

A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete
Abstract
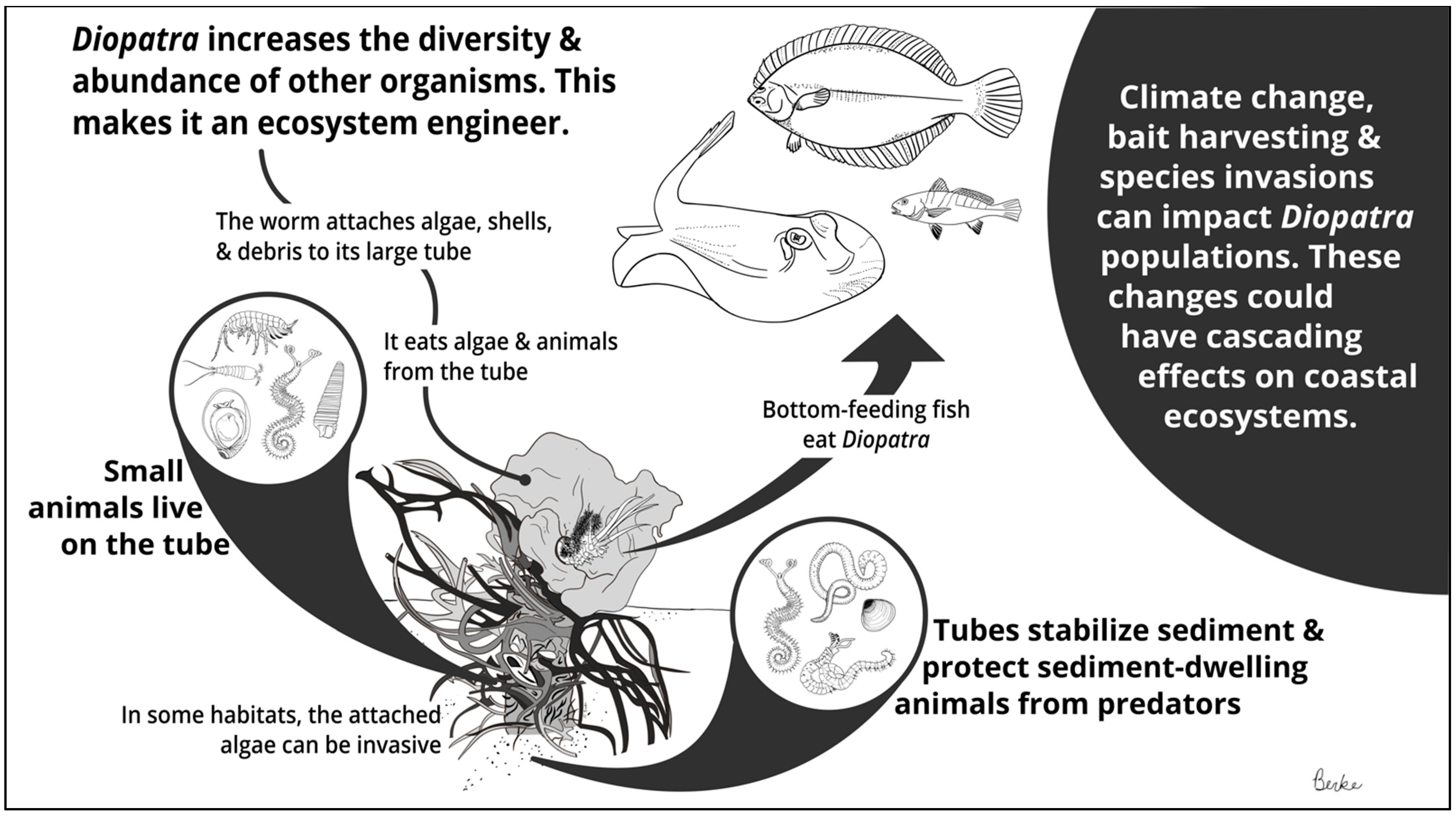
:Simple Summary
Abstract
1. Introduction
Taxonomic Caveat
2. Natural History
3. Physical Effects of the Tube
4. Facilitation of Macroalgae and Plants

{kind=link}
| Study | Species | Location | Effects |
|---|---|---|---|
| Mangum et al., 1968 [31] | D. cuprea | Chesapeake Bay, Virginia, USA | 20 algal species identified from tubes |
| Harwell and Orth 2001 [56] | D. cuprea | Chesapeake Bay, Virginia, USA | Tubes facilitate reproductive seagrass shoots |
| Thomsen 2004 [62] | D. cuprea | Hog Island Bay, Virginia, USA | Tubes facilitate Ulva and A. vermiculophyllum |
| Thomsen & McGlathery 2005; Thomsen et al., 2005 [57,63] | D. cuprea | Hog Island Bay, Virginia, USA | Tubes facilitate invasive alga Agarophyton vermiculophyllum |
| Berke 2012 [11] | D. cuprea | Northwest Atlantic (Massachusetts through Florida, USA) | Tubes support a total of 34 species (as many as 15 species within a single region) |
5. Facilitation of Infauna
| Study | Species | Location | Densities Tested | Effects |
|---|---|---|---|---|
| Woodin 1978 [4] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 6 · 0.01 m−2 | ↑ infaunal richness and abundance at 6 · 0.01 m−2 |
| Woodin 1981 [12] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 6 · 0.01 m−2 | ↑ infaunal abundance at 6 · 0.01 m−2 |
| Bell and Woodin 1984 [65] | D. cuprea | Tom’s Cove, Virginia, USA | 0, 1, 3, 6 · 0.01 m−2 | ↑ polychaete abundance at 6 · 0.01 m−2; no effect on meiofauna |
| Bailey-Brock 1984 [29] | D. leuckarti | Niu Valley, Hawaii, USA | “mounds” up to 21,800 m−2 | 28 species from 7 phyla are associated with mounds |
| Luckenbach 1984 [69] | D. cuprea | North Inlet, South Carolina, USA | 0, ≥9 · 0.01 m−2 | ↑ infaunal abundance near tubes |
| Luckenbach 1984 [70] | D. cuprea | North Inlet, South Carolina, USA | 0, ≥10 · 0.01 m−2 | ↑ infaunal abundance in areas of high tube density |
| Ban and Nelson 1987 [66] | D. cuprea | Indian River Lagoon, Florida, USA | 0, 1, 4 · 0.01 m−2 | No effect |
| Ambrose & Anderson 1990 [71] | D. ornata | Pendleton Artificial Reef, California, USA | Inside vs. outside “beds” | ↑ richness and abundance of infauna and decapods |
| Thomsen et al., 2011 [23] | Diopatra-like onuphid * | Inhaca Island, Mozambique | 0, 1 · 0.01 m−2 | ↑ richness and abundance around single tubes |
| Santos and Aviz 2018 [20] † | D. cuprea * | Algodoal-Maiandeua, Brazil | 0 vs. 25–75 m−2, but each sample from the Diopatra area included only 1 | Seasonal ↑ richness and abundance |
6. Facilitation of Epibenthic Fauna
7. Food Web Connections
8. Human Impacts
8.1. Range Expansions and Climate Change
8.2. Bait Harvesting
8.3. Interactions with Invasive Species
9. Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Verrill, A.E.; Smith, S.I. Report upon the Invertebrate Animals on Vineyard Sound and Adjacent Waters, with an Account of the Physical Features of the Region; U.S. Goverment Printing Office: Washington, DC, USA, 1874.
- Linville, H.R. The Natural History of Some Tube-Forming Annelids. Mark Anniversary Volume; to Edward Laurens Mark, Hersey Professor of Anatomy and Director of the Zoological Laboratory at Harvard University; H. Holt and Company: New York, NY, USA, 1903; pp. 227–237. [Google Scholar]
- Saint-Joseph, B. de Les Annélides Polychètes des Côtes de France (Manche et Océan). Ann. Des Sci. Nat. (Zo-Ologie Paléontol.) 1898, 5, 209–222. [Google Scholar]
- Woodin, S.A. Refuges, Disturbance, and Community Structure: A Marine Soft-Bottom Example. Ecology 1978, 59, 274–284. [Google Scholar] [CrossRef]
- Wright, J.P.; Jones, C. The Concept of Organisms as Ecosystem Engineers Ten Years on: Progress, Limitations, and Challenges. BioScience 2006, 56, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Berke, S.K. Functional Groups of Ecosystem Engineers: A Proposed Classification with Comments on Current Issues. Integr. Comp. Biol. 2010, 50, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, O. Catalogue of the Polychaeta; Allan Hancock Foundation, University of Southern California Press: Los Angeles, CA, USA, 1959. [Google Scholar]
- Day, J.H. Polychaeta of Southern Africa Part 2: Sedentaria; Trustees of the British Museum (Natural History): London, UK, 1967. [Google Scholar]
- Bell, S.S.; Coen, L.D. Investigations on epibenthic meiofauna I. Abundances on and repopulation of the tube-caps of Diopatra cuprea (Polychaeta: Onuphidae) in a subtropical system. Mar. Biol. 1982, 67, 303–309. [Google Scholar] [CrossRef]
- Diaz, R.J.; Cutter, G.R.; Able, K.W. The importance of physical and biogenic structure to juvenile fishes on the shallow inner continental shelf. Estuaries 2003, 26, 12–20. [Google Scholar] [CrossRef]
- Berke, S.K. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Prog. Ser. 2012, 447, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Woodin, S. Disturbance and Community Structure in a Shallow Water Sand Flat. Ecology 1981, 62, 1052–1066. [Google Scholar] [CrossRef]
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesian region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505–535. [Google Scholar]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes—Fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2018, 32, 1–9. [Google Scholar]
- Elgetany, A.H.; Van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species delineation in the speciation grey zone—The case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar] [CrossRef]
- Hektoen, M.M.; Willassen, E.; Budaeva, N. Phylogeny and Cryptic Diversity of Diopatra (Onuphidae, Annelida) in the East Atlantic. Biology 2022, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Choe, S. On the Life History of the Polychaete Worm, Diopatra neapolitana Delle Chiaje. Nippon Suisan Gakkaishi 1960, 26, 430–437. [Google Scholar] [CrossRef]
- Parameswaran, V.S. Distribution of Diopatra neapolitana Delle Chiaje (Polychaeta) in the South West Coast of India. Indian J. Mar. Sci. 1973, 2, 62–63. [Google Scholar]
- Fauchald, K.; Berke, S.K.; Woodin, S.A. Diopatra (Onuphidae: Polychaeta) from Intertidal Sediments in Southwestern Europe. Zootaxa 2012, 3395, 47–58. [Google Scholar] [CrossRef]
- Santos, T.; Aviz, D. Macrobenthic fauna associated with Diopatra cuprea (Onuphidae: Polychaeta) tubes on a macrotidal sandy beach of the Brazilian Amazon Coast. J. Mar. Biol. Assoc. UK 2018, 99, 751–759. [Google Scholar] [CrossRef]
- Monro, C. XVII.—On the post-larval stage in Diopatra cuprea, Bosc, a Polychætous Annelid of the family Eunicidæ. Ann. Mag. Nat. Hist. 1924, 14, 193–199. [Google Scholar] [CrossRef]
- De, C. Application of a Biological Tool for Estimating Current Annual Rates of Erosion and Deposition in Modern Coastal Environments: A Case Study in the Bay of Bengal Coast. Mar. Georesour. Geotechnol. 2002, 20, 209–220. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Muth, M.F.; McGlathery, K.J. Tube-forming polychaetes enhance invertebrate diversity and abundance in sandy sediments of Mozambique, Africa. Afr. J. Mar. Sci. 2011, 33, 327–332. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2020, 191, 637–671. [Google Scholar] [CrossRef]
- Hartman, O. Atlas of the Sedentariate Polychaetous Annelids from California; Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1969. [Google Scholar]
- Paxton, H.; Fadlaoui, S.; Lechapt, J.P. Diopatra marocensis, a New Brooding Species of Onuphidae (Annelida, Polychaeta). J. Mar. Biol. Assoc. UK 1995, 75, 949–955. [Google Scholar] [CrossRef]
- Cunha, T.; Hall, A.; Queiroga, H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fish. Res. 2005, 76, 56–66. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H. Ecology of the tube-building polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community structure, and sediment stabilizing properties. Zool. J. Linn. Soc. 1984, 80, 191–199. [Google Scholar] [CrossRef]
- Hakkim, V.M.A. Salinity Tolerance of Diopatra neapolitana Delle Chiaje: Annelida-Polychaeta. Indian J. Mar. Sci. 1975, 4, 99–101. [Google Scholar]
- Mangum, C.P.; Santos, S.L.; Rhodes, W.R. Distribution and feeding in the onuphid polychaete, Diopatra cuprea (Bosc). Mar. Biol. 1968, 2, 33–40. [Google Scholar] [CrossRef]
- Peckol, P.; Baxter, D. Population Dynamics of the Onuphid Polychaete Diopatra cuprea (Bosc) along a Tidal Exposure Gradient. Estuar. Coast. Shelf Sci. 1986, 22, 371–377. [Google Scholar] [CrossRef]
- Myers, A.C. Some Palaeoichnological Observations on the Tube of Diopatra cuprea (Bosc): Polychaeta, Onuphidae. In Trace Fossils; Seel House Press: Liverpool, UK, 1970; pp. 331–334. [Google Scholar]
- Myers, A.C. Tube-worm-sediment relationships of Diopatra cuprea (Polychaeta: Onuphidae). Mar. Biol. 1972, 17, 350–356. [Google Scholar] [CrossRef]
- Pettibone, M.H. Polychaete Worms of the New England Region. I. Aphroditidae through Trochochaetidae. Bull. USA Natl. Mus. 1963, 227, 1–361. [Google Scholar] [CrossRef]
- Wethey, D.S.; Woodin, S.A.; Berke, S.K.; Dubois, S.F. Climate hindcasts: Exploring the disjunct distribution of Diopatra biscayensis. Invertebr. Biol. 2016, 135, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.L. The role of drift kelp in the population ecology of a Diopatra ornata Moore (Polychaeta: Onuphidae) ecotone. J. Exp. Mar. Biol. Ecol. 1992, 156, 253–272. [Google Scholar] [CrossRef]
- Berke, S.K.; Miller, M.; Woodin, S.A. Modelling the Energy—Mortality Trade-Offs of Invertebrate Decorating Behaviour. Evol. Ecol. Res. 2006, 8, 1409–1425. [Google Scholar]
- Brenchley, G.A. Predator detection and avoidance: Ornamentation of tube-caps of Diopatra spp. (Polychaeta: Onuphidae). Mar. Biol. 1976, 38, 179–188. [Google Scholar] [CrossRef]
- Berke, S.K.; Woodin, S.A. Tube Decoration May Not Be Cryptic for Diopatra cuprea (Polychaeta: Onuphidae). Biol. Bull. 2008, 214, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, G.A.; Tidball, J.G. Tube-Cap Orientations of Diopatra cuprea (Bosc)(Polychaeta): The Compromise between Physiology and Foraging. Mar. Behav. Physiol. 1980, 7, 1–13. [Google Scholar] [CrossRef]
- Miller, U.C.; Bock, M.J.; Turner, E.J. Deposit and suspension feeding in oscillatory flows and sediment fluxes. J. Mar. Res. 1992, 50, 489–520. [Google Scholar] [CrossRef]
- Fager, E.W. Marine Sediments: Effects of a Tube-Building Polychaete. Science 1964, 143, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Featherstone, R.P.; Risk, M.J. Effect of tube-building polychaetes on intertidal sediments on the Minas Basin, Bay of Fundy. J. Sediment. Res. 1977, 47, 446–450. [Google Scholar] [CrossRef]
- Rhoads, D.C.; Yingst, J.Y.; Ullman, W.J. Seafloor Stability in Central Long Island Sound: Part I. Temporal Changes in Erodibility of Fine-Grained Sediment. In Estuarine Interactions; Academic Press: New York, NY, USA, 1978; pp. 221–244. [Google Scholar]
- Bolam, S.G.; Fernandes, T.F. Dense aggregations of tube-building polychaetes: Response to small-scale disturbances. J. Exp. Mar. Biol. Ecol. 2002, 269, 197–222. [Google Scholar] [CrossRef]
- Eckman, J.E.; Nowell, A.R.M.; Jumars, P.A. Sediment Destabilization by Animal Tubes. J. Mar. Res. 1981, 39, 361–374. [Google Scholar]
- Eckman, J.E.; Nowell, A.R.M. Boundary skin friction and sediment transport about an animal-tube mimic. Sedimentology 1984, 31, 851–862. [Google Scholar] [CrossRef]
- Eckman, J.E. Flow disruption by an animal-tube mimic affects sediment bacterial colonization. J. Mar. Res. 1985, 43, 419–435. [Google Scholar] [CrossRef]
- Eckman, J.E.; Thistle, D. Effects of flow about a biologically produced structure on harpacticoid copepods in San Diego Trough. Deep Sea Res. Part. A Oceanogr. Res. Pap. 1991, 38, 1397–1416. [Google Scholar] [CrossRef]
- Friedrichs, M.; Graf, G.; Springer, B. Skimming flow induced over a simulated polychaete tube lawn at low population densities. Mar. Ecol. Prog. Ser. 2000, 192, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Keller, E.L.; Berke, S.K.; Needham, C.N.; Salerno, C.R. A Double-Edged Sword: Infaunal Responses to Agarophyton vermicu-lophyllum in the Mid-Atlantic United States. Estuaries Coasts J. ERF 2019, 42, 1924–1937. [Google Scholar] [CrossRef]
- Ong, B.; Krishnan, S. Changes in the macrobenthos community of a sand flat after erosion. Estuar. Coast. Shelf Sci. 1995, 40, 21–33. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Altieri, A.; Tuya, F.; Gulbransen, D.; McGlathery, K.J.; Holmer, M.; Silliman, B.R. Habitat Cascades: The Conceptual Context and Global Relevance of Facilitation Cascades via Habitat Formation and Modification. Integr. Comp. Biol. 2010, 50, 158–175. [Google Scholar] [CrossRef]
- Gribben, P.E.; Angelini, C.; Altieri, A.H.; Bishop, M.J.; Thomsen, M.S.; Bulleri, F. Facilitation Cascades in Marine Ecosystems: A Synthesis and Future Directions. Oceanogr. Mar. Biol. 2019, 57, 127–168. [Google Scholar]
- Harwell, M.C.; Orth, R.J. Influence of a tube-dwelling polychaete on the dispersal of fragmented reproductive shoots of eelgrass. Aquat. Bot. 2001, 70, 1–7. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Gurgel, C.F.D.; Fredericq, S.; McGlathery, K.J. Gracilaria vermiculophylla (Rhodophyta, Gracilariales) in Hog Island Bay, Virginia: A Cryptic Alien and Invasive Macroalga and Taxonomic Correction. J. Phycol. 2005, 42, 139–141. [Google Scholar] [CrossRef]
- Wright, J.T.; Byers, J.E.; Devore, J.L.; Sotka, E.E. Engineering or food? mechanisms of facilitation by a habitat-forming invasive seaweed. Ecology 2014, 95, 2699–2706. [Google Scholar]
- Hu, Z.-M.; Juan, L.-B. Adaptation mechanisms and ecological consequences of seaweed invasions: A review case of agarophyte Gracilaria vermiculophylla. Biol. Invasions 2013, 16, 967–976. [Google Scholar] [CrossRef]
- Johnston, C.; Lipcius, R. Exotic macroalga Gracilaria vermiculophylla provides superior nursery habitat for native blue crab in Chesapeake Bay. Mar. Ecol. Prog. Ser. 2012, 467, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.S.; Ramus, A.P.; Long, Z.T.; Silliman, B.R. A seaweed increases ecosystem multifunctionality when invading bare mudflats. Biol. Invasions 2018, 21, 27–36. [Google Scholar] [CrossRef]
- Thomsen, M.S. Macroalgal Distribution Patterns and Ecological Performances in a Tidal Coastal Lagoon, with Emphasis on the Non-indigenous Codium fragile. Estuaries Coasts 2006, 29, 465–473. [Google Scholar] [CrossRef]
- Thomsen, M.; McGlathery, K. Facilitation of macroalgae by the sedimentary tube forming polychaete Diopatra cuprea. Estuar. Coast. Shelf Sci. 2005, 62, 63–73. [Google Scholar] [CrossRef]
- Eckman, J.E. Hydrodynamic processes affecting benthic recruitment1. Limnol. Oceanogr. 1983, 28, 241–257. [Google Scholar] [CrossRef]
- Bell, S.S.; Woodin, S. Community unity: Experimental evidence for meiofauna and macrofauna. J. Mar. Res. 1984, 42, 605–632. [Google Scholar] [CrossRef]
- Ban, S.M.; Nelson, W.G. Role of Diopatra cuprea Bosc (Polychaeta: Onuphidae) Tubes in Structuring a Subtropical Infaunal Community. Bull. Mar. Sci. 1987, 40, 11–21. [Google Scholar]
- Luckenbach, M.W. Effects of adult infauna on new recruits: Implications for the role of biogenic refuges. J. Exp. Mar. Biol. Ecol. 1987, 105, 197–206. [Google Scholar] [CrossRef]
- Glassom, D.; Branch, G. Impact of predation by greater flamingos Phoenicopterus ruber on the macrofauna of two southern African lagoons. Mar. Ecol. Prog. Ser. 1997, 149, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Luckenbach, M. Settlement and early post-settlement survival in the recruitment of Mulinia lateralis (Bivalvia). Mar. Ecol. Prog. Ser. 1984, 17, 245–250. [Google Scholar] [CrossRef]
- Luckenbach, M.W. Biogenic structure and foraging by five species of shorebirds (Charadrii). Estuar. Coast. Shelf Sci. 1984, 19, 691–696. [Google Scholar] [CrossRef]
- Ambrose, R.F.; Anderson, T.W. Influence of an artificial reef on the surrounding infaunal community. Mar. Biol. 1990, 107, 41–52. [Google Scholar] [CrossRef]
- Bell, S.S.; Coen, L.D. Investigations on epibenthic meiofauna II: Influence of microhabitat and macroalgae on abundance of small invertebrates on Diopatra cuprea (Bosc) (Polychaeta: Onuphidae) tube-caps in Virginia. J. Exp. Mar. Biol. Ecol. 1982, 61, 175–188. [Google Scholar] [CrossRef]
- Byers, J.E.; Gribben, P.; Yeager, C.; Sotka, E.E. Impacts of an abundant introduced ecosystem engineer within mudflats of the southeastern US coast. Biol. Invasions 2012, 14, 2587–2600. [Google Scholar] [CrossRef]
- Dudley, E.C.; Baker, A.J.; Cariello, A.; Leong, K.M.; Ballantine, A. The Occurrence of Living Mollusks on Diopatra Tube-Caps. Veliger 1989, 32, 229–230. [Google Scholar]
- Mangum, C.P.; Cox, C.D. Analysis of the Feeding Response in the Onuphid Polychaete Diopatra cuprea (Bosc). Biol. Bull. 1971, 140, 215–229. [Google Scholar] [CrossRef]
- Watanabe, K.; Tamai, K.; Numachi, K. Feeding-Habits and Distribution Pattern of 2 Forms of Flatfishes (Genus Pleuron-ich-thys) in Kiisuido Channel Waters Off the Pacific Coast of Central Japan. Nippon Suisan Gakkaishi 1992, 58, 2253–2260. [Google Scholar] [CrossRef]
- Bowman, R.E.; Stillwell, C.E.; Michaels, W.L.; Grosslein, M.D. Food of Northwest Atlantic Fishes and Two Common Species of Squid. NOAA Tech. Memo. 2000, NMFS-NE-155. [Google Scholar]
- Steimle, F. Demersal Fish and American Lobster Diets in the Lower Hudson-Raritan Estuary; US Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northeast Region, Northeast Fisheries Science Center: Woods Hole, MA, USA, 2000.
- Paine, R.T. Trophic Relationships of 8 Sympatric Predatory Gastropods. Ecology 1963, 44, 63–73. [Google Scholar] [CrossRef]
- Rosado, M.C.; Bruxelas, A.; Castro, J.J. Ecological Effects of the Commercial Use of Diopatra neapolitana Delle Chiaje, 1841 and Solen marginatus Montagu and Structure and Dynamics of the Macrofaunal Community in a Sand Beach of the Sado Estuary (Portugal). Estud. Biol. E Conserv. Da Nat. 1993, 9, 40. [Google Scholar]
- Pires, A.; Freitas, R.; Quintino, V.; Rodrigues, A. Can Diopatra neapolitana (Annelida: Onuphidae) regenerate body damage caused by bait digging or predation? Estuar. Coast. Shelf Sci. 2012, 110, 36–42. [Google Scholar] [CrossRef]
- Merz, R.A. Textures and traction: How tube-dwelling polychaetes get a leg up. Invertebr. Biol. 2015, 134, 61–77. [Google Scholar] [CrossRef]
- Berke, S.K.; Cruz, V.; Osman, R.W. Sublethal Predation and Regeneration in Two Onuphid Polychaetes: Patterns and Impli-cations. Biol Bull. 2009, 217, 242–252. [Google Scholar] [CrossRef]
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range shifts and species diversity in marine ecosystem engineers: Patterns and predictions for European sedimentary habitats. Glob. Ecol. Biogeogr. 2010, 19, 223–232. [Google Scholar] [CrossRef]
- Mangum, C. Low Temperature Blockage of the Feeding Response in Boreal and Temperate Zone Polychaetes. Estuaries Coasts 1969, 1, 64–65. [Google Scholar] [CrossRef]
- Woodin, S.A.; Wethey, D.; Dubois, S.F. Population structure and spread of the polychaete Diopatra biscayensis along the French Atlantic coast: Human-assisted transport by-passes larval dispersal. Mar. Environ. Res. 2014, 102, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Galaska, M.P.; Wethey, D.S.; Arias, A.; Dubois, S.F.; Halanych, K.M.; Woodin, S.A. The impact of aquaculture on the genetics and distribution of the onuphid annelid Diopatra biscayensis. Ecol. Evol. 2021, 11, 6184–6194. [Google Scholar] [CrossRef]
- Whitney, N.M.; Wanamaker, A.D.; Ummenhofer, C.C.; Johnson, B.J.; Cresswell-Clay, N.; Kreutz, K.J. Rapid 20th century warming reverses 900-year cooling in the Gulf of Maine. Commun. Earth Environ. 2022, 3, 1–15. [Google Scholar] [CrossRef]
- Allen, M.J. Embryological Development of the Polychaetous Annelid, Diopatra cuprea (Bosc). Biol. Bull. 1959, 116, 339–361. [Google Scholar] [CrossRef]
- Lima, F.; Wethey, D. Robolimpets: Measuring intertidal body temperatures using biomimetic loggers. Limnol. Oceanogr. Methods 2009, 7, 347–353. [Google Scholar] [CrossRef]
- Smale, D.; Wernberg, T. Satellite-derived SST data as a proxy for water temperature in nearshore benthic ecology. Mar. Ecol. Prog. Ser. 2009, 387, 27–37. [Google Scholar] [CrossRef]
- Gilman, S.; Hayford, H.; Craig, C.; Carrington, E. Body temperatures of an intertidal barnacle and two whelk predators in relation to shore height, solar aspect, and microhabitat. Mar. Ecol. Prog. Ser. 2015, 536, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Judge, R.; Choi, F.; Helmuth, B. Recent Advances in Data Logging for Intertidal Ecology. Front. Ecol. Evol. 2018, 6, 213. [Google Scholar] [CrossRef] [Green Version]
- Wethey, D.S.; Woodin, S.A. Climate change and Arenicola marina: Heat waves and the southern limit of an ecosystem engineer. Estuar. Coast. Shelf Sci. 2022, 276, 10801. [Google Scholar] [CrossRef]
- Wethey, D.S.; Brin, L.D.; Helmuth, B.; Mislan, K. Predicting intertidal organism temperatures with modified land surface models. Ecol. Model. 2011, 222, 3568–3576. [Google Scholar] [CrossRef]
- Macho, G.; Woodin, S.A.; Wethey, D.S.; Vázquez, E. Impacts of Sublethal and Lethal High Temperatures on Clams Exploited in European Fisheries. J. Shellfish Res. 2016, 35, 405–419. [Google Scholar] [CrossRef]
- Freitas, F.; Cunha, T.; Hall, A.; Queiroga, H. Diopatra Neapolitana, Importância Sócio-Económica e Sustentabilidade Das Capturas, No Canal de Mira, Ria de Aveiro; Actas das Jornadas da Ria de Aveiro: Aveiro, Portugal, 2011; p. 66.
- Aleixo, A.; Queiroga, H.; Xenarios, S.; Lillebø, A. Catch Estimates and Bioecomomic Analysis of Bait Digging: The Case of the Tube Worm Diopatra neapolitana. Bioforsk Rep. 2014, 9, 1–32. [Google Scholar]
- Gurgel, C.F.D.; Norris, J.N.; Schmidt, W.E.; Le, H.N.; Fredericq, S. Systematics of the Gracilariales (Rhodophyta) including new subfamilies, tribes, subgenera, and two new genera, Agarophyton gen. nov. and Crassa gen. nov. Phytotaxa 2018, 374, 1–23. [Google Scholar]
- Kollars, N.; Byers, J.; Sotka, E. Invasive décor: An association between a native decorator worm and a non-native seaweed can be mutualistic. Mar. Ecol. Prog. Ser. 2016, 545, 135–145. [Google Scholar] [CrossRef]

| Study | Species | Location | Community Type | Densities Tested | Effects |
|---|---|---|---|---|---|
| Mangum et al., 1968 [31] | D. cuprea | Chesapeake Bay, Virginia, USA | Epifauna | Single tubes | Tubes support 49 species of Arthropoda, Mollusca, Annelida, and other invertebrate phyla. |
| Bell & Coen 1982 [9] | D. cuprea | Tampa Bay, FL | Meiofauna | Single tubes | Tubes support crustacean nauplii, copepods, and nematodes (tens to hundreds per tube). Polychaetes also found on tubes. |
| Bell & Coen 1982 [72] | D. cuprea | Tom’s Cove, Virginia, USA | Meiofauna | 1 or 4 · 0.01 m−2 | Tubes support nauplii, copepods, amphipods, ostracods, nematodes, and polychaetes. Abundances increase when algae are also present, with no effect of tube density. |
| Dudley et al., 1989 [74] | D. ornata | Venado Beach, Panama | Epifaunal mollusca | Single tubes | Tubes support seven species of gastropod, four bivalves, and a chiton. |
| Dudley et al., 1989 [74] | D. cuprea | Virginia, USA and Woods Hole, Massachusetts, USA | Epifaunal mollusca | Single tubes | Tubes support five species of gastropod and four bivalves. |
| Diaz et al., 2003 [10] | D. cuprea | Mid-Atlantic Bight | Juvenile epibenthic fish | “mat” | Juvenile fish were twice as abundant in tube mats as bare sand during the day (reversed at night). Eight species of fish were associated with D. cuprea tubes. |
| Thomsen et al., 2011 [23] | Diopatra-like onuphid * | Inhaca Island, Mozambique | Epifauna | Single tubes | ↑ richness and abundance on single tubes |
| Santos and Aviz 2018 [20] † | D. cuprea * | Algodoal-Maiandeua, Brazil | Epifauna and Infauna | Single tubes | Seasonal ↑ richness and abundance |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berke, S.K. A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology 2022, 11, 1485. https://doi.org/10.3390/biology11101485
Berke SK. A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology. 2022; 11(10):1485. https://doi.org/10.3390/biology11101485
Chicago/Turabian StyleBerke, Sarah K. 2022. "A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete" Biology 11, no. 10: 1485. https://doi.org/10.3390/biology11101485
APA StyleBerke, S. K. (2022). A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology, 11(10), 1485. https://doi.org/10.3390/biology11101485