
The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Race Analysis
2.1.1. Survey
2.1.2. Sampling
2.1.3. Isolation and Purification
2.1.4. Race Identification
2.2. Disease Assessment
2.3. RAPD Analysis of DNA for P. triticina Races
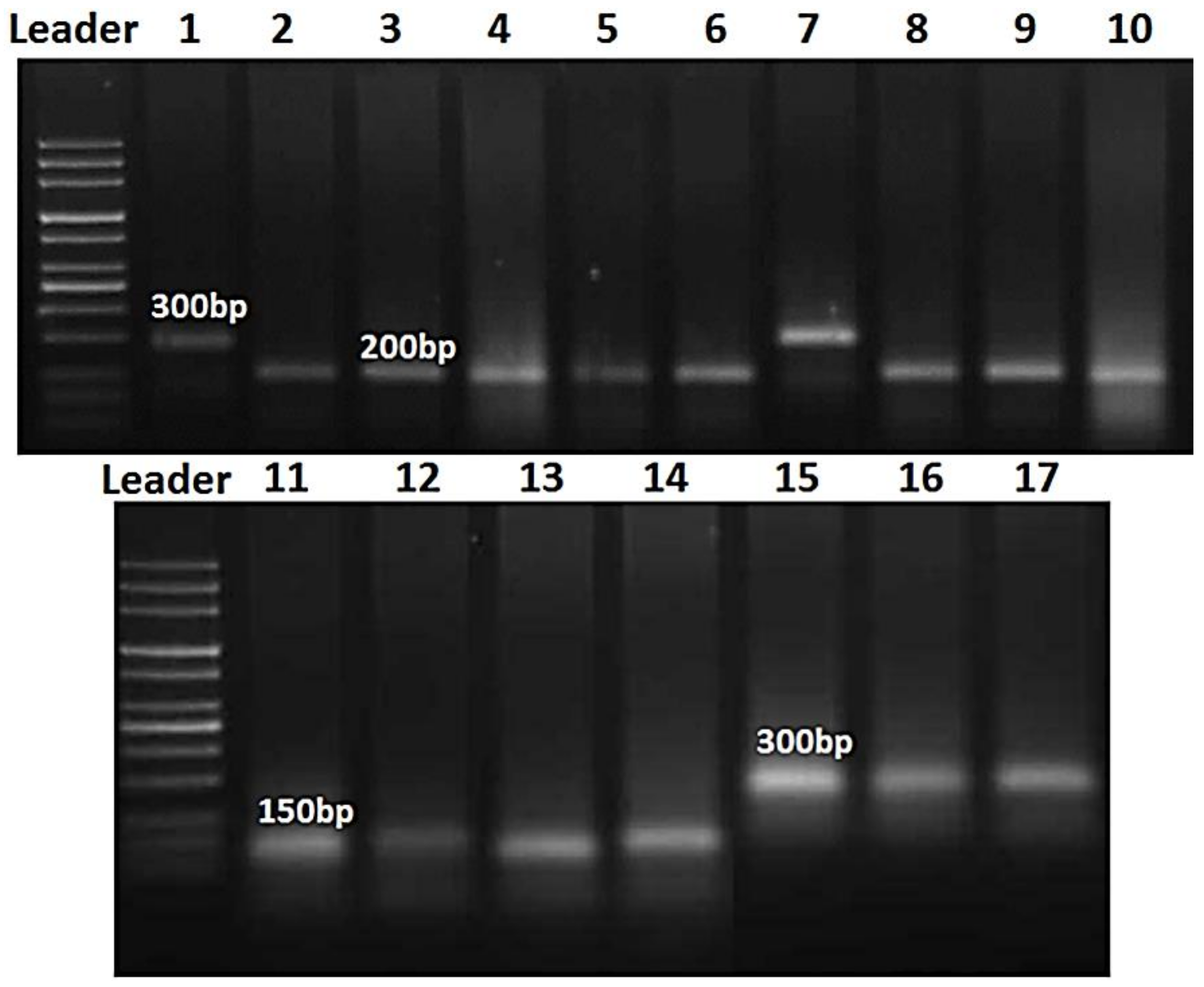
DNA Extraction and PCR Amplification
2.4. Efficacy of Resistant Genes to Wheat Leaf Rust at Seedling Stage
2.5. Molecular Markers Procedure for Identification of Resistance Genes
2.5.1. Plant Sampling
2.5.2. Polymerase Chain Reaction
2.6. The Principal Component Analysis (PCA) and Two-Way Hierarchical Cluster Analysis (HCA)
3. Results
3.1. Assessment of Wheat Cultivars Resistance to Leaf Rust at Adult Stage
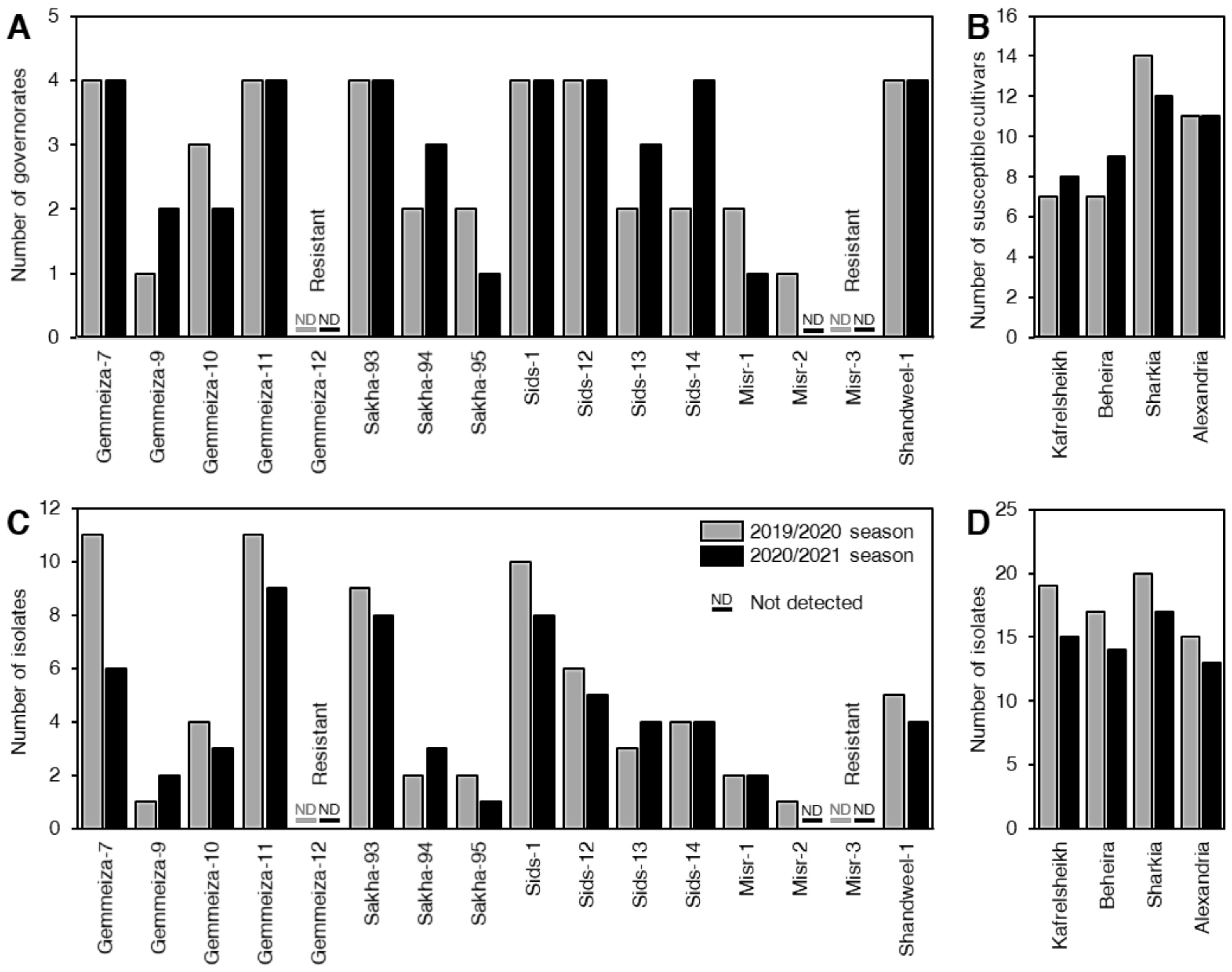
3.2. Distribution of P. triticina Isolates between Different Cultivars, Locations, and Physiological Races
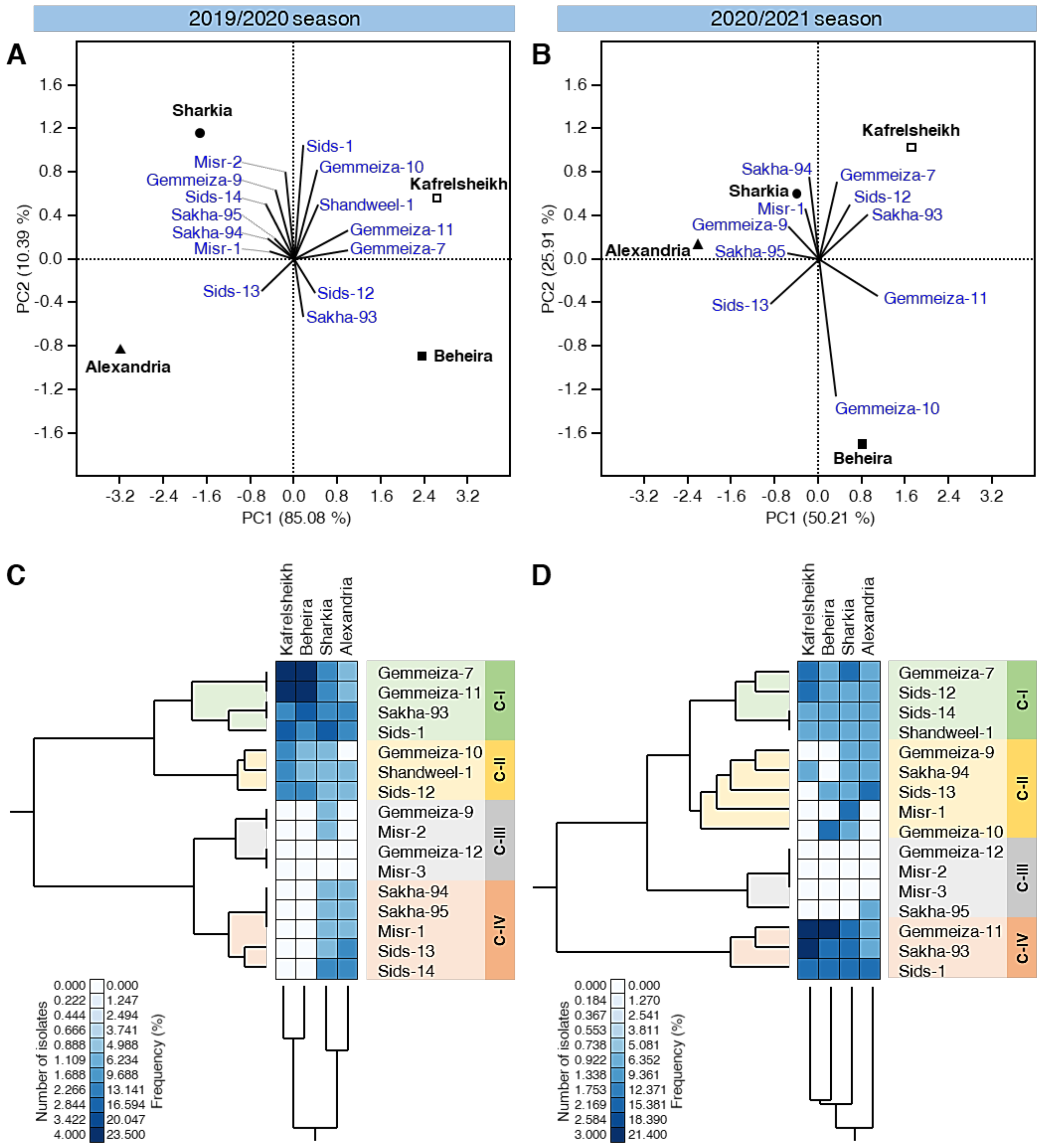
3.3. The Effect of Location-Cultivars Association on the Number of Isolates and Their Frequency (%)
3.4. PCA Reveals Differences in Wheat Cultivation among Different Locations
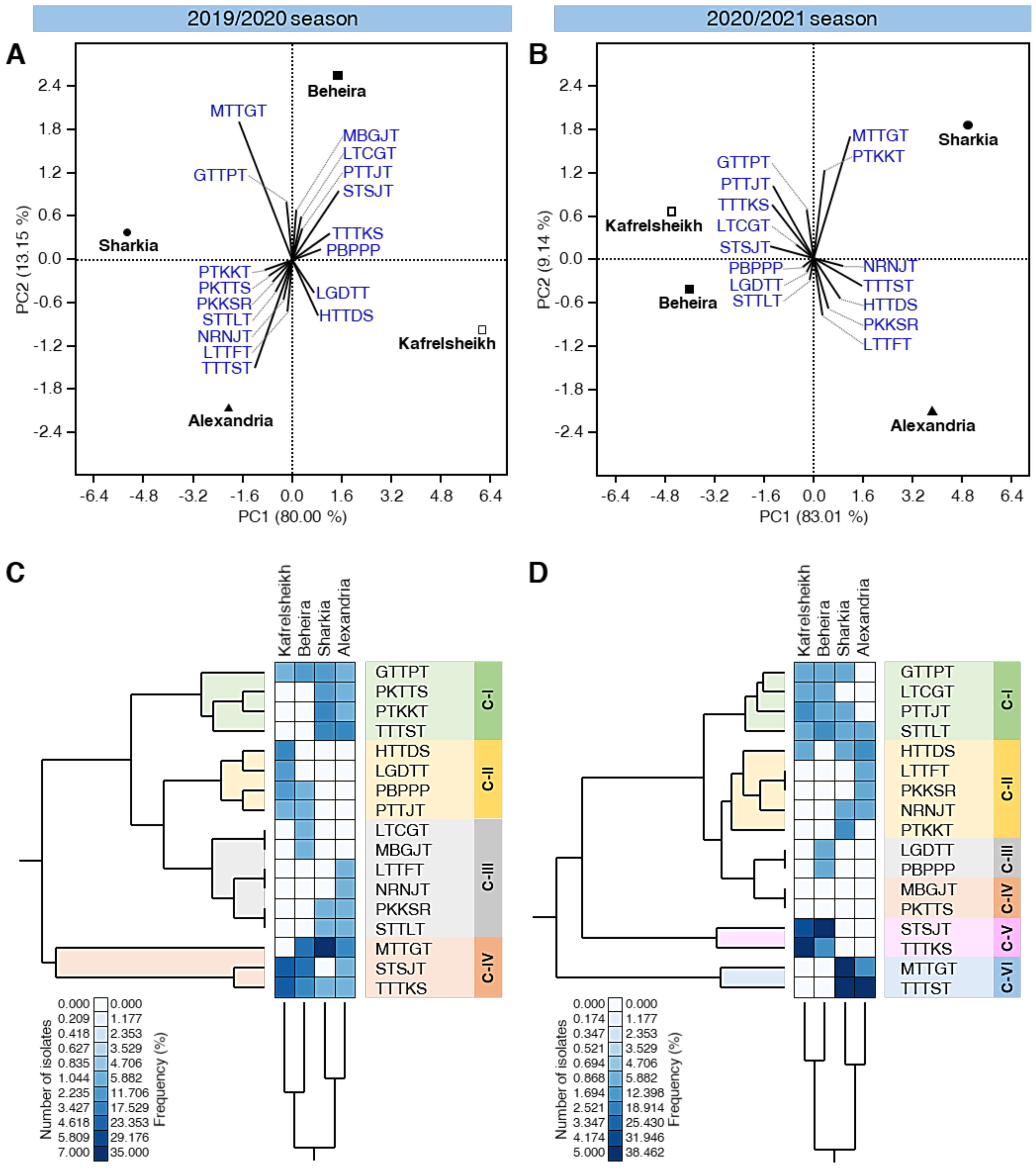
3.5. Race Analysis of P. triticina Isolates
3.6. PCA Reveals Differences in the Distribution of Physiological Races of P. triticina among Locations
3.7. Identification of Physiological Leaf Rust Races on the Egyptian Wheat Cultivars
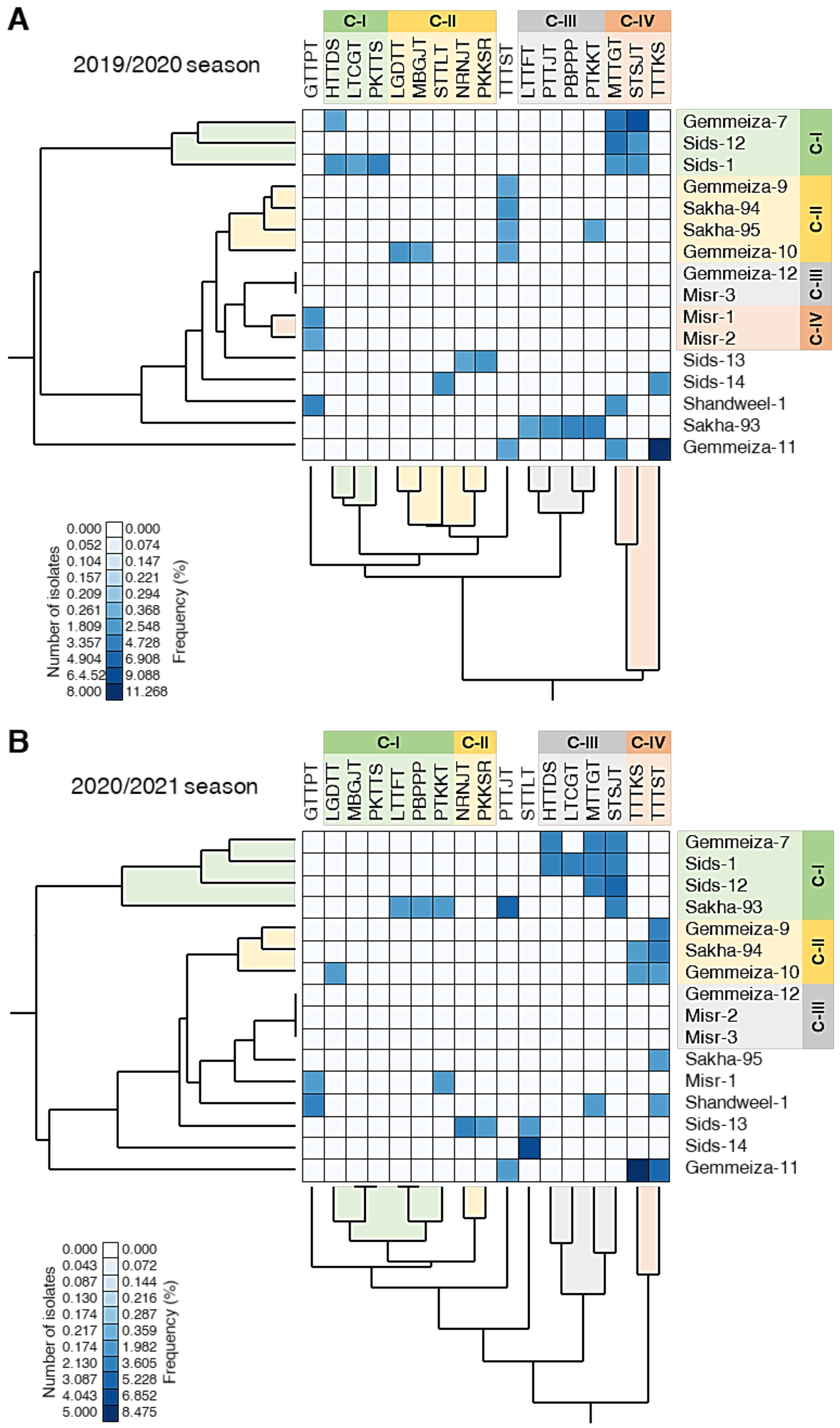
3.8. PCA Reveals Differences in the Distribution of Physiological Races of P. triticina among Cultivars
3.9. Random Amplified Polymorphic DNA (RAPD) Assay Cluster Analysis Using RAPD Markers
3.10. Effectiveness of Leaf Rust Resistance Genes at Seedling Stage
3.11. Molecular Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/statistics/en (accessed on 20 September 2020).
- Statista. Annual Wheat Production in Egypt from 2010 to 2020. Statista Global Data Platform. Available online: https://www.statista.com/statistics/972549/egypt-wheat-production (accessed on 25 June 2021).
- Statista. Principal Importing Countries of Wheat, Flour and Wheat Products from 2014/2015 to 2020/2021. Statista Global Data Platform. Available online: https://www.statista.com/statistics/190435/principle-importing-countries-of-wheat-flour-and-products (accessed on 20 June 2021).
- Wally, A.; Akingbe, O. Grain and Feed Annual: Egypt Is Able to Secure a Steady Supply of Grains during the COVID-19 Pandemic; United States Department of Agriculture (USDA), Foreign Agricultural Service (FAS). 2021. Available online: https://apps.fas.usda.gov/newgainapi/api/Report/DownloadReportByFileName?fileName=Grain+and+Feed+Annual_Cairo_Egypt_03-15-2021.pdf (accessed on 21 June 2021).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Simón, M.R.; Börner, A.; Struik, P.C. Editorial: Fungal wheat diseases: Etiology, breeding, and integrated management. Front. Plant Sci. 2021, 12, 671060. [Google Scholar] [CrossRef] [PubMed]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Ciudad de México, Mexico, 1992; p. 81. Available online: https://rusttracker.cimmyt.org/wp-content/uploads/2011/11/rustdiseases.pdf (accessed on 17 June 2015).
- Huerta-Espino, J.; Singh, R.P.; Germán, S.; McCallum, B.D.; Park, R.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Ali, R.; Omara, R.; Ali, Z.A. Effect of leaf rust infection on yield and technical properties in grains of some Egyptian wheat cultivars. Menoufia J. Plant Prot. 2016, 1, 19–35. [Google Scholar] [CrossRef]
- Nazim, M.; El-Sanawani, M.Z.; Boulot, O.A. Partial resistance to leaf rust in some Egyptian wheat cultivars. In Proceedings of the 6th Congress of the Egyptian Phytopathological Society, Cairo, Egypt, 10–14 June 1990; pp. 77–79. [Google Scholar]
- Soliman, N.E.K.; Abdelbacki, A.M.M.; Najeeb, M.A.; Omara, R.I. geographical distribution of physiologic races of Puccinia triticina and postulation of resistance genes in new wheat cultivars in Egypt. Int. J. Phytopathol. 2012, 1, 73–80. [Google Scholar] [CrossRef]
- Nazim, M.; Aly, M.M.; Shafik, I.; Abd El-Malek, N.I. Frequency of virulence and virulence formula of wheat leaf rust races identified in Egypt during 2004/05-2007/08. Egypt. J. Phytopathol. 2010, 38, 77–88. [Google Scholar]
- Bolton, M.D.; Kolmer, J.A.; Garvin, D.F. Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hak, T.M.; El-Shehedi, A.A.; Nazim, M. The source of inoculum of wheat leaf rust in relation to wind direction. Egypt. J. Phytopathol. 1974, 6, 17–25. [Google Scholar]
- McVey, D.V.; Nazim, M.; Leonard, K.J.; Long, D.L. Patterns of virulence diversity in Puccinia triticina on wheat in Egypt and the United States in 1998–2000. Plant Dis. 2004, 88, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Negm, S.S.; Boulot, O.A.; Hermas, G.A. Virulence dynamics and diversity in wheat leaf rust (Puccinia triticina) populations in Egypt during 2009–2010 and 2010–2011 growing seasons. Egypt. J. Appl. Sci. 2013, 28, 183–212. [Google Scholar]
- El-Orabey, W.M.; Sallam, M.E.; Omara, R.; Abd El-Malik, N.I. Geographical distribution of Puccinia triticina physiologic races in Egypt during 2012–2014 growing seasons. Afr. J. Agric. Res. 2015, 10, 4193–4203. [Google Scholar] [CrossRef] [Green Version]
- El-Orabey, W.M.; Omara, R.I.; Abou-Zeid, M.A. Diversity and virulence dynamics within Puccinia triticina populations in Egypt. J. Plant Prot. Pathol. 2018, 9, 735–745. [Google Scholar] [CrossRef]
- Torabi, M.; Nazari, K.; Afshari, F. Genetic of pathogenicity of Puccinia recondita f. sp. tritici, the causal agent of leaf rust of wheat. Iran. J. Agric. Sci. 2001, 32, 625–635. [Google Scholar]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, J.; Morris, C.; Somers, D.J. Catalogue of gene symbols for wheat. In Proceedings of the 11th International Wheat Genetics Symposium, Brisbane, QLD, Australia, 24–29 August 2008; pp. 1–197. [Google Scholar]
- Singla, J.; Linda, L.; Thomas, W.; Urmil, B.; Simon, G.K.; Beat, K. Characterization of Lr75: A partial, broad-spectrum leaf rust resistance gene in wheat. Theor. Appl. Genet. 2017, 130, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Leonova, I.N.; Ekaterina, S.S.; Elena, A.S. Genome-wide association study of leaf rust resistance in Russian spring wheat varieties. BMC Plant Biol. 2020, 20, 135. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2015–2016 Supplement. Komugi Wheat Genetic Resources Database. 2016. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2015.pdf (accessed on 4 April 2020).
- Kolmer, J.A.; Singh, R.P.; Garvin, D.F.; Viccars, L.; William, H.M.; Huerta-Espino, J.; Ogbonnaya, F.; Raman, H.; Orford, S.; Bariana, H.S.; et al. Analysis of the Lr34/Yr18 rust resistance region in wheat germplasm. Crop. Sci. 2008, 48, 1841–1852. [Google Scholar] [CrossRef] [Green Version]
- Admassu, B.; Lind, V.; Friedt, W.; Ordon, F. Virulence analysis of Puccinia graminis f.sp. tritici populations in Ethiopia with special consideration of Ug99. Plant Pathol. 2009, 58, 362–369. [Google Scholar] [CrossRef]
- Boroujeni, F.R.; Arzani, A.; Afshari, F.; Torabi, M. Postulation of leaf rust resistance genes in Iranian wheat cultivars and breeding lines. Can. J. Plant Pathol. 2011, 33, 550–558. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Liu, J.Q. Virulence and molecular polymorphism in international collections of the wheat leaf rust fungus Puccinia triticina. Phytopathology 2000, 90, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Park, R.F.; Jahoor, A.; Felesenstein, F.G. Population structure of Puccinia recondita in Western Europe during 1995, as assessed by variability in pathogenicity and molecular markers. J. Phytopathol. 2000, 148, 169–179. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia graminis f.sp. tritici; The Agricultural Research Service (ARS): Washington, DC, USA; United States Department of Agriculture (USDA): Washington, DC, USA, 1962; p. 54. [Google Scholar]
- Long, D.L.; Kolmer, J.A. A North American system of nomenclature for Puccinia recondita f. sp. tritici. Phytopathology 1989, 79, 525–529. [Google Scholar] [CrossRef]
- Johnston, C.O.; Browder, L.E. Seventh revision of the international register of physiologic races of Puccinia recondita f.sp. tritici. Plant Dis. Rep. 1966, 50, 756–760. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Devos, K.; Gale, M. The genetic maps of wheat and their potential in plant breeding. Outlook Agric. 1993, 22, 93–99. [Google Scholar] [CrossRef]
- Green, G.J. Stem rust of wheat, barley and rye in Canada in 1964. Can. Plant Dis. Surv. 1965, 45, 23–29. [Google Scholar]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Boulot, O.A.; Aly, A.A. Partial resistance of wheat (Triticum aestivum) to leaf rust (Puccinia triticina) in Egypt. A. Evaluation of seven Egyptian wheat cultivars for partial resistance against leaf rust, under field conditions. Egypt. J. Agric. Res. 2014, 92, 835–850. [Google Scholar]
- Kolmer, J.A. Physiologic Specialization of Puccinia triticina in Canada in 1998. Plant Dis. 2001, 85, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Park, R.F.; Bariana, H.S.; Wellings, C.R.; Wallwork, H. Detection and occurrence of a new pathotype of Puccinia triticina with virulence for Lr24 in Australia. Aust. J. Agric. Res. 2002, 53, 1069–1076. [Google Scholar] [CrossRef]
- Kolmer, J.A. Effect of sexual recombination in two populations of the wheat leaf rust fungus Puccinia recondita. Can. J. Bot. 1992, 70, 359–363. [Google Scholar] [CrossRef]
- McIntosh, R.; Wellings, C.; Park, R. Wheat Rusts: An Atlas of Resistance Genes; CSIRO Publishing: Collingwood, VIC, Australia, 1995; p. 213. [Google Scholar]
- Park, R.; Burdon, J.; Jahoor, A. Evidence for somatic hybridization in nature in Puccinia recondita f. sp. tritici, the leaf rust pathogen of wheat. Mycol. Res. 1999, 103, 715–723. [Google Scholar] [CrossRef]
- Park, R.F.; Burdon, J.J.; McIntosh, R.A. Studies on the origin, spread, and evolution of an important group of Puccinia recondita f. sp. tritici pathotypes in Australasia. Eur. J. Plant Pathol. 1995, 101, 613–622. [Google Scholar] [CrossRef]
- Wang, X.; McCallum, B. Fusion body formation, germ tube anastomosis, and nuclear migration during the germination of urediniospores of the wheat leaf rust fungus, Puccinia triticina. Phytopathology 2009, 99, 1355–1364. [Google Scholar] [CrossRef] [Green Version]
- Kolmer, J.A.; Long, D.L.; Hughes, M.E. Physiologic specialization of Puccinia triticina on wheat in the United States in 2003. Plant Dis. 2005, 89, 1201–1206. [Google Scholar] [CrossRef] [Green Version]
- Abdelbacki, A.M.; Omara, R.I.; Soliman, N.E.; Najeeb, M.A. Molecular markers identification of leaf rust resistant genes Lr19, Lr21, Lr24, Lr47 and Lr51 in selected Egyptian wheat cultivars. Int. J. Phytopathol. 2015, 4, 55–62. [Google Scholar] [CrossRef]
- Omara, R.I.; Abdelaal, K.A. Biochemical, histopathological and genetic analysis associated with leaf rust infection in wheat plants (Triticum aestivum L.). Physiol. Mol. Plant Pathol. 2018, 104, 48–57. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Long, D.L.; Hughes, M.E. Physiologic specialization of Puccinia triticina on wheat in the United States in 2009. Plant Dis. 2011, 95, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Manninger, K. Occurrence and virulence of wheat leaf rust in Hungary during 1999–2001. Plant Prot. Sci. 2002, 38, 408–410. [Google Scholar] [CrossRef]
- Omara, R.; Abdelaal, K. Molecular and genetic analysis of leaf rust resistance genes in two new Egyptian wheat cultivars. Egypt. J. Phytopathol. 2017, 45, 33–52. [Google Scholar] [CrossRef]
- Vida, G.; Gál, M.; Uhrin, A.; Veisz, O.; Wang, Z.; Kiss, T.; Karsai, I.; Bedő, Z. Application of molecular markers in breeding for leaf rust resistance in wheat. In Proceedings of the 60th Tagung der Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs, Irdning, Austria, 24–26 November 2009; pp. 65–71. [Google Scholar]
- Meshkova, L.V.; Rosseeva, L.P.; Korenyuk, E.A.; Belan, I.A. Dynamics of distribution of the wheat leaf rust pathotypes virulent to the cultivars with Lr9 gene in Omsk region. Mikol. Fitopatol. 2012, 46, 397–400. (In Russian) [Google Scholar]
- Gupta, S.K.; Charpe, A.; Prabhu, K.V.; Haq, Q.M.R. Identification and validation of molecular markers linked to the leaf rust resistance gene Lr19 in wheat. Theor. Appl. Genet. 2006, 113, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Bipinraj, A.; Honrao, B.; Prashar, M.; Bhardwaj, S.; Rao, S.; Tamhankar, S. Validation and identification of molecular markers linked to the leaf rust resistance gene Lr28 in wheat. J. Appl. Genet. 2011, 52, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Nemati, Z.; Mostowfizadeh-Ghalamfarsa, R.; Dadkhodaie, A.; Mehrabi, R.; Steffenson, B. Virulence of leaf rust physiological races in Iran from 2010 to 2017. Plant Dis. 2020, 104, 363–372. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Governorate | Season/Number of Samples/Isolates per Governorate | Total | ||||
|---|---|---|---|---|---|---|---|
| 2019/20 | 2020/21 | ||||||
| No. of Samples | No. of Isolates | No. of Samples | No. of Isolates | No. of Samples | No. of Isolates | ||
| 1 | Kafrelsheikh | 25 | 19 | 22 | 15 | 47 | 34 |
| 2 | Beheira | 22 | 17 | 20 | 14 | 42 | 31 |
| 3 | Sharkia | 27 | 20 | 25 | 17 | 52 | 37 |
| 4 | Alexandria | 20 | 15 | 18 | 13 | 38 | 28 |
| Total | 94 | 71 | 85 | 59 | 179 | 130 | |
| No. | Cultivar | Governorates/Seasons/Disease Severity | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Kafrelsheikh | Beheira | Sharkia | Alexandria | ||||||
| 2019/20 | 2020/21 | 2019/20 | 2020/21 | 2019/20 | 2020/21 | 2019/20 | 2020/21 | ||
| 1 | Gemmeiza-7 | 60S a | 70S | 70S | 80S | 80S | 90S | 70S | 90S |
| 2 | Gemmeiza-9 | 5R b | 5MR | 10MR | 5R | 5MS | 10S | SS | 5S |
| 3 | Gemmeiza-10 | 5MS | 5MR | 10MS | 5S | 5S | 10S | 5R | 5MR |
| 4 | Gemmeiza-11 | 70S | 80S | 60S | 90S | 80S | 80S | 70S | 90S |
| 5 | Gemmeiza-12 | 5R | 5MR | TrR | TrMR | 5MR | 5R | 5R | 5MR |
| 6 | Sakha-93 | 50S | 80S | 40S | 70S | 70S | 70S | 60S | 80S |
| 7 | Sakha-94 | 5MR d | 5MS | 5MR | 5R | 5MS | 10MS | TrMS | 5MS |
| 8 | Sakha-95 | 5MR | TrMR | 10MR | 10R | 5S | 5R | 5MS | 5S |
| 9 | Sids-1 | 40S | 60S | 50S | 70S | 70S | 50S | 80S | 80S |
| 10 | Sids-12 | 5MS e | 10MS | 5S | 5MS | 5S | 5MS | 10MS | 5S |
| 11 | Sids-13 | 5MR | 10MR | 10R | 5S | 10S | 10S | 5S | 10S |
| 12 | Sids-14 | 5MR | 5S | 10R | 20MS | 10MS | 10S | 5MS | 10S |
| 13 | Misr-1 | 10R | 5MR | TrMR | 5R | 5MS | 5MS | 5MS | 5MR |
| 14 | Misr-2 | 5R | TrR c | TrMR | TrR | 5MS | 5MR | TrMR | TrMR |
| 15 | Misr-3 | TrMR | TrR | 5MR | TrMR | 5MR | 5R | TrMR | TrR |
| 16 | Shandweel-1 | 5MS | 5S | 5MS | 5S | 10MS | 5MS | 5S | 5S |
| Cultivar | Kafrelsheikh | Beheira | Sharkia | Alexandria | Total | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | 2019/2020 | 2020/2021 | |||||||||||
| NI a | FR b | NI | FR | NI | FR | NI | FR | NI | FR | NI | FR | NI | FR | NI | FR | NI | FR | NI | FR | |
| Gemmeiza-7 | 4 | 21.1 | 2 | 13.3 | 4 | 23.5 | 1 | 7.1 | 2 | 10.0 | 2 | 12.0 | 1 | 6.7 | 1 | 7.7 | 11 | 15.5 | 6 | 10.2 |
| Gemmeiza-9 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 5.0 | 1 | 5.9 | 0 | 0.0 | 1 | 7.7 | 1 | 1.4 | 2 | 3.4 |
| Gemmeiza-10 | 2 | 10.5 | 0 | 0.0 | 1 | 5.9 | 2 | 14.3 | 1 | 5.0 | 1 | 5.9 | 0 | 0.0 | 0 | 0.0 | 4 | 5.6 | 3 | 5.1 |
| Gemmeiza-11 | 4 | 21.1 | 3 | 20.0 | 4 | 23.5 | 3 | 21.4 | 2 | 10.0 | 2 | 12.0 | 1 | 6.7 | 1 | 7.7 | 11 | 15.5 | 9 | 15.3 |
| Gemmeiza-12 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| Sakha-93 | 2 | 10.5 | 3 | 20.0 | 3 | 17.7 | 2 | 14.3 | 2 | 10.0 | 2 | 12.0 | 2 | 13.0 | 1 | 7.7 | 9 | 12.7 | 8 | 13.6 |
| Sakha-94 | 0 | 0.0 | 1 | 6.7 | 0 | 0.0 | 0 | 0.0 | 1 | 5.0 | 1 | 5.9 | 1 | 6.7 | 1 | 7.7 | 2 | 2.8 | 3 | 5.1 |
| Sakha-95 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 5.0 | 0 | 0.0 | 1 | 6.7 | 1 | 7.7 | 2 | 2.8 | 1 | 1.7 |
| Sids-1 | 3 | 15.8 | 2 | 13.3 | 2 | 11.8 | 2 | 14.3 | 3 | 15.0 | 2 | 12.0 | 2 | 13.0 | 2 | 15.0 | 10 | 14.1 | 8 | 13.6 |
| Sids-12 | 2 | 10.5 | 2 | 13.3 | 2 | 11.8 | 1 | 7.1 | 1 | 5.0 | 1 | 5.9 | 1 | 6.7 | 1 | 7.7 | 6 | 8.5 | 5 | 8.5 |
| Sids-13 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 7.1 | 1 | 5.0 | 1 | 5.9 | 2 | 13.0 | 2 | 15.0 | 3 | 4.2 | 4 | 6.8 |
| Sids-14 | 0 | 0.0 | 1 | 6.7 | 0 | 0.0 | 1 | 7.1 | 2 | 10.0 | 1 | 5.9 | 2 | 13.0 | 1 | 7.7 | 4 | 5.6 | 4 | 6.8 |
| Misr-1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 5.0 | 2 | 12.0 | 1 | 6.7 | 0 | 0.0 | 2 | 2.8 | 2 | 3.4 |
| Misr-2 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 5.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 1.4 | 0 | 0.0 |
| Misr-3 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| Shandweel-1 | 2 | 10.5 | 1 | 6.7 | 1 | 5.9 | 1 | 7.1 | 1 | 5.0 | 1 | 12.0 | 1 | 6.7 | 1 | 7.7 | 5 | 7.1 | 4 | 6.8 |
| Total | 19 | - | 15 | - | 17 | - | 14 | - | 20 | - | 17 | - | 15 | - | 13 | - | 71 | - | 59 | - |
| No. | Race | Virulence Formula | Season/No. of Isolates and Frequency (%) | Total | ||||
|---|---|---|---|---|---|---|---|---|
| 2019/2020 | 2020/2021 | |||||||
| NI a | FR b | NI | FR | NI | FR | |||
| 1 | GTTPT | 2a, 9, 16, 24, 26, 3Ka, 11, 17, 30, 10, 21, 2b, 14b, 15,36, 42 | 6 | 8.45 | 3 | 5.08 | 9 | 6.92 |
| 2 | HTTDS | 2a, 3, 9, 16, 24, 26, 3Ka, 11, 17, 30, 21, 14b, 15, 36 | 3 | 4.23 | 4 | 6.78 | 7 | 5.38 |
| 3 | LGDTT | 1, 9, 16, 24, 26, 17, 10, 18, 21, 2b, 14b, 15, 36, 42 | 2 | 2.82 | 1 | 1.69 | 3 | 2.31 |
| 4 | LTCGT | 1, 9, 16, 24, 26, 30, 18, 14b, 15, 36, 42 | 1 | 1.41 | 2 | 3.39 | 3 | 2.31 |
| 5 | LTTFT | 1, 9, 16, 24, 26, 3Ka, 11, 17, 30, 21, 14b, 15, 36, 42 | 1 | 1.41 | 1 | 1.69 | 2 | 1.54 |
| 6 | MBGJT | 1, 3, 11, 18, 21, 14b, 15, 36, 42 | 1 | 1.41 | - | 0.00 | 1 | 0.77 |
| 7 | MTTGT | 1,3, 9, 16, 24, 26, 3Ka, 11, 17, 30, 18, 14b, 15, 36, 42 | 14 | 19.70 | 7 | 11.90 | 21 | 16.20 |
| 8 | NRNJT | 1, 2c, 9, 16, 26, 3Ka, 17, 18, 21, 14b, 15, 36, 42 | 1 | 1.41 | 2 | 3.39 | 3 | 2.31 |
| 9 | PBPPP | 1, 2c, 3, 3Ka, 17, 30, 10, 21, 2b, 14b, 36, 42 | 3 | 4.23 | 1 | 1.69 | 4 | 3.08 |
| 10 | PKKSR | 1, 2c, 3, 16, 24, 26, 11, 17, 30, 10, 18, 21, 14b, 15, 42 | 2 | 2.82 | 1 | 1.69 | 3 | 2.31 |
| 11 | PKTTS | 1, 2c, 3, 16, 24, 26, 3Ka, 11, 17, 30, 10, 18, 21, 2b, 14b, 15,36 | 3 | 4.23 | - | 0.00 | 3 | 2.31 |
| 12 | PTKKT | 1, 2c, 3, 9, 16, 24, 26, 11, 17, 30, 18, 21, 2b, 14b, 15, 36, 42 | 4 | 5.63 | 2 | 3.39 | 6 | 4.62 |
| 13 | PTTJT | 1, 2c, 3, 9, 16, 24, 26, 3Ka, 11, 17, 30, 18, 21, 14b, 15, 36, 42 | 2 | 2.82 | 4 | 6.78 | 6 | 4.62 |
| 14 | STSJT | 1, 2a, 2c, 9, 16, 24, 26, 3Ka, 11,17, 18, 21, 14b, 15, 36, 42 | 10 | 14.10 | 9 | 15.30 | 19 | 14.60 |
| 15 | STTLT | 1, 2a, 2c, 9, 16, 24, 26, 3Ka, 11, 17, 30, 10, 14b, 15, 36, 42 | 2 | 2.82 | 5 | 8.47 | 7 | 5.38 |
| 16 | TTTKS | 1, 2a, 2c, 3, 9, 16, 24, 26, 3Ka, 11, 17, 30, 18, 21, 2b, 14b, 15, 36 | 10 | 14.10 | 7 | 11.90 | 17 | 13.10 |
| 17 | TTTST | 1, 2a, 2c, 3, 9, 16, 24, 26, 3Ka, 11, 17, 30, 10, 18, 21, 14b, 15, 36, 42 | 6 | 8.45 | 10 | 16.90 | 16 | 12.30 |
| Total | 71 | 100.0 | 59 | 100.0 | 130 | 100.0 | ||
| Cultivar | Governorate/Leaf Rust Races | |||
|---|---|---|---|---|
| Kafrelsheikh | Beheira | Sharkia | Alexandria | |
| 2019/2020 Season | ||||
| Gemmeiza-7 | HTTDS STSJT | MTTGT - | MTTGT - | STSJT - |
| Gemmeiza-9 | - | - | TTTST | - |
| Gemmeiza-10 | LGDTT | MBGJT | TTTST | - |
| Gemmeiza-11 | TTTKS | TTTKS | MTTGT | TTTST |
| Gemmeiza-12 | - | - | - | - |
| Sakha-93 | PBPPP PTTJT | PBPPP PTTJT | PTKKT - | LTTFT PTKKT |
| Sakha-94 | - | - | TTTST | TTTST |
| Sakha-95 | - | - | PTKKT | TTTST |
| Sids-1 | HTTDS STSJT | STSJT LTCGT | MTTGT PKTTS | PKTTS MTTGT |
| Sids-12 | STSJT | STSJT | MTTGT | MTTGT |
| Sids-13 | - - | - - | PKKSR - | NRNJT PKKSR |
| Sids-14 | - - | - - | TTTKS STTLT | TTTKS STTLT |
| Misr-1 | - | - | GTTPT | GTTPT |
| Misr-2 | - | - | GTTPT | - |
| Misr-3 | - | - | - | - |
| Shandweel-1 | GTTPT | GTTPT | MTTGT | MTTGT |
| 2020/2021 season | ||||
| Gemmeiza-7 | STSJT HTTDS | STSJT - | MTTGT - | HTTDS - |
| Gemmeiza-9 | - | - | TTTST | TTTST |
| Gemmeiza-10 | - - | LGDTT TTTKS | TTTST | - |
| Gemmeiza-11 | PTTJT TTTKS | TTTKS - | TTTST - | TTTST - |
| Gemmeiza-12 | - | - | - | - |
| Sakha-93 | STSJT PTTJT | PTTJT PBPPP | PTKKT PTTJT | LTTFT - |
| Sakha-94 | TTTKS | - | TTTST | TTTST |
| Sakha-95 | - | - | - | TTTST |
| Sids-1 | STSJT LTCGT | STSJT LTCGT | MTTGT HTTDS | HTTDS MTTGT |
| Sids-12 | STSJT | STSJT | MTTGT | MTTGT |
| Sids-13 | - - | STTLT - | NRNJT - | NRNJT PKKSR |
| Sids-14 | STTLT | STTLT | STTLT | STTLT |
| Misr-1 | - - | - - | GTTPT PTKKT | - - |
| Misr-2 | - | - | - | - |
| Misr-3 | - | - | - | - |
| Shandweel-1 | GTTPT | GTTPT | MTTGT | TTTST |
| No. | Lr Genes | Season/No. of Isolates and Virulence Frequency (%) | Total | ||||
|---|---|---|---|---|---|---|---|
| 2019/2020 | 2020/2021 | ||||||
| NI a | FR b | NI | FR | NI | FR | ||
| 1 | Lr1 | 62 | 87.32 | 52 | 88.14 | 114 | 87.69 |
| 2 | Lr2a | 37 | 52.11 | 38 | 64.41 | 75 | 57.69 |
| 3 | Lr2b | 28 | 39.44 | 14 | 23.73 | 42 | 32.31 |
| 4 | Lr2c | 43 | 60.56 | 41 | 69.49 | 84 | 64.62 |
| 5 | Lr3 | 48 | 67.61 | 36 | 61.02 | 84 | 64.62 |
| 6 | Lr3ka | 59 | 83.1 | 49 | 83.05 | 108 | 83.08 |
| 7 | Lr9 | 62 | 87.32 | 57 | 96.61 | 119 | 91.54 |
| 8 | Lr10 | 27 | 38.03 | 26 | 44.07 | 53 | 40.77 |
| 9 | Lr11 | 64 | 90.14 | 53 | 89.83 | 117 | 90.00 |
| 10 | Lr14b | 71 | 100.00 | 59 | 100.00 | 130 | 100.00 |
| 11 | Lr15 | 68 | 95.77 | 58 | 98.31 | 126 | 96.92 |
| 12 | Lr16 | 67 | 94.37 | 58 | 98.31 | 125 | 96.15 |
| 13 | Lr17 | 69 | 97.18 | 57 | 96.61 | 126 | 96.92 |
| 14 | Lr18 | 57 | 80.28 | 46 | 77.97 | 103 | 79.23 |
| 15 | Lr19 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 |
| 16 | Lr21 | 54 | 76.06 | 45 | 76.27 | 99 | 76.15 |
| 17 | Lr24 | 67 | 94.37 | 58 | 98.31 | 125 | 96.15 |
| 18 | Lr26 | 43 | 60.56 | 40 | 67.80 | 83 | 63.85 |
| 19 | Lr28 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 |
| 20 | Lr30 | 57 | 80.28 | 47 | 79.66 | 104 | 80.00 |
| 21 | Lr36 | 69 | 97.18 | 58 | 98.31 | 127 | 97.69 |
| 22 | Lr42 | 52 | 73.24 | 42 | 71.19 | 94 | 72.31 |
| Total | 71 | - | 59 | - | 130 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omara, R.I.; Nehela, Y.; Mabrouk, O.I.; Elsharkawy, M.M. The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars. Biology 2021, 10, 925. https://doi.org/10.3390/biology10090925
Omara RI, Nehela Y, Mabrouk OI, Elsharkawy MM. The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars. Biology. 2021; 10(9):925. https://doi.org/10.3390/biology10090925
Chicago/Turabian StyleOmara, Reda Ibrahim, Yasser Nehela, Ola Ibrahim Mabrouk, and Mohsen Mohamed Elsharkawy. 2021. "The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars" Biology 10, no. 9: 925. https://doi.org/10.3390/biology10090925
APA StyleOmara, R. I., Nehela, Y., Mabrouk, O. I., & Elsharkawy, M. M. (2021). The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars. Biology, 10(9), 925. https://doi.org/10.3390/biology10090925