
Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Climate Change Effects on Plant Pathogens and Diseases
3. Ecological Roles of Microorganisms from Specific Disease-Suppressive Soils
4. Rhizobiome to the Service of Plant Health, When Plants “Cry for Help”
5. Core Microbiome—“Few but Good”
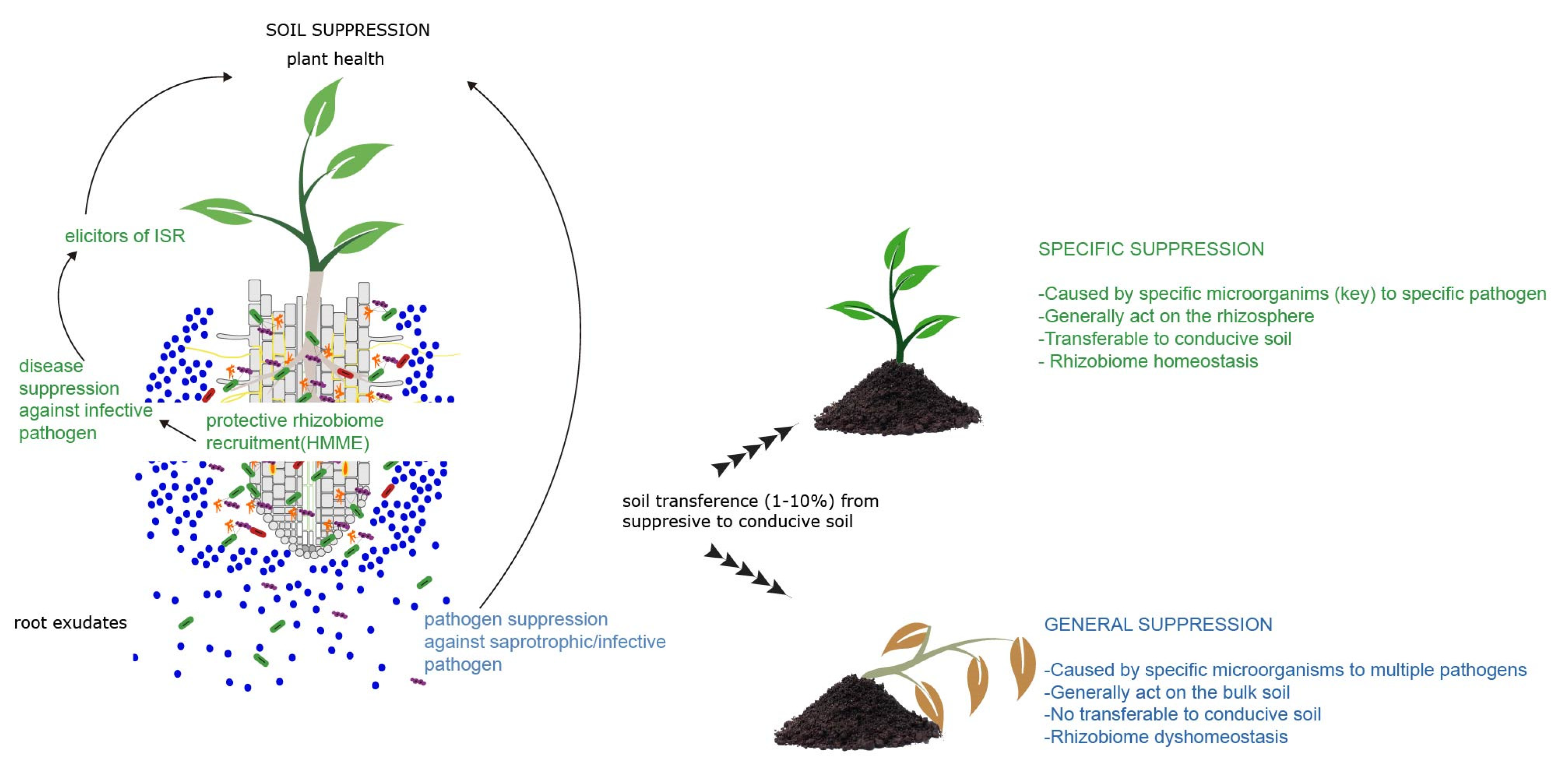
6. Exploiting Host-Mediated Microbiota Engineering for Protection of Plants against Diseases
7. The New Generation of Personalized Bioinoculants by Inducing the Host-Mediated Microbiota Engineering—Perspectives and Future Remarks
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Garrett, K.A.; Nita, M.; De Wolf, E.D.; Esker, P.D.; Gomez-Montano, L.; Sparks, A.H. Plant Pathogens as Indicators of Climate Change, 2nd ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2015; ISBN 9780444635242. [Google Scholar]
- Fones, H.N.; Gurr, S.J. NOX ious gases and the unpredictability of emerging plant pathogens under climate change. BMC Biol. 2017, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Daszak, P.; Cunningham, A.; Hyatt, A. Emerging Infectious Diseases of Wildlife- Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- French, S.; Levy-Booth, D.; Samarajeewa, A.; Shannon, K.E.; Smith, J.; Trevors, J.T. Elevated temperatures and carbon dioxide concentrations: Effects on selected microbial activities in temperate agricultural soils. World J. Microbiol. Biotechnol. 2009, 25, 1887–1900. [Google Scholar] [CrossRef]
- Meisner, A.; De Boer, W. Strategies to maintain natural biocontrol of soil-borne crop diseases during severe drought and rainfall events. Front. Microbiol. 2018, 9, 2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, A.L.; Davis, R. Internal decay of onions caused by Enterobacter cloacae. Plant Dis. 1990, 74, 692–694. [Google Scholar] [CrossRef]
- Wang, G.F.; Xie, G.L.; Zhu, B.; Huang, J.S.; Liu, B.; Kawicha, P.; Benyon, L.; Duan, Y.P. Identification and characterization of the Enterobacter complex causing mulberry (Morus alba) wilt disease in China. Eur. J. Plant Pathol. 2010, 126, 465–478. [Google Scholar] [CrossRef]
- Masyahit, M.; Sijam, K.; Awang, Y.; Satar, M.G.M. First report on bacterial soft rot disease on dragon fruit (Hylocereus spp.) caused by Enterobacter cloacae in Peninsular Malaysia. Int. J. Agric. Biol. 2009, 11, 659–666. [Google Scholar]
- Nishijima, K.A.; Wall, M.M.; Siderhurst, M.S. Demonstrating pathogenicity of Enterobacter cloacae on Macadamia and identifying associated volatiles of gray kernel of Macadamia in Hawaii. Plant Dis. 2007, 91, 1221–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Martinez, R.; de la Cerda, K.A.; Costa, H.S.; Cooksey, D.A.; Wong, F.P. Phylogenetic Relationships of Xylella fastidiosa Strains Isolated from Landscape Ornamentals in Southern California. Phytopathology 2007, 97, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M. Take-All Decline and Beneficial Pseudomonads. In Principles of Plant-Microbe Interactions; Springer: Cham, Germany, 2015; pp. 363–370. [Google Scholar] [CrossRef]
- Rivera-Becerril, F.; van Tuinen, D.; Chatagnier, O.; Rouard, N.; Béguet, J.; Kuszala, C.; Soulas, G.; Gianinazzi-Pearson, V.; Martin-Laurent, F. Impact of a pesticide cocktail (fenhexamid, folpel, deltamethrin) on the abundance of Glomeromycota in two agricultural soils. Sci. Total Environ. 2017, 577, 84–93. [Google Scholar] [CrossRef]
- Cook, R.J. Take-all of wheat. Physiol. Mol. Plant Pathol. 2003, 62, 73–86. [Google Scholar] [CrossRef]
- Andrade, O.; Campillo, R.; Peyrelongue, A. Soils suppressive against Gaeumannomyces graminis var. tritici identified under wheat crop monoculture in southern Chile. Cienc. e Investig. Agrar. 2011, 38, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Durán, P.; Jorquera, M.; Viscardi, S.; Carrion, V.J. Screening and Characterization of Potentially Suppressive Soils against Gaeumannomyces graminis under Extensive Wheat Cropping by Chilean Indigenous Communities. Front. Microbiol. 2017, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease Suppressive Soils: New Insights from the Soil Microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Mendes, L.W.; Raaijmakers, J.M.; De Hollander, M.; Mendes, R.; Tsai, S.M. Influence of resistance breeding in common bean on rhizosphere microbiome composition and function. ISME J. 2018, 12, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Strom, N.; Hu, W.; Haarith, D.; Chen, S.; Bushley, K. Interactions between soil properties, fungal communities, the soybean cyst nematode, and crop yield under continuous corn and soybean monoculture. Appl. Soil Ecol. 2020, 147, 103388. [Google Scholar] [CrossRef]
- Termorshuizen, A.J.; Jeger, M.J. Strategies of soilborne plant pathogenic fungi in relation to disease suppression. Fungal Ecol. 2008, 1, 108–114. [Google Scholar] [CrossRef]
- Duran, P.; Tortella, G.; Viscardi, S.; Barra, P.J.; Carrion, V.J.; Mora, M.D.L.L.; Pozo, M.J. Microbial Community Composition in Take-All Suppressive Soils. Front. Microbiol. 2018, 9, 2198. [Google Scholar] [CrossRef]
- Chng, S.; Cromey, M.G.; Dodd, S.L.; Stewart, A.; Butler, R.C.; Jaspers, M.V. Take-all decline in New Zealand wheat soils and the microorganisms associated with the potential mechanisms of disease suppression. Plant Soil 2015, 397, 239–259. [Google Scholar] [CrossRef]
- Mazzola, M. Mechanisms of natural soil suppressiveness to soilborne diseases. Antonie Van Leeuwenhoek 2002, 81, 557–564. [Google Scholar] [CrossRef]
- Benítez, M.S.; Tustas, F.B.; Rotenberg, D.; Kleinhenz, M.D.; Cardina, J.; Stinner, D.; Miller, S.A.; McSpadden Gardener, B.B. Multiple statistical approaches of community fingerprint data reveal bacterial populations associated with general disease suppression arising from the application of different organic field management strategies. Soil Biol. Biochem. 2007, 39, 2289–2301. [Google Scholar] [CrossRef]
- Kwak, Y.S.; Weller, D.M. Take-all of wheat and natural disease suppression: A review. Plant Pathol. J. 2013, 29, 125–135. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, X.; Li, L. Human gut microbiome: The second genome of human body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Khoruts, A.; Sadowsky, M.J. Understanding the mechanisms of faecal microbiota transplantation. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Haney, C.H.; Samuel, B.S.; Bush, J.; Ausubel, F.M. Associations with rhizosphere bacteria can confer an adaptive advantage to plants. Nat. Plants 2015, 1, 15051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Gao, Z.; Tian, B.; Bi, K.; Chen, T.; Liu, H.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. Endosphere microbiome comparison between symptomatic and asymptomatic roots of Brassica napus infected with Plasmodiophora brassicae. PLoS ONE 2017, 12, e0185907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Bae, B. Optimization of Biological Phosphorus Removal from Contaminated Sediments with Phosphate-Solubilizing Microorganisms. J. Biosci. Bioeng. 2005, 99, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Staley, C.; Ferrieri, A.P.; Tfaily, M.M.; Cui, Y.; Chu, R.K.; Wang, P.; Shaw, J.B.; Ansong, C.K.; Brewer, H.; Norbeck, A.D.; et al. Diurnal cycling of rhizosphere bacterial communities is associated with shifts in carbon metabolism. Microbiome 2017, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.; Bhuyan, S.K.; Yadava, P.K.; Varma, A.; Tuteja, N. Emergence of plant and rhizospheric microbiota as stable interactomes. Protoplasma 2017, 254, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Aleklett, K.; Leff, J.W.; Fierer, N.; Hart, M. Wild plant species growing closely connected in a subalpine meadow host distinct root-associated bacterial communities. PeerJ 2015, 2015, e804. [Google Scholar] [CrossRef] [Green Version]
- Samad, A.; Trognitz, F.; Compant, S.; Antonielli, L.; Sessitsch, A. Shared and host-specific microbiome diversity and functioning of grapevine and accompanying weed plants. Environ. Microbiol. 2017, 19, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Micallef, S.A.; Shiaris, M.P.; Colón-Carmona, A. Influence of Arabidopsis thaliana accessions on rhizobacterial communities and natural variation in root exudates. J. Exp. Bot. 2009, 60, 1729–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiza, L.; St-Arnaud, M.; Yergeau, E. Harnessing phytomicrobiome signaling for rhizosphere microbiome engineering. Front. Plant Sci. 2015, 6, 507. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Rocha-Granados, M.D.C.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Germida, J.J.; Siciliano, S.D.; de Freitas, J.R.; Seib, A.M. Diversity of root-associated bacteria associated with held-grown canola (Brassica napus L.) and wheat (Triticum aestivum L.). Fems Microbiol. Ecol. 1998, 26, 43–50. [Google Scholar] [CrossRef]
- Marquez-Santacruz, H.A.; Hernandez-Leon, R.; Orozco-Mosqueda, M.C.; Velazquez-Sepulveda, I.; Santoyo, G. Diversity of bacterial endophytes in roots of Mexican husk tomato plants (Physalis ixocarpa) and their detection in the rhizosphere. Genet. Mol. Res. 2010, 9, 2372–2380. [Google Scholar] [CrossRef]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Mueller, U.G.; Sachs, J.L. Engineering Microbiomes to Improve Plant and Animal Health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Rillig, M.C.; Tsang, A.; Roy, J. Microbial Community Coalescence for Microbiome Engineering. Front. Microbiol. 2016, 7, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to Consider Phytophthora infestans a Reemerging Pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Kumar, A.; Barman, M.; Pal, S.; Bandopadhyay, P. Impact of Climate Variability on Phenology of Rice. In Agronomic Crops; Springer Nature Singapore Pte Ltd.: Singapore, 2020; pp. 13–28. [Google Scholar] [CrossRef]
- Sevik, H.; Cetin, M.; Ozel, H.B.; Erbek, A.; Zeren Cetin, I. The effect of climate on leaf micromorphological characteristics in some broad-leaved species. Environ. Dev. Sustain. 2021, 23, 6395–6407. [Google Scholar] [CrossRef]
- Eastburn, D.M.; Degennaro, M.M.; Delucia, E.H.; Dermody, O.; Mcelrone, A.J. Elevated atmospheric carbon dioxide and ozone alter soybean diseases at SoyFACE. Glob. Chang. Biol. 2010, 16, 320–330. [Google Scholar] [CrossRef]
- Mhamdi, A.; Noctor, G. High CO2 primes plant biotic stress defences through redox-linked pathways. Plant Physiol. 2016, 172, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, X.; Sun, Z.; Shao, S.; Hu, L.; Ye, M.; Zhou, Y.; Xia, X.; Yu, J.; Shi, K. Antagonism between phytohormone signalling underlies the variation in disease susceptibility of tomato plants under elevated CO2. J. Exp. Bot. 2015, 66, 1951–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prank, M.; Kenaley, S.C.; Bergstrom, G.C.; Acevedo, M.; Mahowald, N.M. Climate change impacts the spread potential of wheat stem rust, a significant crop disease. Environ. Res. Lett. 2019, 14, 124053. [Google Scholar] [CrossRef] [Green Version]
- Hovmøller, M.S.; Yahyaoui, A.H.; Milus, E.A.; Justesen, A.F. Rapid global spread of two aggressive strains of a wheat rust fungus. Mol. Ecol. 2008, 17, 3818–3826. [Google Scholar] [CrossRef] [PubMed]
- Milus, E.A.; Kristensen, K.; Hovmøller, M.S. Evidence for Increased Aggressiveness in a Recent Widespread Strain of Puccinia striiformis f. sp. tritici Causing Stripe Rust of Wheat. Phytopathology 2009, 99, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Váry, Z.; Mullins, E.; Mcelwain, J.C.; Doohan, F.M. The severity of wheat diseases increases when plants and pathogens are acclimatized to elevated carbon dioxide. Glob. Chang. Biol. 2015, 21, 2661–2669. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ishiguro, K.; Nakajima, T.; Kim, H.Y.; Okada, M.; Kobayashi, K. Effects of Elevated Atmospheric CO2 Concentration on the Infection of Rice Blast and Sheath Blight. Phytopathology 2006, 96, 425–431. [Google Scholar] [CrossRef] [Green Version]
- McElrone, A.J.; Reid, C.D.; Hoye, K.A.; Hart, E.; Jackson, R.B. Elevated CO2 reduces disease incidence and severity of a red maple fungal pathogen via changes in host physiology and leaf chemistry. Glob. Chang. Biol. 2005, 11, 1828–1836. [Google Scholar] [CrossRef]
- Hibberd, J.M.; Whitbread, R.; Farrar, J.F. Effect of 700 μmol mol−1 CO2 and infection with powdery mildew on the growth and carbon partitioning of barley. New Phytol. 1996, 134, 309–315. [Google Scholar] [CrossRef]
- Carter, T.R.; Saarikko, R.A. Estimating regional crop potential in Finland under a changing climate. Agric. For. Meteorol. 1996, 79, 301–313. [Google Scholar] [CrossRef]
- Chitarra, W.; Siciliano, I.; Ferrocino, I.; Gullino, M.L.; Garibaldi, A. Effect of elevated atmospheric CO2 and temperature on the disease severity of rocket plants caused by Fusarium wilt under phytotron conditions. PLoS ONE 2015, 10, e0140769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelsey, P.; Cooke, D.E.L.; Lynott, J.S.; Lees, A.K. Crop connectivity under climate change: Future environmental and geographic risks of potato late blight in Scotland. Glob. Chang. Biol. 2016, 22, 3724–3738. [Google Scholar] [CrossRef]
- Bonde, M.R.; Nester, S.E.; Berner, D.K. Effects of frequency of “extreme” temperature highs on development of soybean rust. Phytopathology 2013, 103, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Robson, J.D.; Wright, M.G.; Almeida, R.P.P. Biology of Pentalonia nigronervosa (Hemiptera, Aphididae) on Banana Using Different Rearing Methods. Environ. Entomol. 2007, 36, 46–52. [Google Scholar] [CrossRef]
- Anhalt, M.D.; Almeida, R.P.P. Effect of temperature, vector life stage, and plant access period on transmission of banana bunchy top virus to banana. Phytopathology 2008, 98, 743–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.J.; Pirie, E.J.; Evans, N.; Delourme, R.; King, G.J.; Fitt, B.D.L. Quantitative resistance to symptomless growth of Leptosphaeria maculans (phoma stem canker) in Brassica napus (oilseed rape). Plant Pathol. 2009, 58, 314–323. [Google Scholar] [CrossRef]
- Lukas, S.; Abbas, S.J.; Kössler, P.; Karlovsky, P.; Potthoff, M.; Joergensen, R.G. Fungal plant pathogens on inoculated maize leaves in a simulated soil warming experiment. Appl. Soil Ecol. 2018, 124, 75–82. [Google Scholar] [CrossRef]
- Manici, L.M.; Bregaglio, S.; Fumagalli, D.; Donatelli, M. Modelling soil borne fungal pathogens of arable crops under climate change. Int. J. Biometeorol. 2014, 58, 2071–2083. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.P.; Fawcett, L.; Anthony, S.G.; Young, C. A model for sclerotinia sclerotiorum infection and disease development in lettuce, based on the effects of temperature, relative humidity and ascospore density. PLoS ONE 2014, 9, e94049. [Google Scholar] [CrossRef] [PubMed]
- Bidzinski, P.; Ballini, E.; Ducasse, A.; Michel, C.; Zuluaga, P.; Genga, A.; Chiozzotto, R.; Morel, J.-B. Transcriptional Basis of Drought-Induced Susceptibility to the Rice Blast Fungus Magnaporthe oryzae. Front. Plant Sci. 2016, 7, 1558. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.M.D.; Toyota, K. Effect of Moisture Conditions and Pre-Incubation at Low Temperature on Bacterial Wilt of Tomato Caused by Ralstonia solanacearum. Microbes Environ. 2004, 19, 244–247. [Google Scholar] [CrossRef] [Green Version]
- Johansen, T.J.; Dees, M.W.; Hermansen, A. High soil moisture reduces common scab caused by Streptomyces turgidiscabies and Streptomyces europaeiscabiei in potato. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2015, 65, 193–198. [Google Scholar] [CrossRef]
- Homet, P.; González, M.; Matías, L.; Godoy, O.; Pérez-Ramos, I.M.; García, L.V.; Gómez-Aparicio, L. Exploring interactive effects of climate change and exotic pathogens on Quercus suber performance: Damage caused by Phytophthora cinnamomi varies across contrasting scenarios of soil moisture. Agric. For. Meteorol. 2019, 276–277, 107605. [Google Scholar] [CrossRef]
- Zhou, Y.; Van Leeuwen, S.K.; Pieterse, C.M.J.; Bakker, P.A.H.M.; Van Wees, S.C.M. Effect of atmospheric CO2 on plant defense against leaf and root pathogens of Arabidopsis. Eur. J. Plant Pathol. 2019, 154, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Chang. 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant–Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [Green Version]
- Zayan, S.A. Impact of climate change on plant diseases and IPM strategies. Arab J. Plant Prot. 2018, 36, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Zhao, M.; Shen, Z.; Deng, X.; Ou, Y.; Tao, C.; Liu, H.; Li, R.; Shen, Q. Biofertilizer application triggered microbial assembly in microaggregates associated with tomato bacterial wilt suppression. Biol. Fertil. Soils 2020, 56, 551–563. [Google Scholar] [CrossRef]
- Kerry, B.R.; Crump, D.H.; Mullen, L.A. Natural control of the cereal cyst nematode, Heterodera avenae Woll., by soil fungi at three sites. Crop Prot. 1982, 1, 99–109. [Google Scholar] [CrossRef]
- El-Tarabily, K.A. Suppression of Rhizoctonia solani diseases of sugar beet by antagonistic and plant growth-promoting yeasts. J. Appl. Microbiol. 2004, 96, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, S.; Saleena, L.M.; Vasudevan, P.; Nair, S. Biological suppression of rice diseases by Pseudomonas spp. under saline soil conditions. Plant Soil 2003, 251, 73–82. [Google Scholar] [CrossRef]
- Li, X.; de Boer, W.; Zhang, Y.; Ding, C.; Zhang, T.; Wang, X. Suppression of soil-borne Fusarium pathogens of peanut by intercropping with the medicinal herb Atractylodes lancea. Soil Biol. Biochem. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Mixtures of plant disease suppressive bacteria enhance biological control of multiple tomato pathogens. Biol. Fertil. Soils 2002, 36, 260–268. [Google Scholar] [CrossRef]
- Latz, E.; Eisenhauer, N.; Rall, B.C.; Scheu, S.; Jousset, A. Unravelling Linkages between Plant Community Composition and the Pathogen-Suppressive Potential of Soils. Sci. Rep. 2016, 6, 23584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Huang, X.; Shi, D.; Sun, F.; Lu, H.; Liu, J.; Wu, W. Efficacy of sludge and manure compost amendments against Fusarium wilt of cucumber. Environ. Sci. Pollut. Res. 2012, 19, 3895–3905. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Z.V.; Morandi, M.A.B.; Bettiol, W. Induction of suppressiveness to Fusarium wilt of chrysanthemum with composted sewage sludge. Trop. Plant Pathol. 2013, 38, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Cotxarrera, L.; Trillas-Gay, M.I.; Steinberg, C.; Alabouvette, C. Use of sewage sludge compost and Trichoderma asperellum isolates to suppress Fusarium wilt of tomato. Soil Biol. Biochem. 2002, 34, 467–476. [Google Scholar] [CrossRef]
- Li, B.; Xie, G.; Soad, A.; Coosemans, J. Suppression of Meloidogyne javanica by antagonistic and plant. J. Zhejiang Univ. Sci. 2005, 6, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Löbmann, M.T.; Vetukuri, R.R.; de Zinger, L.; Alsanius, B.W.; Grenville-Briggs, L.J.; Walter, A.J. The occurrence of pathogen suppressive soils in Sweden in relation to soil biota, soil properties, and farming practices. Appl. Soil Ecol. 2016, 107, 57–65. [Google Scholar] [CrossRef]
- Amir, H.; Alabouvette, C. Involvement of soil abiotic factors in the mechanisms of soil suppressiveness to fusarium wilts. Soil Biol. Biochem. 1993, 25, 157–164. [Google Scholar] [CrossRef]
- Cha, J.; Han, S.; Hong, H.; Cho, H.; Kim, D.; Kwon, Y.; Kwon, S.; Crüsemann, M.; Lee, Y.B.; Kim, J.F.; et al. Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 2016, 10, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, N.J.A.; Backhouse, D.B.; Burgess, L.W.C. Enhanced suppression of Gaeumannomyces graminis var. tritici by retention of residues in a cereal cropping system. Australas. Plant Pathol. 2006, 35, 43–48. [Google Scholar] [CrossRef]
- Shiomi, Y.; Nishiyama, M.; Onizuka, T.; Marumoto, T. Comparison of bacterial community structures in the rhizoplane of tomato plants grown in soils suppressive and conducive towards bacterial wilt. Appl. Environ. Microbiol. 1999, 65, 3996–4001. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, N.; Tiedje, J.M.; Quensen, J.F.; Meng, Q.; Hao, J.J. Microbial communities associated with potato common Scab-Suppressive soil determined by pyrosequencing analyses. Plant Dis. 2012, 96, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Ghini, R.; Morandi, M.A.B. Biotic and abiotic factors associated with soil suppressiveness to Rhizodonia solani. Sci. Agric. 2006, 63, 153–160. [Google Scholar] [CrossRef]
- Cook, R.J. Making greater use of introduced microorganisms for biological control of plant pathogens. Annu. Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef]
- Duran, P.; Mora, M.D.L.L. Plant–Soil–Microorganism Interaction Involved in Natural Suppression of Take-All Disease; Kaushal, M., Prasad, R., Eds.; Springer Nature: Singapore, 2021; ISBN 9781461479789. [Google Scholar]
- Rovira, A.D.; Campbell, R. A scanning electron microscope study of interactions between micro-organisms and Gaeumannomyces graminis (Syn. Ophiobolus graminis) on wheat roots. Microb. Ecol. 1975, 2, 177–185. [Google Scholar] [CrossRef]
- Cook, R.J.; Rovira, A.D. The role of bacteria in the biological control of Gaeumannomyces graminis by suppressive soils. Soil Biol. Biochem. 1976, 8, 269–273. [Google Scholar] [CrossRef]
- Ayyadurai, N.; Ravindra Naik, P.; Sakthivel, N. Functional characterization of antagonistic fluorescent pseudomonads associated with rhizospheric soil of rice (Oryza sativa L.). J. Microbiol. Biotechnol. 2007, 17, 919–927. [Google Scholar]
- Kilani-Feki, O.; Khiari, O.; Culioli, G.; Ortalo-Magné, A.; Zouari, N.; Blache, Y.; Jaoua, S. Antifungal activities of an endophytic Pseudomonas fluorescens strain Pf1TZ harbouring genes from pyoluteorin and phenazine clusters. Biotechnol. Lett. 2010, 32, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Validov, S.; Mavrodi, O.; De La Fuente, L.; Boronin, A.; Weller, D.; Thomashow, L.; Mavrodi, D. Antagonistic activity among 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. FEMS Microbiol. Lett. 2005, 242, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamali, F.; Sharifi-Tehrani, A.; Lutz, M.P.; Maurhofer, M. Influence of host plant genotype, presence of a pathogen, and coinoculation with pseudomonas fluorescens strains on the rhizosphere expression of hydrogen cyanide- and 2,4-diacetylphloroglucinol biosynthetic genes in P. fluorescens biocontrol strain CHA0. Microb. Ecol. 2009, 57, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.-S.; Bakker, P.A.H.M.; Glandorf, D.C.M.; Rice, J.T.; Paulitz, T.C.; Weller, D.M. Diversity, virulence, and 2,4-diacetylphloroglucinol sensitivity of Gaeumannomyces graminis var. tritici isolates from Washington state. Phytopathology 2009, 99, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Cordovez, V.; Carrion, V.J.; Etalo, D.W.; Mumm, R.; Zhu, H. Diversity and functions of volatile organic compounds produced by Streptomyces from a disease-suppressive soil. Front. Microbiol. 2015, 6, 1081. [Google Scholar] [CrossRef] [Green Version]
- Carrión, V.J.; Cordovez, V.; Tyc, O.; Etalo, D.W.; de Bruijn, I.; de Jager, V.C.L.; Medema, M.H.; Eberl, L.; Raaijmakers, J.M. Involvement of Burkholderiaceae and sulfurous volatiles in disease-suppressive soils. ISME J. 2018, 12, 2307–2321. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-R.; Jeon, C.-W.; Shin, J.-H.; Weller, D.M.; Thomashow, L.; Kwak, Y.-S. Function and Distribution of a Lantipeptide in Strawberry Fusarium wilt Disease–Suppressive Soils. Mol. Plant-Microbe Interact. 2018, 32, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Mazzola, M. Soil immune responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Wang, L.; Zhu, K.; Hou, S.; Chen, L.; Mi, D.; Gui, Y.; Qi, Y.; Jiang, C.; Guo, J.H. Plant root exudates are involved in Bacillus cereusAR156 mediated biocontrol against Ralstonia solanacearum. Front. Microbiol. 2019, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; De Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Ráez, J.A.; Pozo, M.J.; García-garrido, J.M. Strigolactones: A cry for help in the rhizosphere. Botany 2011, 89, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Zitzelsberger, C.; Buchbauer, G. Essential Oils as “A Cry for Help”. A Review. Nat. Prod. Commun. 2015, 10, 1127–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuberre, C.; Plancot, B.; Driouich, A.; Moore, J.P.; Bardor, M.; Gügi, B.; Vicré, M. Plant immunity is compartmentalized and specialized in roots. Front. Plant Sci. 2018, 871, 1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcel, S.; Paszkowski, U.; Sawers, R.; Oakeley, E.; Angliker, H. Tissue-adapted invasion strategies of the rice blast fungus Magnaporthe oryzae. Plant Cell 2010, 22, 3177–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Lemanceau, P.; Blouin, M.; Muller, D.; Moënne-Loccoz, Y. Let the Core Microbiota Be Functional. Trends Plant Sci. 2017, 22, 583–595. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Trivedi, C.; Hamonts, K.; Anderson, I.C.; Singh, B.K. Keystone microbial taxa regulate the invasion of a fungal pathogen in agro-ecosystems. Soil Biol. Biochem. 2017, 111, 10–14. [Google Scholar] [CrossRef]
- Chapelle, E.; Mendes, R.; Bakker, P.A.H.; Raaijmakers, J.M. Fungal invasion of the rhizosphere microbiome. ISME J. 2016, 10, 265–268. [Google Scholar] [CrossRef] [Green Version]
- van der Voort, M.; Kempenaar, M.; van Driel, M.; Raaijmakers, J.M.; Mendes, R. Impact of soil heat on reassembly of bacterial communities in the rhizosphere microbiome and plant disease suppression. Ecol. Lett. 2016, 19, 375–382. [Google Scholar] [CrossRef]
- Han, Y.; Xu, L.; Liu, L.; Yi, M. Illumina sequencing reveals a rhizosphere bacterial community associated with foxtail millet smut disease suppression. Plant Soil 2017, 410, 411–421. [Google Scholar] [CrossRef]
- Acuña, J.J.; Marileo, L.G.; Araya, M.A.; Rilling, J.I.; Larama, G.A.; Mora, M.L.; Epstein, S.; Jorquera, M.A. In Situ Cultivation Approach to Increase the Culturable Bacterial Diversity in the Rhizobiome of Plants. J. Soil Sci. Plant Nutr. 2020, 20, 1411–1426. [Google Scholar] [CrossRef]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J. Soil Sci. Plant Nutr. 2021, 21, 49–68. [Google Scholar] [CrossRef]
- Acuña, J.J.; Jorquera, M.A.; Barra, P.J.; Crowley, D.E.; de la Luz Mora, M. Selenobacteria selected from the rhizosphere as a potential tool for Se biofortification of wheat crops. Biol. Fertil. Soils 2013, 49, 175–185. [Google Scholar] [CrossRef]
- Barra, P.J.; Pontigo, S.; Delgado, M.; Parra–Almuna, L.; Duran, P.; Valentine, A.J.; Jorquera, M.A.; de la Luz Mora, M. Phosphobacteria inoculation enhances the benefit of P–fertilization on Lolium perenne in soils contrasting in P–availability. Soil Biol. Biochem. 2019, 136, 107516. [Google Scholar] [CrossRef]
- Barra, P.J.; Viscardi, S.; Jorquera, M.A.; Duran, P.A.; Valentine, A.J.; de la Luz Mora, M. Understanding the Strategies to Overcome Phosphorus–Deficiency and Aluminum–Toxicity by Ryegrass Endophytic and Rhizosphere Phosphobacteria. Front. Microbiol. 2018, 9, 1155. [Google Scholar] [CrossRef]
- Barra, P.J.; Inostroza, N.G.; Mora, M.L.; Crowley, D.E.; Jorquera, M.A. Bacterial consortia inoculation mitigates the water shortage and salt stress in an avocado (Persea americana Mill.) nursery. Appl. Soil Ecol. 2017, 111, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Barra, P.J.; Inostroza, N.G.; Acuña, J.J.; Mora, M.L.; Crowley, D.E.; Jorquera, M.A. Formulation of bacterial consortia from avocado (Persea americana Mill.) and their effect on growth, biomass and superoxide dismutase activity of wheat seedlings under salt stress. Appl. Soil Ecol. 2016, 102, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New frontiers in agriculture productivity: Optimised microbial inoculants and in situ microbiome engineering. Biotechnol. Adv. 2019, 37, 107371. [Google Scholar] [CrossRef] [PubMed]
- Inostroza, N.G.; Barra, P.J.; Wick, L.Y.; Mora, M.L.; Jorquera, M.A. Effect of rhizobacterial consortia from undisturbed arid- and agro-ecosystems on wheat growth under different conditions. Lett. Appl. Microbiol. 2017, 64, 158–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Gong, T.; Wang, J.; Li, G.; Liu, Y.; Zhen, J.; Ning, M.; Yue, D.; Du, Z.; Chen, G. Effects of Compound Microbial Fertilizer on Soil Characteristics and Yield of Wheat (Triticum aestivum L.). J. Soil Sci. Plant Nutr. 2020, 20, 2740–2748. [Google Scholar] [CrossRef]
- Moretti, L.G.; Crusciol, C.A.C.; Bossolani, J.W.; Momesso, L.; Garcia, A.; Kuramae, E.E.; Hungria, M. Bacterial Consortium and Microbial Metabolites Increase Grain Quality and Soybean Yield. J. Soil Sci. Plant Nutr. 2020, 20, 1923–1934. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.J.; Jorquera, M.A.; Azcón, R.; Paredes, C.; Rengel, Z.; de la Luz Mora, M. Endophytic bacteria from selenium-supplemented wheat plants could be useful for plant-growth promotion, biofortification and Gaeumannomyces graminis biocontrol in wheat production. Biol. Fertil. Soils 2014, 50, 983–990. [Google Scholar] [CrossRef]
- Pineda, A.; Kaplan, I.; Bezemer, T.M. Steering Soil Microbiomes to Suppress Aboveground Insect Pests. Trends Plant Sci. 2017, 22, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Mattupalli, C.; Eversole, K.; Leach, J.E. Enabling sustainable agriculture through understanding and enhancement of microbiomes. New Phytol. 2021, 230, 2129–2147. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, M.; Droby, S. The postharvest microbiome: The other half of sustainability. Biol. Control 2019, 137, 104025. [Google Scholar] [CrossRef]
- Koskella, B.; Bergelson, J. The study of host—microbiome (co) evolution across levels of selection. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190604. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Herren, C.M.; McMahon, K.D. Keystone taxa predict compositional change in microbial communities. Environ. Microbiol. 2018, 20, 2207–2217. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Wu, X.; Yang, J.; Ruan, H.; Wang, S.; Yang, Y.; Naeem, I.; Wang, L.; Liu, L.; Wang, D. The diversity and co-occurrence network of soil bacterial and fungal communities and their implications for a new indicator of grassland degradation. Ecol. Indic. 2021, 129, 107989. [Google Scholar] [CrossRef]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Duran, P.; Pearce, D.A. Niche Differentiation in the Composition, Predicted Function, and Co-occurrence Networks in Bacterial Communities Associated With Antarctic Vascular Plants. Front. Microbiol. 2020, 11, 1036. [Google Scholar] [CrossRef] [PubMed]
- Jeewani, P.H.; Gunina, A.; Tao, L.; Zhu, Z.; Kuzyakov, Y.; Van Zwieten, L.; Guggenberger, G.; Shen, C.; Yu, G.; Singh, B.P.; et al. Rusty sink of rhizodeposits and associated keystone microbiomes. Soil Biol. Biochem. 2020, 147, 107840. [Google Scholar] [CrossRef]
- Bao, Y.; Dolfing, J.; Guo, Z.; Chen, R.; Wu, M.; Li, Z.; Lin, X.; Feng, Y. Important ecophysiological roles of non-dominant Actinobacteria in plant residue decomposition, especially in less fertile soils. Microbiome 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Ishimoto, C.K.; Aono, A.H.; Nagai, J.S.; Sousa, H.; Miranda, A.R.L.; Melo, V.M.M.; Mendes, L.W.; Araujo, F.F.; de Melo, W.J.; Kuroshu, R.M.; et al. Microbial co-occurrence network and its key microorganisms in soil with permanent application of composted tannery sludge. Sci. Total Environ. 2021, 789, 147945. [Google Scholar] [CrossRef]
- Zheng, Y.; Han, X.; Zhao, D.; Wei, K.; Yuan, Y.; Li, Y.; Liu, M.; Zhang, C.S. Exploring Biocontrol Agents From Microbial Keystone Taxa Associated to Suppressive Soil: A New Attempt for a Biocontrol Strategy. Front. Plant Sci. 2021, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Dastogeer, K.M.G.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant microbiome—An account of the factors that shape community composition and diversity. Curr. Plant Biol. 2020, 23, 100161. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Hartmann, M. Networking in the Plant Microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Müller, D.B. Establishing Causality: Opportunities of Synthetic Communities for Plant Microbiome Research. Cell Host Microbe 2017, 22, 142–155. [Google Scholar] [CrossRef]
- Panke-Buisse, K.; Poole, A.C.; Goodrich, J.K.; Ley, R.E.; Kao-Kniffin, J. Selection on soil microbiomes reveals reproducible impacts on plant function. ISME J. 2015, 9, 980–989. [Google Scholar] [CrossRef]
- Jochum, M.D.; McWilliams, K.L.; Pierson, E.A.; Jo, Y.K. Host-mediated microbiome engineering (HMME) of drought tolerance in the wheat rhizosphere. PLoS ONE 2019, 14, e0225933. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.; Choudhary, M.; Gupta, S.; Dhar, M.K. Engineering Host Microbiome for Crop Improvement and Sustainable Agriculture. Front. Microbiol. 2021, 12, 1125. [Google Scholar] [CrossRef] [PubMed]
- French, E.; Kaplan, I.; Iyer-Pascuzzi, A.; Nakatsu, C.H.; Enders, L. Emerging strategies for precision microbiome management in diverse agroecosystems. Nat. Plants 2021, 7, 256–267. [Google Scholar] [CrossRef]
- Rodriguez, R.; Durán, P. Natural Holobiome Engineering by Using Native Extreme Microbiome to Counteract the Climate Change Effects. Front. Bioeng. Biotechnol. 2020, 8, 568. [Google Scholar] [CrossRef]
- Niu, B.; Paulson, J.N.; Zheng, X.; Kolter, R. Simplified and representative bacterial community of maize roots. Proc. Natl. Acad. Sci. USA 2017, 114, E2450–E2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.O.; Tjards, S.; Rischling, J.; Nguyen, D.T.; Cressler, C.E. Multiple generations of antibiotic exposure and isolation influence host fitness and the microbiome in a model zooplankton species. bioRxiv 2021, 68502. [Google Scholar] [CrossRef]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.V.; Bhattarai, S.; Rojas-Correa, M.; Purkayastha, A.; Holler, D.; Da Qu, M.; Mitchell, W.G.; Yang, J.; Fountain, S.; Zeamer, A.; et al. The intestinal and oral microbiomes are robust predictors of covid-19 severity the main predictor of covid-19-related fatality. medRxiv 2021. [Google Scholar] [CrossRef]
- Bucci, V.; Xavier, J.B. Towards predictive models of the human gut microbiome. J. Mol. Biol. 2014, 426, 3907–3916. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate Variable | Host | Incidence | Pathogen | Location | Reference |
|---|---|---|---|---|---|
| >CO2 | Wheat | increase increase | Fusarium sp. Septoria tritici | In vitro (Ireland) In vitro (Ireland) | [55] |
| Rice | increase | Magnaporthe oryzae | Japan | [56] | |
| Soybean | decrease | Peronospora manshurica | USA | [49] | |
| Soybean | increase | Septoria glycines | USA | ||
| neutral | Fusarium virguliforme | USA | |||
| Duke forest | decrease | Phyllosticta minima Acer rubrum | USA | [57] | |
| Barley | decrease | Erysiphe graminis | In vitro (UK) | [58] | |
| >CO2 + >T° | Wheat, barley, oat, potato, maize | increase | Globodera rostochiensis Meloidogyne chitwoodi Phytophthora infestans | Finland | [59] |
| Eruca sativa | increase | Fusarium oxysporum | In vitro (Italy) | [60] | |
| >T° | Potato | increase | Phytophtora infestans (first half of plant growing season) | UK | [61] |
| decrease | Phytophtora infestans (second half of plant growing season) | UK | |||
| Wheat | increase | Puccinia striformis | USA, Mexico, Denmark, Eritrea | [53] | |
| Soybean | increase | Phakopsora pachyrhizi | In vitro (USA) | [62] | |
| Banana bunchy top virus (BBTV) | increase | Pentalotia nigronervosa | In vitro (USA) | [63,64] | |
| Brasica napus | increase | Leptosphaeria maculans | France | [65] | |
| Maize | neutral | Fusarium culmorum | Germany | [66] | |
| increase | Fusarium oxysporum | ||||
| decrease | Rhizoctonia solani | ||||
| Cereals | increase | Fusarium nivale | Italy | [67] | |
| Wheat, barley | increase | Bipolaris sorokiniana | |||
| Several early crops | increase | Pythium ultimum | |||
| Horticultural crops | increase | Sclerotinia minor | |||
| Sunflower, sorghum, maize, cotton, soybean, etc. | increase | Macrophomina phaseolina | |||
| Lettuce | increase | Sclerotinia sclerotiorum | In vitro (UK) | [68] | |
| <Wa | Rice | increase | Magnaporthe oryzae | France | [69] |
| Tomato | increase | Ralstonia solanaceum | In vitro (Japan) | [70] | |
| Potato | decrease | Streptomyces turgidiscabies Streptomyces europaeiscabiei | In vitro (Norway) | [71] | |
| Soybean | increase | Peronospora manshurica | USA | [49] | |
| increase | Septoria glycines | ||||
| increase | Fusarium virguliforme |
| Pathogen | Country/Source Soil | Plant | Suppresser | Reference |
|---|---|---|---|---|
| Fungi | ||||
| Rhizoctonia solani Fusarium sp. | Brazil/pasture, fallow ground, forest | Common bean | Abiotic (hydrolysis of fluorescein diacetate, CO2) Biotic (total microbial activity) | [78] |
| Rhizoctonia solani | Egypt | Sugar beet | Plant growth promoting (PGP yeast), Candida valida, Rhodotorula glutinis, Trichosporon asahii) | [79] |
| Rhizoctonia solani | India | Rice | Pseudomonas spp. | [80] |
| Rhizoctonia solani Pythium aphanidermatum, Fusarium oxysporum | Belgium | Mung bean | PGP rhizobacteria (Brevibacillus brevis, Bacillus subtilis) | [81] |
| Rhizoctonia solani Macrophomina phaseolina Fusarium solani | Pakistan | Tomatoes | PGP rhizobacteria (Pseudomonas fluorescens, Pseudomonas aeruginosa, Bradyrhizobium japonicum) | [82] |
| Rhizoctonia solani | Germany | Sugar beet | Abiotic (pH) Biotic (Actinomyces, Bacillus, Pseudomona) | [83] |
| Rhizoctonia solani | Netherlands | Sugar beet | Proteobacteria, Firmicutes, Actinobacteria | [84] |
| Fusarium sp. | substrate | Cucumber | Sludge compost: sewage sludge (pig manure), sawdust, matured sludge compost | [85] |
| Fusarium sp. | substrate | Chrysanthemum | Composted sewage sludge into the Pinus bark-based substrate | [86] |
| Fusarium sp. | substrate | Tomatoes | Sewage sludge and yard wastes | [87] |
| Fusarium spp. | China | Peanut | Intercropping of peanut with Atractylodes lancea | [88] |
| Pythium ultimum | Sweden | Wheat | Permanent soil cover and a balanced nutrient | [89] |
| Fusarium oxysporum | Algeria | Palm groves | Soil abiotic factors (i.e., clay addition to sansy soil) | [90] |
| Fusarium oxysporum | Korea | Strawberry | Actinobacteria | [91] |
| Fusarium oxysporum | Brasil | Common bean | Pseudomonadaceae, bacillaceae, solibacteraceae and cytophagaceae | [19] |
| Fusarium solani | Pakistan | Tomatoes | PGP rhizobacteria (Pseudomonas fluorescens, Pseudomonas aeruginosa, Bradyrhizobium japonicum) | [82] |
| Gaeumannomyces graminis | Chile | Wheat | Soil microbiome | [15] |
| Gaeumannomyces graminis | Chile | Wheat | Endophytic microbiome | [17,22] |
| Gaeumannomyces graminis | Australia | Wheat | Stubble retention and reduced tillage | [92] |
| Bacteria | ||||
| Ralstonia solanacearum | Japan | Tomato | Soil bacteria | [93] |
| Xanthomonas oryzae | India | Rice | Pseudomonas spp. | [80] |
| Streptomyces spp. | USA | Potato | Lysobacter, acidobacteria | [94] |
| Nematode | ||||
| Heterodera avenae | UK | Oat | Verticillium chlamydosporium, Nematophthora gynophila | [95] |
| Meloidogyne javanica | Belgium | Mung bean | PGP rhizobacteria (Brevibacillus brevis, Bacillus subtilis) | [81] |
| Meloidogyne javanica | Pakistan | Tomatoes | PGP rhizobacteria (Pseudomonas fluorescens, Pseudomonas aeruginosa, Bradyrhizobium japonicum) | [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durán, P.; Tortella, G.; Sadowsky, M.J.; Viscardi, S.; Barra, P.J.; Mora, M.d.l.L. Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change. Biology 2021, 10, 865. https://doi.org/10.3390/biology10090865
Durán P, Tortella G, Sadowsky MJ, Viscardi S, Barra PJ, Mora MdlL. Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change. Biology. 2021; 10(9):865. https://doi.org/10.3390/biology10090865
Chicago/Turabian StyleDurán, Paola, Gonzalo Tortella, Michael J. Sadowsky, Sharon Viscardi, Patricio Javier Barra, and Maria de la Luz Mora. 2021. "Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change" Biology 10, no. 9: 865. https://doi.org/10.3390/biology10090865
APA StyleDurán, P., Tortella, G., Sadowsky, M. J., Viscardi, S., Barra, P. J., & Mora, M. d. l. L. (2021). Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change. Biology, 10(9), 865. https://doi.org/10.3390/biology10090865