
Isolation and Identification of Long Non-Coding RNAs in Exosomes Derived from the Serum of Colorectal Carcinoma Patients


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Serum Preparation and Storage
2.3. Vesicle Isolation from Serum Sample
2.4. Cell Culture and Vesicle Isolation from Culture Supernatants of SW480-7
2.5. Exosomal Protein Determination
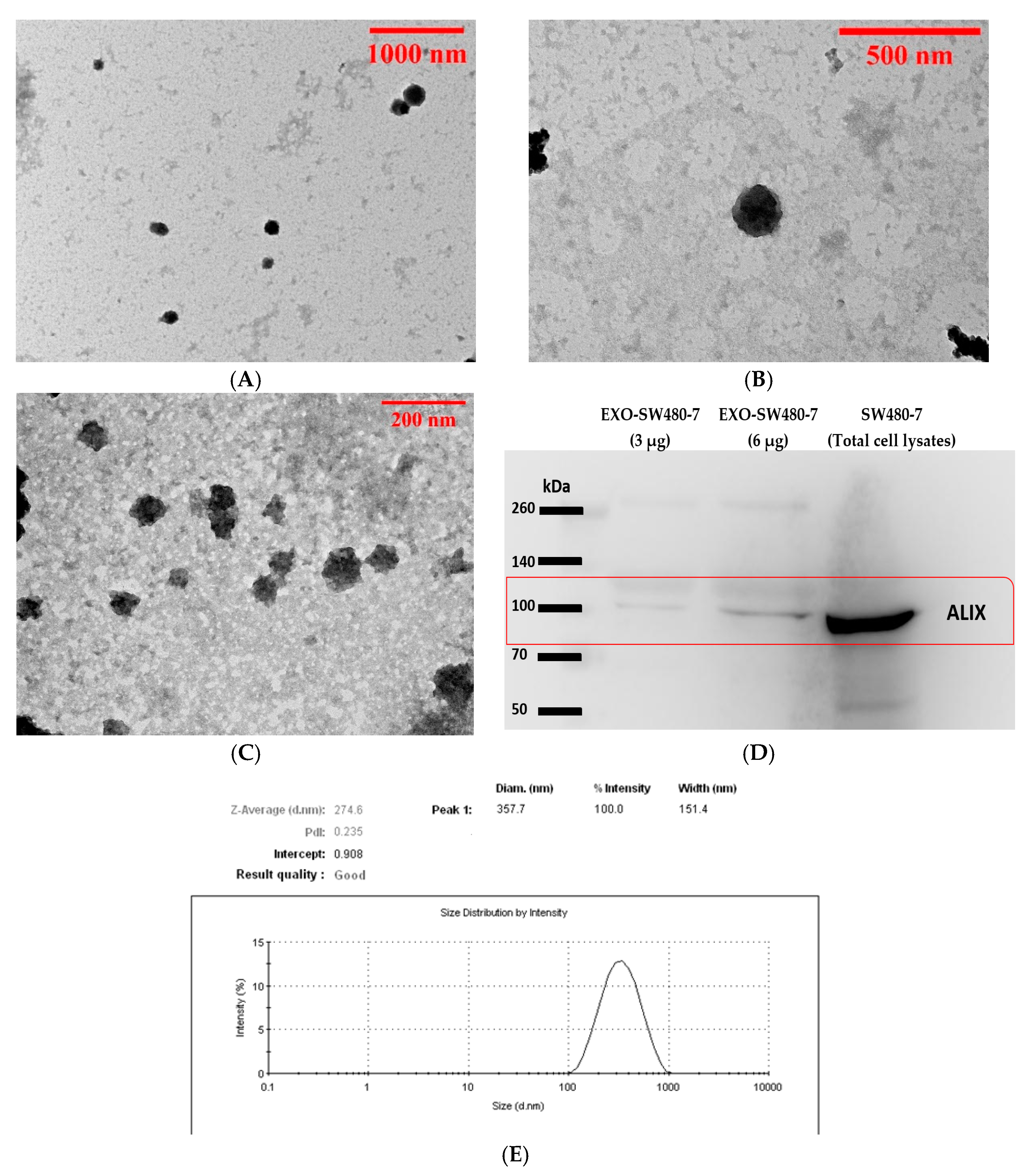
2.6. Detection of ALIX/PDCD6IP in Exosomes
2.7. Size Distribution Analysis
2.8. Determination of Exosome Morphology by Transmission Electron Microscopy (TEM)
2.9. RNA Extraction
2.10. Reverse Transcription (RT)
2.11. Detection of lncRNA in Exosomes by RT-PCR
2.12. Statistical Analysis
3. Results
3.1. Characterization of Exosomes Derived from Serum Sample and SW480-7 Cell Line
3.2. Clinical and Pathological Characteristics
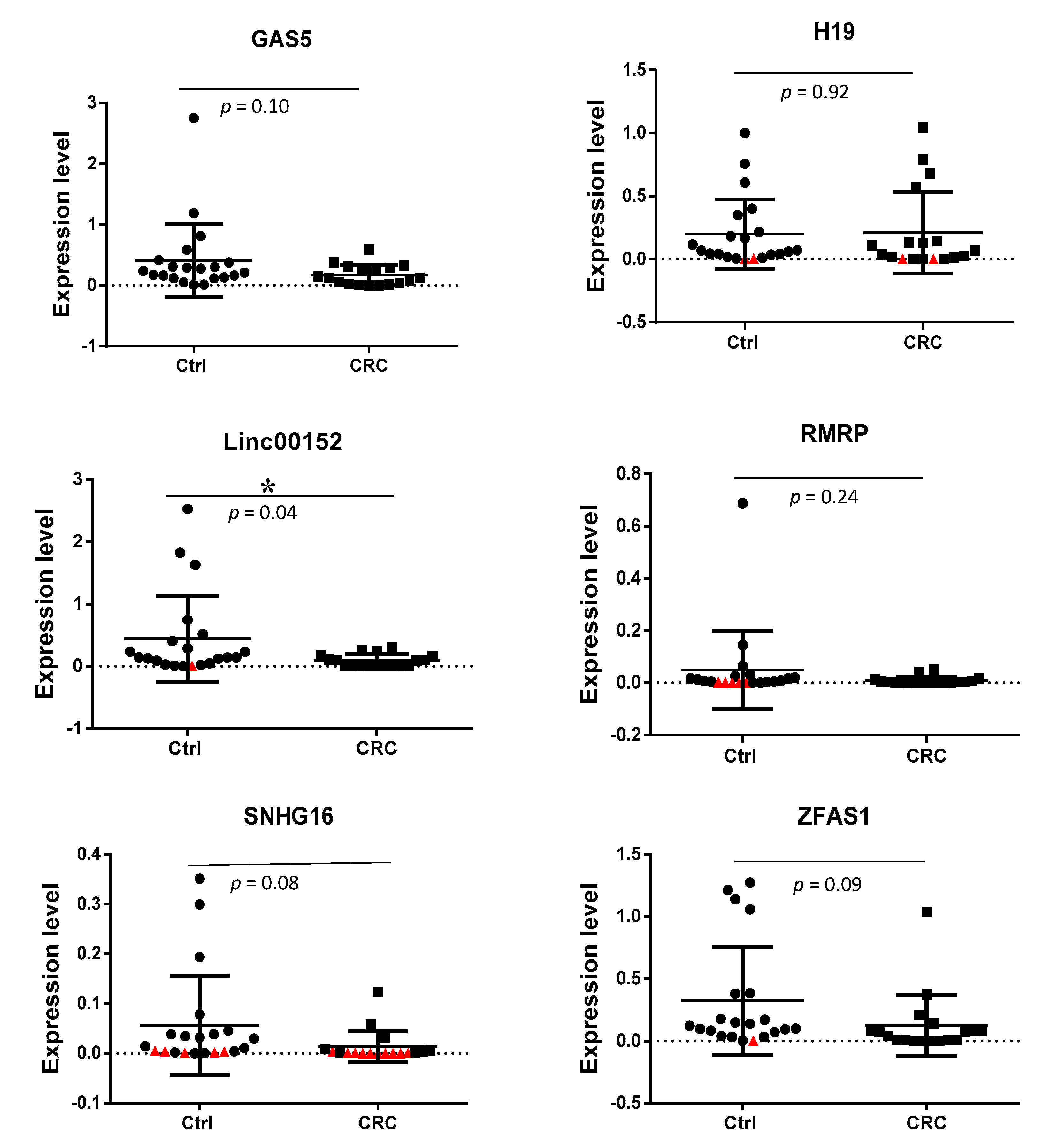
3.3. Detection of Exosomal lncRNAs Expression in CRC Patients and Non-Cancer Individuals
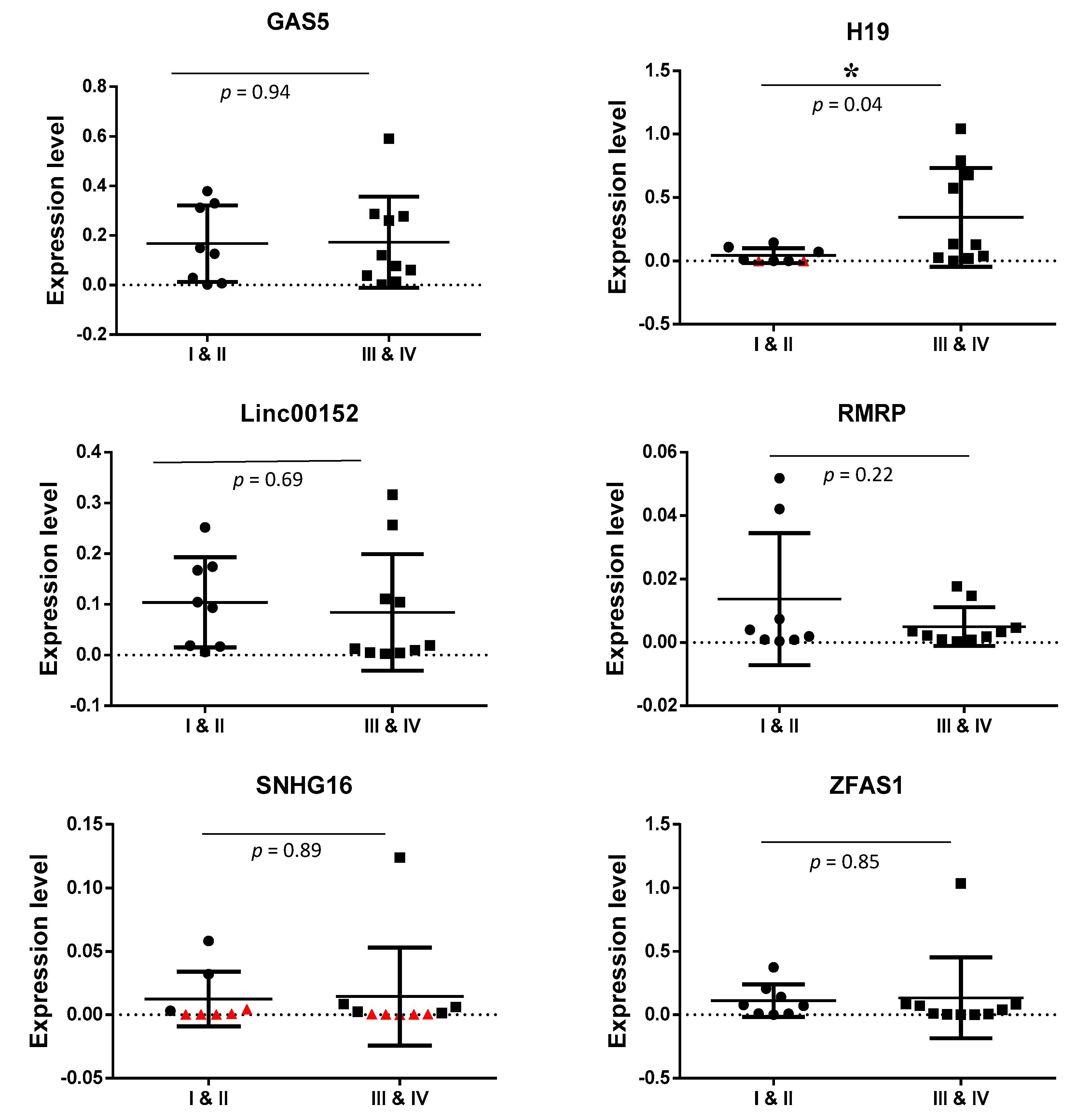
3.4. Detection of Exosomal lncRNAs Expression in Early and Advanced Stages of CRC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moriarity, A.; O’Sullivan, J.; Kennedy, J.; Mehigan, B.; McCormick, P. Current targeted therapies in the treatment of advanced colorectal cancer: A review. Ther. Adv. Med. Oncol. 2016, 8, 276–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Tang, B.; Xiao, Y.F.; Xie, R.; Li, B.S.; Dong, H.; Zhou, J.Y.; Yang, S.M. Long non-coding RNAs in colorectal cancer. Oncotarget 2016, 7, 5226–5239. [Google Scholar] [CrossRef] [Green Version]
- Saus, E.; Brunet-Vega, A.; Iraola-Guzman, S.; Pegueroles, C.; Gabaldon, T.; Pericay, C. Long Non-Coding RNAs As Potential Novel Prognostic Biomarkers in Colorectal Cancer. Front. Genet. 2016, 7, e54. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Wang, M.; Ma, N.; Xu, Y.; Jiang, Y.; Gao, X. Long noncoding RNAs: Novel players in colorectal cancer. Cancer Lett. 2015, 361, 13–21. [Google Scholar] [CrossRef]
- Ward, M.; McEwan, C.; Mills, J.D.; Janitz, M. Conservation and tissue-specific transcription patterns of long noncoding RNAs. J. Hum. Transcr. 2015, 1, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Raut, S.K.; Khullar, M. The Big Entity of New RNA World: Long Non-Coding RNAs in Microvascular Complications of Diabetes. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef]
- Balas, M.M.; Johnson, A.M. Exploring the mechanisms behind long noncoding RNAs and cancer. Non-Coding Rna Res. 2018, 3, 108–117. [Google Scholar] [CrossRef]
- Smolle, M.; Uranitsch, S.; Gerger, A.; Pichler, M.; Haybaeck, J. Current status of long non-coding RNAs in human cancer with specific focus on colorectal cancer. Int. J. Mol. Sci. 2014, 15, 13993–14013. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.C.; Chen, T.; Zhu, D.X.; Lv, S.X.; Qiu, J.J.; Xu, J.; Yuan, F.L.; Wei, Y. Downregulated long non-coding RNA CLMAT3 promotes the proliferation of colorectal cancer cells by targeting regulators of the cell cycle pathway. Oncotarget 2016, 7, 58931–58938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.D.; Liu, Z.J.; Zhang, E.; Kong, R.; Zhang, Z.H.; Guo, R.H. Decreased expression of long noncoding RNA MEG3 affects cell proliferation and predicts a poor prognosis in patients with colorectal cancer. Tumor Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 4851–4859. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Sawada, G.; Kurashige, J.; Uchi, R.; Matsumura, T.; Ueo, H.; Takano, Y.; Eguchi, H.; Sudo, T.; Sugimachi, K.; et al. Amplification of PVT-1 is involved in poor prognosis via apoptosis inhibition in colorectal cancers. Br. J. Cancer 2014, 110, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, P.; Xu, M.D.; Ni, S.J.; Shen, X.H.; Wei, P.; Huang, D.; Tan, C.; Sheng, W.Q.; Zhou, X.Y.; Du, X. Down-regulation of ncRAN, a long non-coding RNA, contributes to colorectal cancer cell migration and invasion and predicts poor overall survival for colorectal cancer patients. Mol. Carcinog. 2015, 54, 742–750. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Fernandes, A.R.; Baptista, P.V. Exosome in tumor microenvironment: Overview of the crosstalk between normal and cancer cells. Biomed. Res. Int. 2014, 2014, e179486. [Google Scholar] [CrossRef]
- Li, J.; Xue, W.; Lv, J.; Han, P.; Liu, Y.; Cui, B. Identification of potential long non-coding RNA biomarkers associated with the progression of colon cancer. Oncotarget 2017, 8, 75834–75843. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, A.; Yoshioka, Y.; Hirakawa, A.; Yamamoto, Y.; Ishikawa, M.; Ikeda, S.I.; Kato, T.; Niimi, K.; Kajiyama, H.; Kikkawa, F.; et al. A combination of circulating miRNAs for the early detection of ovarian cancer. Oncotarget 2017, 8, 89811–89823. [Google Scholar] [CrossRef] [Green Version]
- Farran, B.; Dyson, G.; Craig, D.; Dombkowski, A.; Beebe-Dimmer, J.L.; Powell, I.J.; Podgorski, I.; Heilbrun, L.; Bolton, S.; Bock, C.H. A study of circulating microRNAs identifies a new potential biomarker panel to distinguish aggressive prostate cancer. Carcinogenesis 2018, 39, 556–561. [Google Scholar] [CrossRef]
- Fang, Z.; Tang, J.; Bai, Y.; Lin, H.; You, H.; Jin, H.; Lin, L.; You, P.; Li, J.; Dai, Z.; et al. Plasma levels of microRNA-24, microRNA-320a, and microRNA-423-5p are potential biomarkers for colorectal carcinoma. J. Exp. Clin. Cancer Res. Cr 2015, 34, e86. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Raju, G.S.; Chang, D.W.; Lin, S.H.; Chen, Z.; Wu, X. Global and targeted circulating microRNA profiling of colorectal adenoma and colorectal cancer. Cancer 2018, 124, 785–796. [Google Scholar] [CrossRef]
- Zheng, G.; Du, L.; Yang, X.; Zhang, X.; Wang, L.; Yang, Y.; Li, J.; Wang, C. Serum microRNA panel as biomarkers for early diagnosis of colorectal adenocarcinoma. Br. J. Cancer 2014, 111, 1985–1992. [Google Scholar] [CrossRef] [Green Version]
- Sapre, N.; Selth, L.A. Circulating MicroRNAs as Biomarkers of Prostate Cancer: The State of Play. Prostate Cancer 2013, 2013, e539680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamam, R.; Hamam, D.; Alsaleh, K.A.; Kassem, M.; Zaher, W.; Alfayez, M.; Aldahmash, A.; Alajez, N.M. Circulating microRNAs in breast cancer: Novel diagnostic and prognostic biomarkers. Cell Death Dis. 2017, 8, e3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragomir, M.; Chen, B.; Calin, G.A. Exosomal lncRNAs as new players in cell-to-cell communication. Transl. Cancer Res. 2018, 7, S243–S252. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Wei, M.; Han, X.; Xu, T.; Cui, M. Advances in the study of exosomal lncRNAs in tumors and the selection of research methods. Biomed. Pharmacother. 2020, 123, e109716. [Google Scholar] [CrossRef] [PubMed]
- He, P.Y.; Yip, W.K.; Chai, B.L.; Chai, B.Y.; Jabar, M.F.; Dusa, N.; Mohtarrudin, N.; Seow, H.F. Inhibition of cell migration and invasion by miR-29a-3p in a colorectal cancer cell line through suppression of CDC42BPA mRNA expression. Oncol. Rep. 2017, 38, 3554–3566. [Google Scholar] [CrossRef] [Green Version]
- Enderle, D.; Spiel, A.; Coticchia, C.M.; Berghoff, E.; Mueller, R.; Schlumpberger, M.; Sprenger-Haussels, M.; Shaffer, J.M.; Lader, E.; Skog, J.; et al. Characterization of RNA from Exosomes and Other Extracellular Vesicles Isolated by a Novel Spin Column-Based Method. PLoS ONE 2015, 10, e0136133. [Google Scholar] [CrossRef] [Green Version]
- Tickner, J.A.; Urquhart, A.J.; Stephenson, S.A.; Richard, D.J.; O’Byrne, K.J. Functions and therapeutic roles of exosomes in cancer. Front. Oncol. 2014, 4, e127. [Google Scholar] [CrossRef] [Green Version]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Feng, W.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. Exosomes promote pre-metastatic niche formation in ovarian cancer. Mol. Cancer 2019, 18, e124. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Ji, X.; Liu, J.; Fan, D.; Zhou, Q.; Chen, C.; Wang, W.; Wang, G.; Wang, H.; Yuan, W.; et al. Effects of exosomes on pre-metastatic niche formation in tumors. Mol. Cancer 2019, 18, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, X.; Gao, S.; Jing, F.; Yang, Y.; Du, L.; Zheng, G.; Li, P.; Li, C.; Wang, C. Exosomal long noncoding RNA CRNDE-h as a novel serum-based biomarker for diagnosis and prognosis of colorectal cancer. Oncotarget 2016, 7, 85551–85563. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.T.; Huang, Y.Y.; Zheng, L.; Qin, S.H.; Xu, X.P.; An, T.X.; Xu, Y.; Wu, Y.S.; Hu, X.M.; Ping, B.H.; et al. Comparison of isolation methods of exosomes and exosomal RNA from cell culture medium and serum. Int. J. Mol. Med. 2017, 40, 834–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Yang, J.; Li, Q.; Xu, B.; Lian, Y.; Miao, L. LINC00152: A pivotal oncogenic long non-coding RNA in human cancers. Cell Prolif. 2017, 50. [Google Scholar] [CrossRef] [Green Version]
- Ou, C.; Sun, Z.; He, X.; Li, X.; Fan, S.; Zheng, X.; Peng, Q.; Li, G.; Li, X.; Ma, J. Targeting YAP1/LINC00152/FSCN1 Signaling Axis Prevents the Progression of Colorectal Cancer. Adv. Sci. (Weinh. Baden-Wurtt. Ger.) 2020, 7, e1901380. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.H.; Fu, J.; Zhang, Z.J.; Ge, C.C.; Yi, Y. LncRNA-LINC00152 down-regulated by miR-376c-3p restricts viability and promotes apoptosis of colorectal cancer cells. Am. J. Transl. Res. 2016, 8, 5286–5297. [Google Scholar]
- Delihas, N. Editorial on the Special Issue: Regulation by non-coding RNAs. Int. J. Mol. Sci. 2013, 14, 21960–21964. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Dai, Y.; Wang, F.; Hou, S. Differentially expressed long non-coding RNAs and the prognostic potential in colorectal cancer. Neoplasma 2016, 63, 977–983. [Google Scholar] [CrossRef]
- Deng, Q.; He, B.; Gao, T.; Pan, Y.; Sun, H.; Xu, Y.; Li, R.; Ying, H.; Wang, F.; Liu, X.; et al. Up-regulation of 91H promotes tumor metastasis and predicts poor prognosis for patients with colorectal cancer. PLoS ONE 2014, 9, e103022. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Meng, T.; Yang, X.H.; Sayim, P.; Lei, C.; Jin, B.; Ge, L.; Wang, H.J. Prognostic and predictive value of long non-coding RNA GAS5 and mircoRNA-221 in colorectal cancer and their effects on colorectal cancer cell proliferation, migration and invasion. Cancer Biomark. Sect. A Dis. Markers 2018, 22, 283–299. [Google Scholar] [CrossRef]
- Galamb, O.; Barták, B.K.; Kalmár, A.; Nagy, Z.B.; Szigeti, K.A.; Tulassay, Z.; Igaz, P.; Molnár, B. Diagnostic and prognostic potential of tissue and circulating long non-coding RNAs in colorectal tumors. World J. Gastroenterol. 2019, 25, 5026–5048. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Dang, W.; Zhang, S.; Yue, W.; Yang, L.; Zhai, X.; Yan, Q.; Lu, J. The role of exosomal noncoding RNAs in cancer. Mol. Cancer 2019, 18, e37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.S.; Liu, Q.H.; Cheng, X.H.; Zhang, W.Y.; Jin, Y.C. The long noncoding RNA ZFAS1 facilitates bladder cancer tumorigenesis by sponging miR-329. Biomed. Pharmacother. 2018, 103, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Fan, C.; Liu, N.; Huang, K.; Fang, X.; Wang, K. Downregulation of the long non-coding RNA ZFAS1 is associated with cell proliferation, migration and invasion in breast cancer. Mol. Med. Rep. 2018, 17, 6405–6412. [Google Scholar] [CrossRef]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [CrossRef]
- Luo, P.; Liang, C.; Zhang, X.; Liu, X.; Wang, Y.; Wu, M.; Feng, X.; Tu, J. Identification of long non-coding RNA ZFAS1 as a novel biomarker for diagnosis of HCC. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zeng, K.; Xu, M.; Hu, X.; Liu, X.; Xu, T.; He, B.; Pan, Y.; Sun, H.; Wang, S. SP1-induced lncRNA-ZFAS1 contributes to colorectal cancer progression via the miR-150-5p/VEGFA axis. Cell Death Dis. 2018, 9, e982. [Google Scholar] [CrossRef]
- Thorenoor, N.; Faltejskova-Vychytilova, P.; Hombach, S.; Mlcochova, J.; Kretz, M.; Svoboda, M.; Slaby, O. Long non-coding RNA ZFAS1 interacts with CDK1 and is involved in p53-dependent cell cycle control and apoptosis in colorectal cancer. Oncotarget 2016, 7, 622–637. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xing, C. Upregulation of long noncoding RNA ZFAS1 predicts poor prognosis and prompts invasion and metastasis in colorectal cancer. Pathol. Res. Pract. 2016, 212, 690–695. [Google Scholar] [CrossRef]
- Christensen, L.L.; True, K.; Hamilton, M.P.; Nielsen, M.M.; Damas, N.D.; Damgaard, C.K.; Ongen, H.; Dermitzakis, E.; Bramsen, J.B.; Pedersen, J.S.; et al. SNHG16 is regulated by the Wnt pathway in colorectal cancer and affects genes involved in lipid metabolism. Mol. Oncol. 2016, 10, 1266–1282. [Google Scholar] [CrossRef]
- Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Scalia, M.; Magnano, A.; Caltabiano, R.; Barbagallo, D.; Biondi, A.; Cappellani, A.; et al. LncRNA UCA1, Upregulated in CRC Biopsies and Downregulated in Serum Exosomes, Controls mRNA Expression by RNA-RNA Interactions. Mol. Ther.-Nucleic Acids 2018, 12, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Ren, M.; Li, Y.; Song, X. LncRNA-RMRP Acts as an Oncogene in Lung Cancer. PLoS ONE 2016, 11, e0164845. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cycles | Duration | Temperature |
|---|---|---|
| 1 | 10 min | 95 °C |
| 40 | 15 s | 95 °C |
| 1 min | 60 °C |
| Clinico-Pathologic Parameters | Number of Patients (n = 18) |
|---|---|
| Sex | |
| Male | 9 |
| Female | 9 |
| Age (years) | |
| <60 | 8 |
| ≥60 | 10 |
| Tumor stage | |
| I | 1 |
| II | 7 |
| III | 7 |
| IV | 3 |
| Tumor location | |
| Colon | 6 |
| Sigmoid | 5 |
| Rectosigmoid | 2 |
| Rectum | 5 |
| Target Genes | lncRNA Expression Level | |||
|---|---|---|---|---|
| Non-Cancer (n = 21) | CRC (n = 18) | p-Value | Fold-Change | |
| GAS5 | 0.0708 | 0.2125 | 0.103 | −3.00 |
| H19 | 0.0255 | 0.0658 | 0.921 | −2.58 |
| LINC00152 | 0.0369 | 0.1190 | 0.040 * | −3.23 |
| RMRP | 0.0030 | 0.0070 | 0.246 | −2.33 |
| SNHG16 | 0.0015 | 0.0121 | 0.085 | −8.06 |
| ZFAS1 | 0.0230 | 0.0230 | 0.094 | −5.12 |
| Target Genes | lncRNAs Expression Level | |||
|---|---|---|---|---|
| Early Stages (n = 8) | Advanced Stages (n = 10) | p-Value | Fold Change | |
| GAS5 | 0.0762 | 0.0646 | 0.942 | 1.18 |
| H19 | 0.0957 | 0.0048 | 0.046 * | 19.59 |
| LINC00152 | 0.0256 | 0.0582 | 0.694 | −2.27 |
| RMRP | 0.0025 | 0.0036 | 0.225 | −1.44 |
| SNHG16 | 0.0012 | 0.0019 | 0.899 | 1.62 |
| ZFAS1 | 0.0171 | 0.0334 | 0.855 | −1.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, C.T.; Azwar, S.; Yip, W.K.; Zahari Sham, S.Y.; Faisal Jabar, M.; Sahak, N.H.; Mohtarrudin, N.; Seow, H.F. Isolation and Identification of Long Non-Coding RNAs in Exosomes Derived from the Serum of Colorectal Carcinoma Patients. Biology 2021, 10, 918. https://doi.org/10.3390/biology10090918
Ng CT, Azwar S, Yip WK, Zahari Sham SY, Faisal Jabar M, Sahak NH, Mohtarrudin N, Seow HF. Isolation and Identification of Long Non-Coding RNAs in Exosomes Derived from the Serum of Colorectal Carcinoma Patients. Biology. 2021; 10(9):918. https://doi.org/10.3390/biology10090918
Chicago/Turabian StyleNg, Chin Tat, Shamin Azwar, Wai Kien Yip, Siti Yazmin Zahari Sham, Mohd Faisal Jabar, Norren Haneezah Sahak, Norhafizah Mohtarrudin, and Heng Fong Seow. 2021. "Isolation and Identification of Long Non-Coding RNAs in Exosomes Derived from the Serum of Colorectal Carcinoma Patients" Biology 10, no. 9: 918. https://doi.org/10.3390/biology10090918
APA StyleNg, C. T., Azwar, S., Yip, W. K., Zahari Sham, S. Y., Faisal Jabar, M., Sahak, N. H., Mohtarrudin, N., & Seow, H. F. (2021). Isolation and Identification of Long Non-Coding RNAs in Exosomes Derived from the Serum of Colorectal Carcinoma Patients. Biology, 10(9), 918. https://doi.org/10.3390/biology10090918