
Antarctic Seabed Assemblages in an Ice-Shelf-Adjacent Polynya, Western Weddell Sea

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
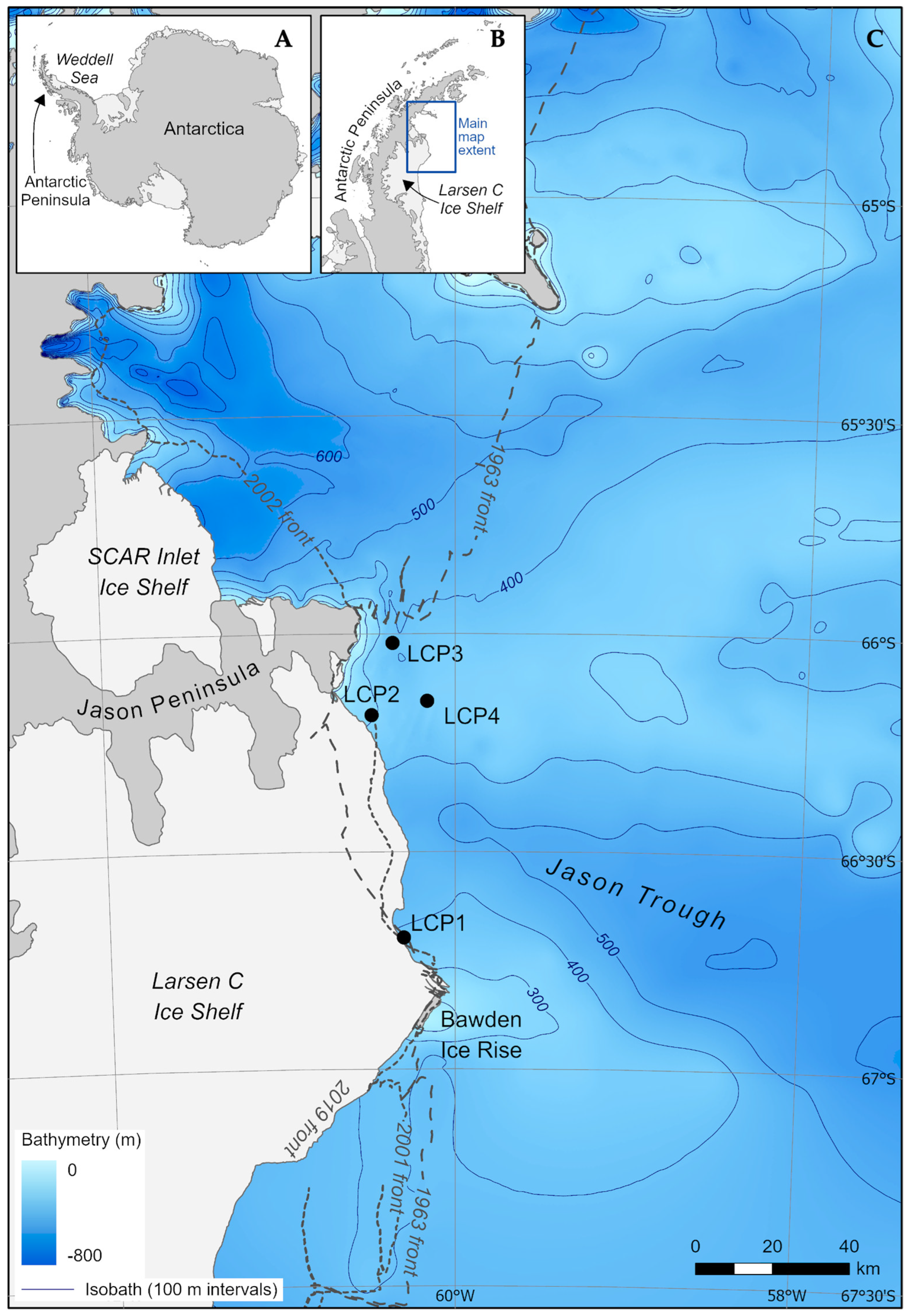
2.1. Study Area and Sites
2.2. Collation of Environmental Data Used in the Study
2.3. Seafloor Imagery Collection
2.4. Post-Processing of Collected Imagery
2.5. Frame Analysis and Assignments
2.6. Statistical Analyses to Explore Biotic Differences and Potential Environmental Drivers
3. Results
3.1. Seafloor Habitat of the Study Area—Observations and Features
3.2. Environmental Differences Determined from Remotely-Sensed Data
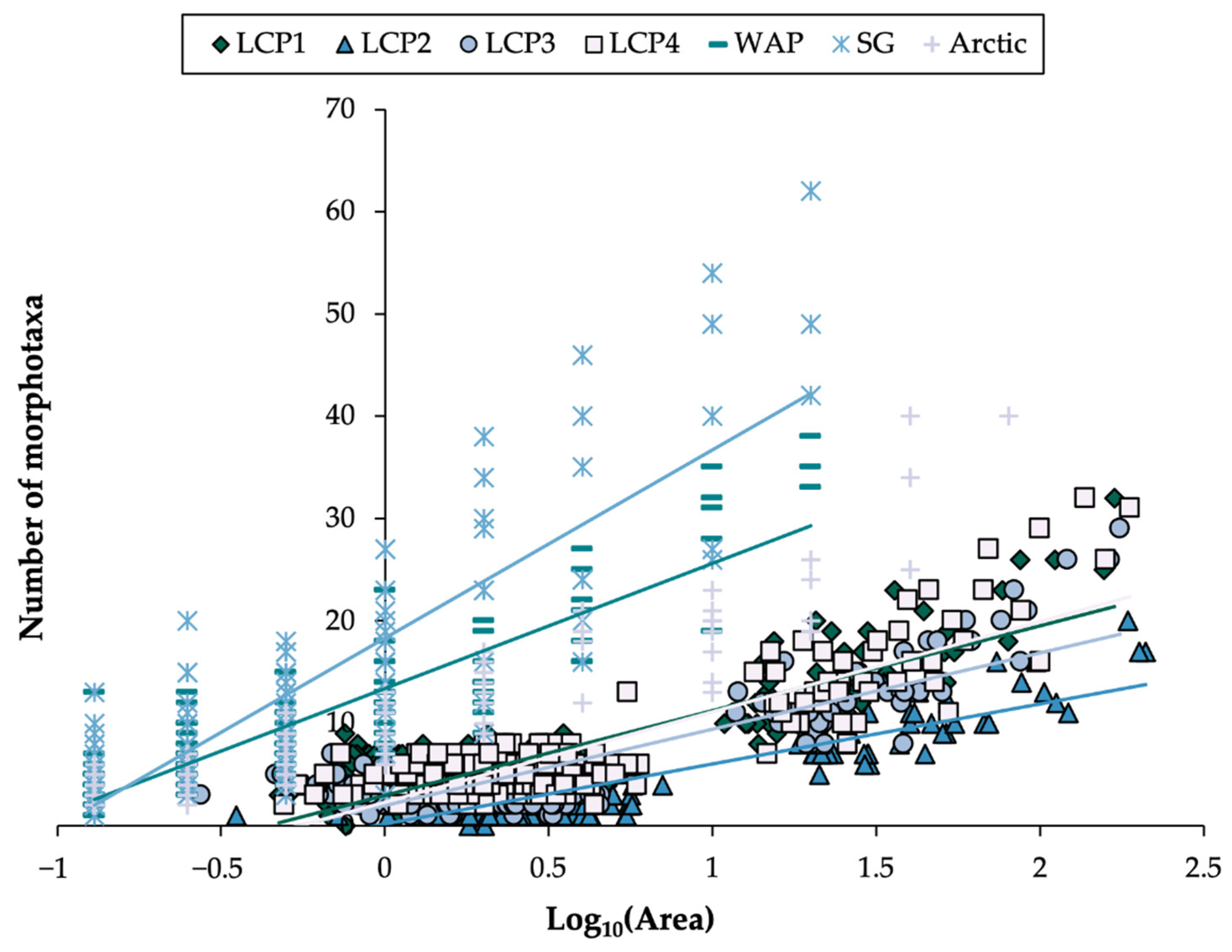
3.3. Megafaunal Richness: Accumulation with Area, and Pattern with Ice-Shelf Proximity and Substratum Hardness
3.4. Megafaunal Density and Pattern with Ice-Shelf Proximity and Substratum Hardness
3.5. Functional Group Richness and Density Patterns with Ice-Shelf Proximity and Substratum Hardness, and Potential Surrogacy of Wider Faunal Patterns
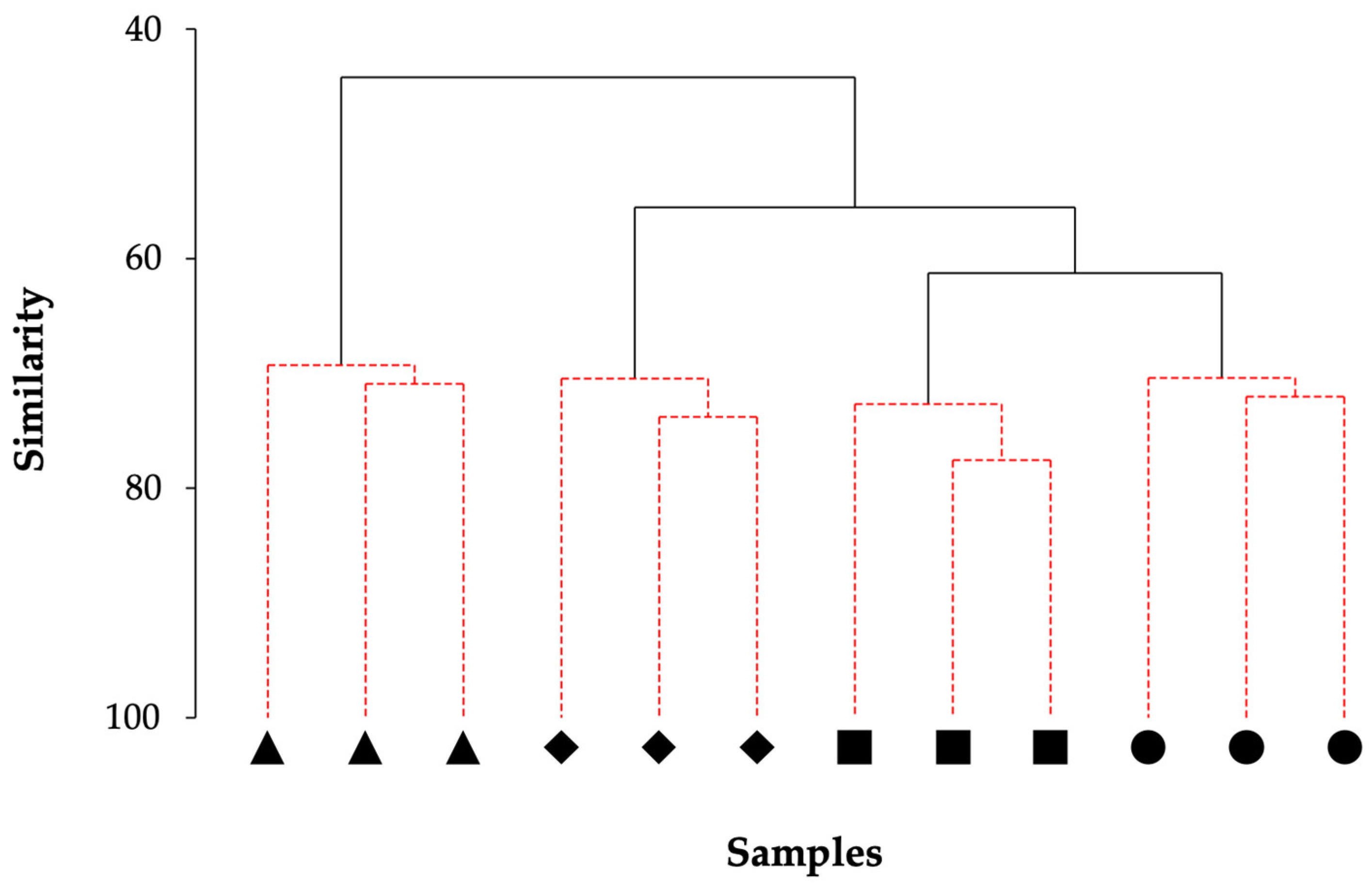
3.6. Megafaunal Assemblage Composition
3.7. Environmental Differences between Study Sites
3.8. Environmental Influences on Megafaunal Assemblage Composition Differences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arntz, W.E.; Brey, T.; Gallardo, V.A. Antarctic Zoobenthos. Oceanogr. Mar. Biol. 1994, 32, 241–304. [Google Scholar]
- Gutt, J.; Isla, E.; Xavier, J.C.; Adams, B.J.; Ahn, I.Y.; Cheng, C.H.C.; Colesie, C.; Cummings, V.J.; di Prisco, G.; Griffiths, H.; et al. Antarctic ecosystems in transition—Life between stresses and opportunities. Biol. Rev. 2021, 96, 798–821. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.F.; Filippelli, G.M.; Florindo, F.; Martin, E.E.; Scher, H.D. Onset and role of the Antarctic Circumpolar Current. Deep Sea Res. II 2007, 54, 2388–2398. [Google Scholar] [CrossRef] [Green Version]
- Dayton, P.; Jarrell, S.; Kim, S.; Thrush, S.; Hammerstrom, K.; Slattery, M.; Parnell, E. Surprising episodic recruitment and growth of Antarctic sponges: Implications for ecological resilience. J. Exp. Mar. Biol. Ecol. 2016, 482, 38–55. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Comiso, J. Solve Antarctica’s sea-ice puzzle. Nature 2017, 547, 275–277. [Google Scholar] [CrossRef]
- Rogers, A.D.; Frinault, B.A.V.; Barnes, D.K.A.; Bindoff, N.L.; Downie, R.; Ducklow, H.W.; Friedlaender, A.S.; Hart, T.; Hill, S.L.; Hofmann, E.E.; et al. Antarctic Futures: An Assessment of Climate-Driven Changes in Ecosystem Structure, Function, and Service Provisioning in the Southern Ocean. Annu. Rev. Mar. Sci. 2020, 12, 87–120. [Google Scholar] [CrossRef] [Green Version]
- Scambos, T.; Hulbe, C.; Fahnestock, M. Climate-Induced Ice Shelf Disintegration in the Antarctic Peninsula. In Antarctic Peninsula Climate Variability: Historical and Paleoenvironmental Perspectives; Domack, E., Levente, A., Burnet, A., Bindschadler, R., Convey, P., Kirby, M., Eds.; Antarctic Research Series; American Geophysical Union: Washington, DC, USA, 2003; Volume 79, pp. 79–92. [Google Scholar]
- Scambos, T.; Ross, R.; Bauer, R.; Yermolin, Y.; Skvarca, P.; Long, D.; Bohlander, J.; Haran, T. Calving and ice-shelf break-up processes investigated by proxy: Antarctic tabular iceberg evolution during northward drift. J. Glaciol. 2008, 54, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Cook, A.J.; Vaughan, D.G. Overview of areal changes of the ice shelves on the Antarctic Peninsula over the past 50 years. Cryosphere 2010, 4, 77–98. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.A.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H.; et al. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef] [Green Version]
- Cook, A.J.; Holland, P.R.; Meredith, M.P.; Murray, T.; Luckman, A.; Vaughan, D.G. Ocean forcing of glacier retreat in the western Antarctic Peninsula. Science 2016, 353, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Eayrs, C.; Li, X.; Raphael, M.N.; Holland, D.M. Rapid decline in Antarctic sea ice in recent years hints at future change. Nat. Geosci. 2021, 14, 460–464. [Google Scholar] [CrossRef]
- Christie, F.D.W.; Benham, T.J.; Batchelor, C.L.; Rack, W.; Montelli, A.; Dowdeswell, J.A. Antarctic ice-shelf advance driven by anomalous atmospheric and sea-ice circulation. Nat. Geosci. 2022, 15, 356–362. [Google Scholar] [CrossRef]
- Gutt, J.; Arndt, J.; Kraan, C.; Dorschel, B.; Schröder, M.; Bracher, A.; Piepenburg, D. Benthic communities and their drivers: A spatial analysis off the Antarctic Peninsula. Limnol. Oceanogr. 2019, 64, 2341–2357. [Google Scholar] [CrossRef] [Green Version]
- Vause, B.J.; Morley, S.A.; Fonseca, V.G.; Jazdzewska, A.; Ashton, G.V.; Barnes, D.K.A.; Giebner, H.; Clark, M.S.; Peck, L.S. Spatial and temporal dynamics of Antarctic shallow soft-bottom benthic communities: Ecological drivers under climate change. BMC Ecol. 2019, 19, 27. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, R.D.; Melbourne-Thomas, J.; Grant, S.M.; Barnes, D.K.A.; Hughes, K.A.; Halfter, S.; Meredith, M.P.; Murphy, E.J.; Trebilco, R.; Hill, S.L. Future Risk for Southern Ocean Ecosystem Services Under Climate Change. Front. Mar. Sci. 2021, 7, 615214. [Google Scholar] [CrossRef]
- Grant, S.M.; Waller, C.L.; Morley, S.A.; Barnes, D.K.A.; Brasier, M.J.; Double, M.C.; Griffiths, H.J.; Hughes, K.A.; Jackson, J.A.; Waluda, C.M.; et al. Local Drivers of Change in Southern Ocean Ecosystems: Human Activities and Policy Implications. Front. Ecol. Evol. 2021, 9, 26. [Google Scholar] [CrossRef]
- De Broyer, C.; Koubbi, P.; Griffiths, H.; Raymond, B.; d’, U.; Van de Putte, A.; Danis, B.; David, B.; Grant, S.; Gutt, J.; et al. Biogeographic Atlas of the Southern Ocean; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; p. 510. [Google Scholar]
- Henley, S.F.; Cavan, E.L.; Fawcett, S.E.; Kerr, R.; Monteiro, T.; Sherrell, R.M.; Bowie, A.R.; Boyd, P.W.; Barnes, D.K.A.; Schloss, I.R.; et al. Changing Biogeochemistry of the Southern Ocean and Its Ecosystem Implications. Front. Mar. Sci. 2020, 7, 581. [Google Scholar] [CrossRef]
- Peck, L.S.; Barnes, D.K.A.; Cook, A.J.; Fleming, A.H.; Clarke, A. Negative feedback in the cold: Ice retreat produces new carbon sinks in Antarctica. Glob. Chang. Biol. 2010, 16, 2614–2623. [Google Scholar] [CrossRef]
- Barnes, D.K.A. Polar zoobenthos blue carbon storage increases with sea ice losses, because across-shelf growth gains from longer algal blooms outweigh ice scour mortality in the shallows. Glob. Chang. Biol. 2017, 23, 5083–5091. [Google Scholar] [CrossRef]
- Barnes, D.K.A. Blue Carbon on Polar and Subpolar Seabeds. In Carbon Capture, Utilization and Sequestration; Agarwal, R.K., Ed.; IntechOpen: London, UK, 2018; pp. 37–56. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.K.A.; Fleming, A.; Sands, C.J.; Quartino, M.L.; Deregibus, D. Icebergs, sea ice, blue carbon and Antarctic climate feedbacks. Philos. Trans. R. Soc. A 2018, 376, 20170176. [Google Scholar] [CrossRef]
- Matschiner, M.; Colombo, M.; Damerau, M.; Ceballos, S.; Hanel, R.; Salzburger, W. The Adaptive Radiation of Notothenioid Fishes in the Waters of Antarctica. In Extremophile Fishes: Ecology, Evolution, and Physiology of Teleosts in Extreme Environments; Riesch, R., Tobler, M., Plath, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 35–57. [Google Scholar] [CrossRef]
- Aronson, R.B.; Thatje, S.; Clarke, A.; Peck, L.S.; Blake, D.B.; Wilga, C.D.; Seibel, B.A. Climate Change and Invasibility of the Antarctic Benthos. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 129–154. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.; Johnston, N.M. Antarctic marine benthic diversity. Oceanogr. Mar. Biol. 2003, 41, 47–114. [Google Scholar] [CrossRef]
- Ashton, G.V.; Morley, S.A.; Barnes, D.K.A.; Clark, M.S.; Peck, L.S. Warming by 1 degrees C Drives Species and Assemblage Level Responses in Antarctica’s Marine Shallows. Curr. Biol. 2017, 27, 2698–2705.e2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, H.J.; Meijers, A.J.S.; Bracegirdle, T.J. More losers than winners in a century of future Southern Ocean seafloor warming. Nat. Clim. Chang. 2017, 7, 749–754. [Google Scholar] [CrossRef]
- Morley, S.A.; Barnes, D.K.A.; Dunn, M.J. Predicting Which Species Succeed in Climate-Forced Polar Seas. Front. Mar. Sci. 2019, 5, 507. [Google Scholar] [CrossRef]
- Rignot, E.; Jacobs, S.; Mouginot, J.; Scheuchl, B. Ice-Shelf Melting Around Antarctica. Science 2013, 341, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Fürst, J.J.; Durand, G.; Gillet-Chaulet, F.; Tavard, L.; Rankl, M.; Braun, M.; Gagliardini, O. The safety band of Antarctic ice shelves. Nat. Clim. Chang. 2016, 6, 479–482. [Google Scholar] [CrossRef]
- Hogg, A.E.; Gudmundsson, G.H. Impacts of the Larsen-C Ice Shelf calving event. Nat. Clim. Chang. 2017, 7, 540–542. [Google Scholar] [CrossRef]
- Barnes, D.K.A.; Kuhn, G.; Hillenbrand, C.D.; Gromig, R.; Koglin, N.; Biskaborn, B.K.; Frinault, B.A.V.; Klages, J.P.; Smith, E.C.; Berger, S.; et al. Richness, growth, and persistence of life under an Antarctic ice shelf. Curr. Biol. 2021, 31, R1566–R1567. [Google Scholar] [CrossRef]
- Griffiths, H.J.; Anker, P.; Linse, K.; Maxwell, J.; Post, A.L.; Stevens, C.; Tulaczyk, S.; Smith, J.A. Breaking All the Rules: The First Recorded Hard Substrate Sessile Benthic Community Far Beneath an Antarctic Ice Shelf. Front. Mar. Sci. 2021, 8, 642040. [Google Scholar] [CrossRef]
- Domack, E.; Ishman, S.; Leventer, A.; Sylva, S.; Willmott, V.; Huber, B. A chemotrophic ecosystem found beneath Antarctic Ice Shelf. Eos 2005, 86, 269–272. [Google Scholar] [CrossRef] [Green Version]
- Riddle, M.J.; Craven, M.; Goldsworthy, P.M.; Carsey, F. A diverse benthic assemblage 100 km from open water under the Amery Ice Shelf, Antarctica. Paleoceanography 2007, 22, PA1204. [Google Scholar] [CrossRef]
- Post, A.L.; Galton-Fenzi, B.K.; Riddle, M.J.; Herraiz-Borreguero, L.; O’Brien, P.E.; Hemer, M.A.; McMinn, A.; Rasch, D.; Craven, M. Modern sedimentation, circulation and life beneath the Amery Ice Shelf, East Antarctica. Cont. Shelf Res. 2014, 74, 77–87. [Google Scholar] [CrossRef]
- Kim, S. Complex life under the McMurdo Ice Shelf, and some speculations on food webs. Antarct. Sci. 2019, 31, 80–88. [Google Scholar] [CrossRef]
- Dowdeswell, J.A.; Batchelor, C.L.; Dorschel, B.; Benham, T.J.; Christie, F.D.W.; Dowdeswell, E.K.; Montelli, A.; Arndt, J.E.; Gebhardt, C. Sea-floor and sea-ice conditions in the western Weddell Sea, Antarctica, around the wreck of Sir Ernest Shackleton’s Endurance. Antarct. Sci. 2020, 32, 301–313. [Google Scholar] [CrossRef]
- Convey, P.; Bindschadler, R.; di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.A.; Mayewski, P.A.; Summerhayes, C.P.; Turner, J. Antarctic climate change and the environment. Antarct. Sci. 2009, 21, 541–563. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, H.J. Antarctic Marine Biodiversity—What Do We Know About the Distribution of Life in the Southern Ocean? PLoS ONE 2010, 5, e11683. [Google Scholar] [CrossRef] [Green Version]
- Gutt, J.; Starmans, A. Structure and biodiversity of megabenthos in the Weddell and Lazarev Seas (Antarctica): Ecological role of physical parameters and biological interactions. Polar Biol. 1998, 20, 229–247. [Google Scholar] [CrossRef]
- Purser, A.; Hehemann, L.; Boehringer, L.; Tippenhauer, S.; Wege, M.; Bornemann, H.; Pineda-Metz, S.E.A.; Flintrop, C.M.; Koch, F.; Hellmer, H.H.; et al. A vast icefish breeding colony discovered in the Antarctic. Curr. Biol. 2022, 32, 842–850.e844. [Google Scholar] [CrossRef]
- Barnes, D.K.A. Iceberg killing fields limit huge potential for benthic blue carbon in Antarctic shallows. Glob. Chang. Biol. 2017, 23, 2649–2659. [Google Scholar] [CrossRef] [Green Version]
- Sahade, R.; Lagger, C.; Torre, L.; Momo, F.; Monien, P.; Schloss, I.; Barnes, D.K.; Servetto, N.; Tarantelli, S.; Tatian, M.; et al. Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Sci. Adv. 2015, 1, e1500050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, L.S.; Brockington, S.; Vanhove, S.; Beghyn, M. Community recovery following catastrophic iceberg impacts in a soft-sediment shallow-water site at Signy Island, Antarctica. Mar. Ecol. Prog. Ser. 1999, 186, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gutt, J. On the direct impact of ice on marine benthic communities, a review. Polar Biol. 2001, 24, 553–564. [Google Scholar] [CrossRef]
- Gutt, J.; Starmans, A. Quantification of iceberg impact and benthic recolonisation patterns in the Weddell Sea (Antarctica). Polar Biol. 2001, 24, 615–619. [Google Scholar] [CrossRef]
- Gutt, J.; Piepenburg, D. Scale-dependent impact on diversity of Antarctic benthos caused by grounding of icebergs. Mar. Ecol. Prog. Ser. 2003, 253, 77–83. [Google Scholar] [CrossRef]
- Smale, D.A.; Barnes, D.K.A.; Fraser, K.P.P.; Peck, L.S. Benthic community response to iceberg scouring at an intensely disturbed shallow water site at Adelaide Island, Antarctica. Mar. Ecol. Prog. Ser. 2008, 355, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Robinson, B.J.O.; Barnes, D.K.A.; Grange, L.J.; Morley, S.A. The Extremes of Disturbance Reduce Functional Redundancy: Functional Trait Assessment of the Shallow Antarctic Benthos. Front. Mar. Sci. 2022, 8, 797112. [Google Scholar] [CrossRef]
- Bertolin, M.L.; Schloss, I.R. Phytoplankton production after the collapse of the Larsen A Ice Shelf, Antarctica. Polar Biol. 2009, 32, 1435–1446. [Google Scholar] [CrossRef]
- Sañé, E.; Isla, E.; Grémare, A.; Gutt, J.; Vétion, G.; DeMaster, D.J. Pigments in sediments beneath recently collapsed ice shelves: The case of Larsen A and B shelves, Antarctic Peninsula. J. Sea Res. 2011, 65, 94–102. [Google Scholar] [CrossRef]
- Gutt, J.; Cape, M.; Dimmler, W.; Fillinger, L.; Isla, E.; Lieb, V.; Lundälv, T.; Pulcher, C. Shifts in Antarctic megabenthic structure after ice-shelf disintegration in the Larsen area east of the Antarctic Peninsula. Polar Biol. 2013, 36, 895–906. [Google Scholar] [CrossRef]
- Domack, E.; Duran, D.; Leventer, A.; Ishman, S.; Doane, S.; McCallum, S.; Amblas, D.; Ring, J.; Gilbert, R.; Prentice, M. Stability of the Larsen B ice shelf on the Antarctic Peninsula during the Holocene epoch. Nature 2005, 436, 681–685. [Google Scholar] [CrossRef]
- Fillinger, L.; Janussen, D.; Lundalv, T.; Richter, C. Rapid glass sponge expansion after climate-induced Antarctic ice shelf collapse. Curr. Biol. 2013, 23, 1330–1334. [Google Scholar] [CrossRef] [Green Version]
- Zwerschke, N.; Morley, S.A.; Peck, L.S.; Barnes, D.K.A. Can Antarctica’s shallow zoobenthos ‘bounce back’ from iceberg scouring impacts driven by climate change? Glob. Chang. Biol. 2021, 27, 3157–3165. [Google Scholar] [CrossRef]
- Bowden, D.A.; Clarke, A.; Peck, L.S.; Barnes, D.K.A. Antarctic sessile marine benthos: Colonisation and growth on artificial substrata over three years. Mar. Ecol. Prog. Ser. 2006, 316, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dayton, P.K.; Kim, S.; Jarrell, S.C.; Oliver, J.S.; Hammerstrom, K.; Fisher, J.L.; O’Connor, K.; Barber, J.S.; Robilliard, G.; Barry, J.; et al. Recruitment, Growth and Mortality of an Antarctic Hexactinellid Sponge, Anoxycalyx joubini. PLoS ONE 2013, 8, e56939. [Google Scholar] [CrossRef] [Green Version]
- Teixidó, N.; Garrabou, J.; Gutt, J.; Arntz, W.E. Recovery in Antarctic benthos after iceberg disturbance: Trends in benthic composition, abundance and growth forms. Mar. Ecol. Prog. Ser. 2004, 278, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Teixidó, N.; Garrabou, J.; Gutt, J.; Arntz, W.E. Iceberg Disturbance and Successional Spatial Patterns: The Case of the Shelf Antarctic Benthic Communities. Ecosystems 2007, 10, 143–158. [Google Scholar] [CrossRef]
- Gerdes, D.; Isla, E.; Knust, R.; Mintenbeck, K.; Rossi, S. Response of Antarctic benthic communities to disturbance: First results from the artificial Benthic Disturbance Experiment on the eastern Weddell Sea Shelf, Antarctica. Polar Biol. 2008, 31, 1469–1480. [Google Scholar] [CrossRef]
- Wellner, J.; Scambos, T.; Domack, E.; Vernet, M.; Leventer, A.; Balco, G.; Brachfeld, S.; Cape, M.; Huber, B.; Ishman, S.; et al. The Larsen Ice Shelf System, Antarctica (LARISSA): Polar Systems Bound Together, Changing Fast. GSA Today 2019, 29, 4–10. [Google Scholar] [CrossRef]
- Gilbert, E.; Kittel, C. Surface Melt and Runoff on Antarctic Ice Shelves at 1.5 °C, 2 °C, and 4 °C of Future Warming. Geophys. Res. Lett. 2021, 48, e2020GL091733. [Google Scholar] [CrossRef]
- Bax, N.; Sands, C.J.; Gogarty, B.; Downey, R.V.; Moreau, C.V.E.; Moreno, B.; Held, C.; Paulsen, M.L.; McGee, J.; Haward, M.; et al. Perspective: Increasing blue carbon around Antarctica is an ecosystem service of considerable societal and economic value worth protecting. Glob. Chang. Biol. 2021, 27, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Gutt, J.; Barratt, I.; Domack, E.; d’Udekem d’Acoz, C.; Dimmler, W.; Grémare, A.; Heilmayer, O.; Isla, E.; Janussen, D.; Jorgensen, E.; et al. Biodiversity change after climate-induced ice-shelf collapse in the Antarctic. Deep Sea Res. II 2011, 58, 74–83. [Google Scholar] [CrossRef]
- Hauquier, F.; Ingels, J.; Gutt, J.; Raes, M.; Vanreusel, A. Characterisation of the Nematode Community of a Low-Activity Cold Seep in the Recently Ice-Shelf Free Larsen B Area, Eastern Antarctic Peninsula. PLoS ONE 2011, 6, e22240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almond, P.M.; Linse, K.; Dreutter, S.; Grant, S.M.; Griffiths, H.J.; Whittle, R.J.; Mackenzie, M.; Reid, W.D.K. In-situ Image Analysis of Habitat Heterogeneity and Benthic Biodiversity in the Prince Gustav Channel, Eastern Antarctic Peninsula. Front. Mar. Sci. 2021, 8, 18. [Google Scholar] [CrossRef]
- Drennan, R.; Dahlgren, T.G.; Linse, K.; Glover, A.G. Annelid Fauna of the Prince Gustav Channel, a Previously Ice-Covered Seaway on the Northeastern Antarctic Peninsula. Front. Mar. Sci. 2021, 7, 595303. [Google Scholar] [CrossRef]
- Brooks, C.M.; Chown, S.L.; Douglass, L.L.; Raymond, B.P.; Shaw, J.D.; Sylvester, Z.T.; Torrens, C.L. Progress towards a representative network of Southern Ocean protected areas. PLoS ONE 2020, 15, e0231361. [Google Scholar] [CrossRef] [Green Version]
- Teschke, K.; Brtnik, P.; Hain, S.; Herata, H.; Liebschner, A.; Pehlke, H.; Brey, T. Planning marine protected areas under the CCAMLR regime—The case of the Weddell Sea (Antarctica). Mar. Policy 2021, 124, 104370. [Google Scholar] [CrossRef]
- Knox, G.A. Biology of the Southern Ocean, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006; p. 640. [Google Scholar] [CrossRef]
- Vernet, M.; Geibert, W.; Hoppema, M.; Brown, P.J.; Haas, C.; Hellmer, H.H.; Jokat, W.; Jullion, L.; Mazloff, M.; Bakker, D.C.E.; et al. The Weddell Gyre, Southern Ocean: Present Knowledge and Future Challenges. Rev. Geophys. 2019, 57, 623–708. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L.; Strong, A.L. Environmental controls of marine productivity hot spots around Antarctica. J. Geophys. Res. Oceans 2015, 120, 5545–5565. [Google Scholar] [CrossRef]
- Dowdeswell, J.; Shears, J.; Batchelor, C.; Christie, F.; Rack, W.; Montelli, A.; Evans, J.; Dowdeswell, E.; Ottesen, D.; Fawcett, S.; et al. The Weddell Sea Expedition 2019: Cruise Scientific Report; Cambridge University: Cambridge, UK, 2019; p. 151. [Google Scholar]
- Cofaigh, C.Ó.; Davies, B.J.; Livingstone, S.J.; Smith, J.A.; Johnson, J.S.; Hocking, E.P.; Hodgson, D.A.; Anderson, J.B.; Bentley, M.J.; Canals, M.; et al. Reconstruction of ice-sheet changes in the Antarctic Peninsula since the Last Glacial Maximum. Quat. Sci. Rev. 2014, 100, 87–110. [Google Scholar] [CrossRef] [Green Version]
- Batchelor, C.L.; Montelli, A.; Ottesen, D.; Evans, J.; Dowdeswell, E.K.; Christie, F.D.W.; Dowdeswell, J.A. New insights into the formation of submarine glacial landforms from high-resolution Autonomous Underwater Vehicle data. Geomorphology 2020, 370, 107396. [Google Scholar] [CrossRef]
- Batchelor, C.L.; Frinault, B.A.V.; Christie, F.D.W.; Montelli, A.; Dowdeswell, J.A. The morphology of pockmarks on the north-east Antarctic Peninsula continental shelf. Antarct. Sci. 2022, 34, 313–324. [Google Scholar] [CrossRef]
- Dorschel, B.; Hehemann, L.; Viquerat, S.; Warnke, F.; Dreutter, S.; Schulze Tenberge, Y.; Accettella, D.; An, L.; Barrios, F.; Bazhenova, E.A.; et al. The International Bathymetric Chart of the Southern Ocean Version 2 (IBCSO v2). Sci. Data 2022, 9, 275. [Google Scholar] [CrossRef]
- Cook, A.; Fox, A.; Thomson, J. Coastal Change Data for the Antarctic Peninsula Region, 1843 to 2008 [Dataset], 1st ed.; Polar Data Centre, Natural Environment Research Council, UK Research & Innovation: Cambridge, UK, 2021. [Google Scholar] [CrossRef]
- Christie, F.; Benham, T.; Batchelor, C.L.; Rack, W.; Montelli, A.; Dowdeswell, J. Antarctic Ice Front Positions, 1979–2021, Supporting “Antarctic Ice-Shelf Advance Driven by Anomalous Atmospheric and Sea-Ice Circulation” [Dataset]; Cambridge Apollo Data Repository; University of Cambridge: Cambridge, UK, 2022. [Google Scholar] [CrossRef]
- Galeron, J.; Herman, R.L.; Arnaud, P.M.; Arntz, W.E.; Hain, S.; Klages, M. Macrofaunal communities on the continental shelf and slope of the southeastern Weddell Sea, Antarctica. Polar Biol. 1992, 12, 283–290. [Google Scholar] [CrossRef]
- Kim, D.-U.; Khim, J.S.; Ahn, I.-Y. Patterns, drivers and implications of ascidian distributions in a rapidly deglaciating fjord, King George Island, West Antarctic Peninsula. Ecol. Indic. 2021, 125, 107467. [Google Scholar] [CrossRef]
- Cook, A.J.; Fox, A.J.; Vaughan, D.G.; Ferrigno, J.G. Retreating glacier fronts on the Antarctic Peninsula over the past half-century. Science 2005, 308, 541–544. [Google Scholar] [CrossRef]
- Ferrigno, J.G.; Foley, K.M.; Swithinbank, C.; Williams, R.S., Jr.; Dalide, L.M. Coastal-Change and Glaciological Map of the Ronne Ice Shelf Area, Antarctica, 1974–2002; IMAP 2600-D; U.S. Geological Survey Publications Warehouse: Reston, VA, USA, 2005; p. 11.
- Haran, T.; Bohlander, J.; Scambos, T.; Painter, T.; Fahnestock, M. MODIS Mosaic of Antarctica 2003-2004 (MOA2004) Image Map, Version 2 [Dataset]; NASA National Snow and Ice Data Center Distributed Active Archive Center: Boulder, CO, USA, 2012. [CrossRef]
- Wuite, J.; Rott, H.; Hetzenecker, M.; Floricioiu, D.; De Rydt, J.; Gudmundsson, G.H.; Nagler, T.; Kern, M. Evolution of surface velocities and ice discharge of Larsen B outlet glaciers from 1995 to 2013. Cryosphere 2015, 9, 957–969. [Google Scholar] [CrossRef] [Green Version]
- Cavalieri, D.J.; Parkinson, C.L.; Gloersen, P.; Zwally, H.J. Sea Ice Concentrations from Nimbus-7 SMMR and DMSP SSM/I-SSMIS Passive Microwave Data, Version 1 (NASA National Snow and Ice Data Center Distributed Active Archive Center, 1996, Updated 2020) [Dataset]; NASA National Snow and Ice Data Center Distributed Active Archive Center: Boulder, CO, USA, 2022. [CrossRef]
- Arrigo, K.R.; van Dijken, G.L.; Bushinsky, S. Primary production in the Southern Ocean, 1997–2006. J. Geophys. Res. 2008, 113, C08004. [Google Scholar] [CrossRef]
- Hutchinson, K.; Deshayes, J.; Sallee, J.B.; Dowdeswell, J.A.; Lavergne, C.; Ansorge, I.; Luyt, H.; Henry, T.; Fawcett, S.E. Water Mass Characteristics and Distribution Adjacent to Larsen C Ice Shelf, Antarctica. J. Geophys. Res. Oceans 2020, 125, e2019JC015855. [Google Scholar] [CrossRef]
- Flynn, R.F.; Bornman, T.G.; Burger, J.M.; Smith, S.; Spence, K.A.M.; Fawcett, S.E. Summertime productivity and carbon export potential in the Weddell Sea, with a focus on the waters adjacent to Larsen C Ice Shelf. Biogeosciences 2021, 18, 6031–6059. [Google Scholar] [CrossRef]
- Jones, D.O.B.; Bett, B.J.; Wynn, R.B.; Masson, D.G. The use of towed camera platforms in deep-water science. Underw. Technol. 2009, 28, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Langenkämper, D.; Zurowietz, M.; Schoening, T.; Nattkemper, T.W. BIIGLE 2.0—Browsing and Annotating Large Marine Image Collections. Front. Mar. Sci. 2017, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.K.A.; Sands, C.J. Functional group diversity is key to Southern Ocean benthic carbon pathways. PLoS ONE 2017, 12, e0179735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, D.K.A.; Sands, C.J.; Cook, A.; Howard, F.; Roman Gonzalez, A.; Munoz-Ramirez, C.; Retallick, K.; Scourse, J.; Van Landeghem, K.; Zwerschke, N. Blue carbon gains from glacial retreat along Antarctic fjords: What should we expect? Glob. Chang. Biol. 2020, 26, 2750–2755. [Google Scholar] [CrossRef] [PubMed]
- Hogg, O.T.; Huvenne, V.A.I.; Griffiths, H.J.; Linse, K. On the ecological relevance of landscape mapping and its application in the spatial planning of very large marine protected areas. Sci. Total Environ. 2018, 626, 384–398. [Google Scholar] [CrossRef]
- Hogg, O.T.; Downie, A.-L.; Vieira, R.P.; Darby, C. Macrobenthic Assessment of the South Sandwich Islands Reveals a Biogeographically Distinct Polar Archipelago. Front. Mar. Sci. 2021, 8, 650241. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2015; p. 296. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; p. 214. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014; p. 262. [Google Scholar]
- Clarke, K.R.; Green, R.H. Statistical Design And Analysis For A Biological Effects Study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Everitt, B. Cluster analysis. Qual. Quant. 1980, 14, 75–100. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Clarke, K.R. Nonparametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Chatfield, C.; Collins, A.J. Principal component analysis. In Introduction to Multivariate Analysis; Springer: Boston, MA, USA, 1980; pp. 57–81. [Google Scholar] [CrossRef]
- Clarke, K.R.; Ainsworth, M. A Method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Evans, J.; Cofaigh, C.Ó. Supraglacial debris along the front of the Larsen-A Ice Shelf, Antarctic Peninsula. Antarct. Sci. 2003, 15, 503–506. [Google Scholar] [CrossRef]
- Mincks, S.L.; Smith, C.R.; DeMaster, D.J. Persistence of labile organic matter and microbial biomass in Antarctic shelf sediments: Evidence of a sediment ‘food bank’. Mar. Ecol. Prog. Ser. 2005, 300, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.R.; Mincks, S.; DeMaster, D.J. A synthesis of bentho-pelagic coupling on the Antarctic shelf: Food banks, ecosystem inertia and global climate change. Deep Sea Res. II 2006, 53, 875–894. [Google Scholar] [CrossRef]
- Niemann, H.; Fischer, D.; Graffe, D.; Knittel, K.; Montiel, A.; Heilmayer, O.; Nöthen, K.; Pape, T.; Kasten, S.; Bohrmann, G.; et al. Biogeochemistry of a low-activity cold seep in the Larsen B area, western Weddell Sea, Antarctica. Biogeosciences 2009, 6, 2383–2395. [Google Scholar] [CrossRef]
- Georgieva, M.N.; Wiklund, H.; Bell, J.B.; Eilertsen, M.H.; Mills, R.A.; Little, C.T.; Glover, A.G. A chemosynthetic weed: The tubeworm Sclerolinum contortum is a bipolar, cosmopolitan species. BMC Evol. Biol. 2015, 15, 280. [Google Scholar] [CrossRef] [Green Version]
- Gutt, J.; Schickan, T. Epibiotic relationships in the Antarctic benthos. Antarct. Sci. 1998, 10, 398–405. [Google Scholar] [CrossRef]
- Smith, J.A.; Hillenbrand, C.D.; Subt, C.; Rosenheim, B.E.; Frederichs, T.; Ehrmann, W.; Andersen, T.J.; Wacker, L.; Makinson, K.; Anker, P.; et al. History of the Larsen C Ice Shelf reconstructed from sub–ice shelf and offshore sediments. Geology 2021, 49, 978–982. [Google Scholar] [CrossRef]
- Moore, J.K.; Abbott, M.R.; Richman, J.G. Location and dynamics of the Antarctic Polar Front from satellite sea surface temperature data. J. Geophys. Res. Oceans 1999, 104, 3059–3073. [Google Scholar] [CrossRef]
- Gili, J.-M.; Zapata-Guardiola, R.; Isla, E.; Vaqué, D.; Barbosa, A.; García-Sancho, L.; Quesada, A. Introduction to the special issue on the Life in Antarctica: Boundaries and Gradients in a Changing Environment (XIth SCAR Biology Symposium). Polar Biol. 2015, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cummings, V.J.; Bowden, D.A.; Pinkerton, M.H.; Halliday, N.J.; Hewitt, J.E. Ross Sea Benthic Ecosystems: Macro- and Mega-faunal Community Patterns From a Multi-environment Survey. Front. Mar. Sci. 2021, 8, 21. [Google Scholar] [CrossRef]
- Clarke, A. Seasonality in the antarctic marine environment. Comp. Biochem. Physiol. B 1988, 90, 461–473. [Google Scholar] [CrossRef]
- Torre, L.; Servetto, N.; Eöry, M.L.; Momo, F.; Tatián, M.; Abele, D.; Sahade, R. Respiratory responses of three Antarctic ascidians and a sea pen to increased sediment concentrations. Polar Biol. 2012, 35, 1743–1748. [Google Scholar] [CrossRef]
- Pasotti, F.; Saravia, L.A.; De Troch, M.; Tarantelli, M.S.; Sahade, R.; Vanreusel, A. Benthic Trophic Interactions in an Antarctic Shallow Water Ecosystem Affected by Recent Glacier Retreat. PLoS ONE 2015, 10, e0141742. [Google Scholar] [CrossRef] [Green Version]
- Zwerschke, N.; Sands, C.J.; Roman-Gonzalez, A.; Barnes, D.K.A.; Guzzi, A.; Jenkins, S.; Muñoz-Ramírez, C.; Scourse, J. Quantification of blue carbon pathways contributing to negative feedback on climate change following glacier retreat in West Antarctic fjords. Glob. Chang. Biol. 2022, 28, 8–20. [Google Scholar] [CrossRef]
- Arrigo, K.R. Phytoplankton dynamics within 37 Antarctic coastal polynya systems. J. Geophys. Res. Oceans 2003, 108, 3271. [Google Scholar] [CrossRef]
- Post, A.L.; Lavoie, C.; Domack, E.W.; Leventer, A.; Shevenell, A.; Fraser, A.D. Environmental drivers of benthic communities and habitat heterogeneity on an East Antarctic shelf. Antarct. Sci. 2017, 29, 17–32. [Google Scholar] [CrossRef]
- Jablonski, D. The future of the fossil record. Science 1999, 284, 2114–2116. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Tittensor, D.P.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Chown, S.L.; Sinclair, B.J.; Leinaas, H.P.; Gaston, K.J. Hemispheric Asymmetries in Biodiversity—A Serious Matter for Ecology. PLoS Biol. 2004, 2, e406. [Google Scholar] [CrossRef]
- Woolley, S.N.; Tittensor, D.P.; Dunstan, P.K.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Wintle, B.A.; Worm, B.; O’Hara, T.D. Deep-sea diversity patterns are shaped by energy availability. Nature 2016, 533, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An inverse latitudinal gradient in speciation rate for marine fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef]
- Souster, T.A.; Barnes, D.K.A.; Hopkins, J. Variation in zoobenthic blue carbon in the Arctic’s Barents Sea shelf sediments. Philos. Trans. R. Soc. A 2020, 378, 20190362. [Google Scholar] [CrossRef]
- Grange, L.J.; Smith, C.R. Megafaunal Communities in Rapidly Warming Fjords along the West Antarctic Peninsula: Hotspots of Abundance and Beta Diversity. PLoS ONE 2013, 8, e77917. [Google Scholar] [CrossRef]
- Post, A.L.; O’Brien, P.E.; Edwards, S.; Carroll, A.G.; Malakoff, K.; Armand, L.K. Upper slope processes and seafloor ecosystems on the Sabrina continental slope, East Antarctica. Mar. Geol. 2020, 422, 106091. [Google Scholar] [CrossRef]
- Hillebrand, H.; Blasius, B.; Borer, E.T.; Chase, J.M.; Downing, J.A.; Eriksson, B.K.; Filstrup, C.T.; Harpole, W.S.; Hodapp, D.; Larsen, S.; et al. Biodiversity change is uncoupled from species richness trends: Consequences for conservation and monitoring. J. Appl. Ecol. 2018, 55, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Gutt, J.; Starmans, A.; Dieckmann, G. Impact of iceberg scouring on polar benthic habitats. Mar. Ecol. Prog. Ser. 1996, 137, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Thatje, S.; Hillenbrand, C.D.; Larter, R. On the origin of Antarctic marine benthic community structure. Trends Ecol. Evol. 2005, 20, 534–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, D.K.A.; Sands, C.J.; Hogg, O.T.; Robinson, B.J.O.; Downey, R.V.; Smith, J.A. Biodiversity signature of the Last Glacial Maximum at South Georgia, Southern Ocean. J. Biogeogr. 2016, 43, 2391–2399. [Google Scholar] [CrossRef]
- Jansen, J.; Hill, N.A.; Dunstan, P.K.; McKinlay, J.; Sumner, M.D.; Post, A.L.; Eléaume, M.P.; Armand, L.K.; Warnock, J.P.; Galton-Fenzi, B.K.; et al. Abundance and richness of key Antarctic seafloor fauna correlates with modelled food availability. Nat. Ecol. Evol. 2018, 2, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Gutt, J.; Starmans, A.; Dieckmann, G. Phytodetritus deposited on the Antarctic shelf and upper slope: Its relevance for the benthic system. J. Mar. Syst. 1998, 17, 435–444. [Google Scholar] [CrossRef]
- Sumida, P.Y.G.; Bernardino, A.F.; Stedall, V.P.; Glover, A.G.; Smith, C.R. Temporal changes in benthic megafaunal abundance and composition across the West Antarctic Peninsula shelf: Results from video surveys. Deep Sea Res. II 2008, 55, 2465–2477. [Google Scholar] [CrossRef]
- Han, H.; Lee, S.; Kim, J.I.; Kim, S.H.; Kim, H.C. Changes in a Giant Iceberg Created from the Collapse of the Larsen C Ice Shelf, Antarctic Peninsula, Derived from Sentinel-1 and CryoSat-2 Data. Remote Sens. 2019, 11, 404. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A. Benthic Marine Habitats in Antarctica. In Foundations for Ecological Research West of the Antarctic Peninsula; Ross, R.M., Hofmann, E.E., Quetin, L.B., Eds.; American Geophysical Union: Washington, DC, USA, 1996; pp. 123–133. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, K.W.; Corr, H.F.J.; Makinson, K.; Pudsey, C.J. Rock debris in an Antarctic ice shelf. Ann. Glaciol. 2012, 53, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Grebmeier, J.M.; Barry, J.P. Benthic Processes in Polynyas. In Polynyas: Windows to the World; Smith, W.O., Barber, D.G., Eds.; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2007; Volume 74, pp. 363–390. [Google Scholar]
- Li, J.Y.Q.; Duce, S.; Joyce, K.E.; Xiang, W. SeeCucumbers: Using Deep Learning and Drone Imagery to Detect Sea Cucumbers on Coral Reef Flats. Drones 2021, 5, 28. [Google Scholar] [CrossRef]
- Mileikovsky, S.A. Types of larval development in marine bottom invertebrates, their distribution and ecological significance: A re-evaluation. Mar. Biol. 1971, 10, 193–213. [Google Scholar] [CrossRef]
- Pearse, J.S.; McClintock, J.B.; Bosch, I. Reproduction of antarctic benthic marine-invertebrates—Tempos, modes, and timing. Am. Zool. 1991, 31, 65–80. [Google Scholar] [CrossRef] [Green Version]
- CCAMLR. CCAMLR VME Taxa Classification Guide; CCAMLR: Hobart, Australia, 2009; p. 4. [Google Scholar]
- FAO. International Guidelines for the Management of Deep-Sea Fisheries in the High Seas; FAO: Rome, Italy, 2009; p. 73. [Google Scholar]
- Bridges, A.E.H.; Barnes, D.K.A.; Bell, J.B.; Ross, R.E.; Howell, K.L. Benthic Assemblage Composition of South Atlantic Seamounts. Front. Mar. Sci. 2021, 8, 1530. [Google Scholar] [CrossRef]
- Post, A.L.; O’Brien, P.E.; Beaman, R.J.; Riddle, M.J.; De Santis, L. Physical controls on deep water coral communities on the George V Land slope, East Antarctica. Antarct. Sci. 2010, 22, 371–378. [Google Scholar] [CrossRef]
- Jones, D.O.B.; Bett, B.J.; Tyler, P.A. Depth-related changes to density, diversity and structure of benthic megafaunal assemblages in the Fimbul ice shelf region, Weddell Sea, Antarctica. Polar Biol. 2007, 30, 1579–1592. [Google Scholar] [CrossRef]
- Ziegler, A.F.; Smith, C.R.; Edwards, K.F.; Vernet, M. Glacial dropstones: Islands enhancing seafloor species richness of benthic megafauna in West Antarctic Peninsula fjords. Mar. Ecol. Prog. Ser. 2017, 583, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Post, A.L.; Lavoie, C.; Domack, E.W.; Leventer, A.; Fernandez, R.; Shipboard Sci, P. Dropstones on a glaciated continental shelf as key habitat, Sabrina Shelf, East Antarctica. In Seafloor Geomorphology as Benthic Habitat: GeoHab Atlas of Seafloor Geomorphic Features and Benthic Habitats, 2nd ed.; Harris, P.T., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2020; pp. 641–653. [Google Scholar]
- Dayton, P.K.; Oliver, J.S. Antarctic Soft-Bottom Benthos in Oligotrophic and Eutrophic Environments. Science 1977, 197, 55–58. [Google Scholar] [CrossRef]
- Clark, G.F.; Stark, J.S.; Palmer, A.S.; Riddle, M.J.; Johnston, E.L. The Roles of Sea-Ice, Light and Sedimentation in Structuring Shallow Antarctic Benthic Communities. PLoS ONE 2017, 12, e0168391. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Metz, S.E.A.; Gerdes, D.; Richter, C. Benthic fauna declined on a whitening Antarctic continental shelf. Nat. Commun. 2020, 11, 2226. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Date(s) Sampled | Latitude (°S) | Longitude (°W) | Water Depth (m) 1 | Distance from Ice-Shelf Front (m) 2 |
|---|---|---|---|---|---|
| LCP1—Bawden Ice Rise | 21–22 January 2019 | −66.6975 | −60.2981 | 397 | 777 |
| LCP2—Jason Peninsula | 20 January 2019 | −66.1868 | −60.4742 | 299 | 2752 |
| LCP3—Jason Peninsula | 14 January 2019 | −66.0219 | −60.3526 | 400 | 9024 |
| LCP4—Jason Peninsula | 23 January 2019 | −66.1544 | −60.1594 | 349 | 16,459 |
| Functional Group | Example Taxa |
|---|---|
| Deposit-feeding crawlers (epifaunal) | Holothurians |
| Soft infaunal deposit-feeders | None discerned |
| Hard burrowing deposit-feeders | Irregular echinoids |
| Flexible strategists | Ophiuroids |
| Grazers | Regular echinoids |
| Soft sessile predator/scavengers 1 | Pennatulaceans, soft corals, anemones (actiniarians), hydroids |
| Hard sessile predator/scavengers 1 | Scleractinian corals, calcaxonian whips, hydrocorals |
| Soft mobile predator/scavengers | Nemerteans, nudibranchs, octopi |
| Hard mobile predator/scavengers | Fish, gastropods, asteroids |
| Arthropod predator/scavengers | Pycnogonids, decapod shrimps |
| Pioneer sessile suspension feeders | Encrusting bryozoans, ascidians, some polychaetes |
| Climax sessile suspension feeders | Demosponges, hexactinellids, brachiopods, some bryozoans |
| Sedentary suspension feeders | None discerned |
| Mobile suspension feeders | Crinoids |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frinault, B.A.V.; Christie, F.D.W.; Fawcett, S.E.; Flynn, R.F.; Hutchinson, K.A.; Montes Strevens, C.M.J.; Taylor, M.L.; Woodall, L.C.; Barnes, D.K.A. Antarctic Seabed Assemblages in an Ice-Shelf-Adjacent Polynya, Western Weddell Sea. Biology 2022, 11, 1705. https://doi.org/10.3390/biology11121705
Frinault BAV, Christie FDW, Fawcett SE, Flynn RF, Hutchinson KA, Montes Strevens CMJ, Taylor ML, Woodall LC, Barnes DKA. Antarctic Seabed Assemblages in an Ice-Shelf-Adjacent Polynya, Western Weddell Sea. Biology. 2022; 11(12):1705. https://doi.org/10.3390/biology11121705
Chicago/Turabian StyleFrinault, Bétina A. V., Frazer D. W. Christie, Sarah E. Fawcett, Raquel F. Flynn, Katherine A. Hutchinson, Chloë M. J. Montes Strevens, Michelle L. Taylor, Lucy C. Woodall, and David K. A. Barnes. 2022. "Antarctic Seabed Assemblages in an Ice-Shelf-Adjacent Polynya, Western Weddell Sea" Biology 11, no. 12: 1705. https://doi.org/10.3390/biology11121705
APA StyleFrinault, B. A. V., Christie, F. D. W., Fawcett, S. E., Flynn, R. F., Hutchinson, K. A., Montes Strevens, C. M. J., Taylor, M. L., Woodall, L. C., & Barnes, D. K. A. (2022). Antarctic Seabed Assemblages in an Ice-Shelf-Adjacent Polynya, Western Weddell Sea. Biology, 11(12), 1705. https://doi.org/10.3390/biology11121705