017, 84, 109–115. [Google Scholar] [CrossRef]
- Dettmer, A.M.; Novak, M.A.; Meyer, J.S.; Suomi, S.J. Population density-dependent hair cortisol concentrations in rhesus monkeys (Macaca mulatta). Psychoneuroendocrinology 2014, 42, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, H.E.; Leckman, J.F.; Coplan, J.D.; Suomi, S.J. Risk and resilience: Early manipulation of macaque social experience and persistent behavioral and neurophysiological outcomes. J. Am. Acad. Child. Adolesc. Psychiatry 2009, 48, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M. Social subordinance as a marker of hypercortisolism. Some unexpected subtleties. Ann. N. Y. Acad. Sci. 1995, 771, 626–639. [Google Scholar] [CrossRef]
- Shively, C.A. Social subordination stress, behavior, and central monoaminergic function in female cynomolgus monkeys. Biol. Psychiatry 1998, 44, 882–891. [Google Scholar] [CrossRef]
- Chamove, A.S.; Bowman, R.E. Rhesus plasma cortisol response at four dominance positions. Aggress. Behav. 1978, 4, 43–55. [Google Scholar] [CrossRef]
- Gust, D.A.; Gordon, T.P.; Hambright, M.K.; Wilson, M.E. Relationship between Social Factors and Pituitary-Adrenocortical Activity in Female Rhesus Monkeys (Macaca mulatta). Horm. Behav. 1993, 27, 318–331. [Google Scholar] [CrossRef]
- Abbott, D.H.; Keverne, E.B.; Bercovitch, F.B.; Shively, C.A.; Mendoza, S.P.; Saltzman, W.; Snowdon, C.T.; Ziegler, T.E.; Banjevic, M.; Garland, T., Jr.; et al. Are subordinates always stressed? A comparative analysis of rank differences in cortisol levels among primates. Horm. Behav. 2003, 43, 67–82. [Google Scholar] [CrossRef]
- Garcia, C.; Huffman, M.; Shimizu, K. Seasonal and reproductive variation in body condition in captive female Japanese macaques (Macaca fuscata). Am. J. Primatol. 2010, 72, 277–286. [Google Scholar] [CrossRef]
- Altmann, J.; Schoeller, D.; Altmann, S.A.; Muruthi, P.; Sapolsky, R.M. Body size and fatness of free-living baboons reflect food availability and activity levels. Am. J. Primatol. 1993, 30, 149–161. [Google Scholar] [CrossRef]
- Bodkin, N.L.; Hannah, J.S.; Ortmeyer, H.K.; Hansen, B.C. Central obesity in rhesus monkeys: Association with hyperinsulinemia, insulin resistance and hypertriglyceridemia? Int. J. Obes. Relat. Metab. Disords 1993, 17, 53–61. [Google Scholar]
- Colman, R.J.; Hudson, J.C.; Barden, H.S.; Kemnitz, J.W. A comparison of dual-energy X-ray absorptiometry and somatometrics for determining body fat in rhesus macaques. Obes. Res. 1999, 7, 90–96. [Google Scholar] [CrossRef]
- Kemnitz, J.W.; Weindruch, R.; Roecker, E.B.; Crawford, K.; Kaufman, P.L.; Ershler, W.B. Dietary restriction of adult male rhesus monkeys: Design, methodology, and preliminary findings from the first year of study. J. Gerontol. 1993, 48, B17–B26. [Google Scholar] [CrossRef]
- Walker, M.L.; Schwartz, S.M.; Wilson, M.E.; Musey, P.I. Estimation of body fat in female rhesus monkeys. Am. J. Phys. Anthropol. 1984, 63, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Laber-Laird, K.; Shively, C.A.; Karstaedt, N.; Bullock, B.C. Assessment of abdominal fat deposition in female cynomolgus monkeys. Int. J. Obes. 1991, 15, 213–220. [Google Scholar] [PubMed]
- Cefalu, W.T.; Wagner, J.D.; Wang, Z.Q.; Bell-Farrow, A.D.; Collins, J.; Haskell, D.; Bechtold, R.; Morgan, T. A study of caloric restriction and cardiovascular aging in cynomolgus monkeys (Macaca fascicularis): A potential model for aging research. J. Gerontol. A Biol. Sci. Med. Sci. 1997, 52, B10–B19. [Google Scholar] [CrossRef] [Green Version]
- Seidell, J.C.; Bakker, C.J.; van der Kooy, K. Imaging techniques for measuring adipose-tissue distribution—A comparison between computed tomography and 1.5-T magnetic resonance. Am. J. Clin. Nutr. 1990, 51, 953–957. [Google Scholar] [CrossRef]
- Wallace, J.M.; Shively, C.A.; Clarkson, T.B. Effects of hormone replacement therapy and social stress on body fat distribution in surgically postmenopausal monkeys. Int. J. Obes. 1999, 23, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizumi, T.; Nakamura, T.; Yamane, M.; Islam, A.H.; Menju, M.; Yamasaki, K.; Arai, T.; Kotani, K.; Funahashi, T.; Yamashita, S.; et al. Abdominal fat: Standardized technique for measurement at CT. Radiology 1999, 211, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooy, K. Changes in Body Composition and Fat Distribution in Response to Weight Loss and Weight Regain. Thesis of the University of Wageningen, 1993. [Google Scholar]
- Yue, F.; Zhang, G.; Quintero, J.E.; Gash, D.M.; Zhang, Z. Role of social interaction, exercise, diet, and age on developing and untreated diabetes in cynomolgus monkeys. Exp. Gerontol. 2017, 96, 82–88. [Google Scholar] [CrossRef]
- Small, M.F. Body fat, rank, and nutritional status in a captive group of rhesus macaques. Int. J. Primatol. 1981, 2, 91–95. [Google Scholar] [CrossRef]
- Whitten, P.L. Diet and dominance among female vervet monkeys (Cercopithecus aethiops). Am. J. Primatol. 1983, 5, 139–159. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zijlmans, D.G.M.; Meijer, L.; Vernes, M.K.; Wubben, J.A.M.; Hofman, L.; Louwerse, A.L.; Sterck, E.H.M.; Langermans, J.A.M.; Stammes, M.A. Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques. Biology 2021, 10, 744. https://doi.org/10.3390/biology10080744
Zijlmans DGM, Meijer L, Vernes MK, Wubben JAM, Hofman L, Louwerse AL, Sterck EHM, Langermans JAM, Stammes MA. Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques. Biology. 2021; 10(8):744. https://doi.org/10.3390/biology10080744
Chicago/Turabian StyleZijlmans, Dian G. M., Lisette Meijer, Marit K. Vernes, Jacqueline A. M. Wubben, Linda Hofman, Annet L. Louwerse, Elisabeth H. M. Sterck, Jan A. M. Langermans, and Marieke A. Stammes. 2021. "Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques" Biology 10, no. 8: 744. https://doi.org/10.3390/biology10080744
APA StyleZijlmans, D. G. M., Meijer, L., Vernes, M. K., Wubben, J. A. M., Hofman, L., Louwerse, A. L., Sterck, E. H. M., Langermans, J. A. M., & Stammes, M. A. (2021). Effect of Housing Conditions on Cortisol and Body Fat Levels in Female Rhesus Macaques. Biology, 10(8), 744. https://doi.org/10.3390/biology10080744