
Assessment of Fish Diversity in the Ma’an Archipelago Special Protected Area Using Environmental DNA

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
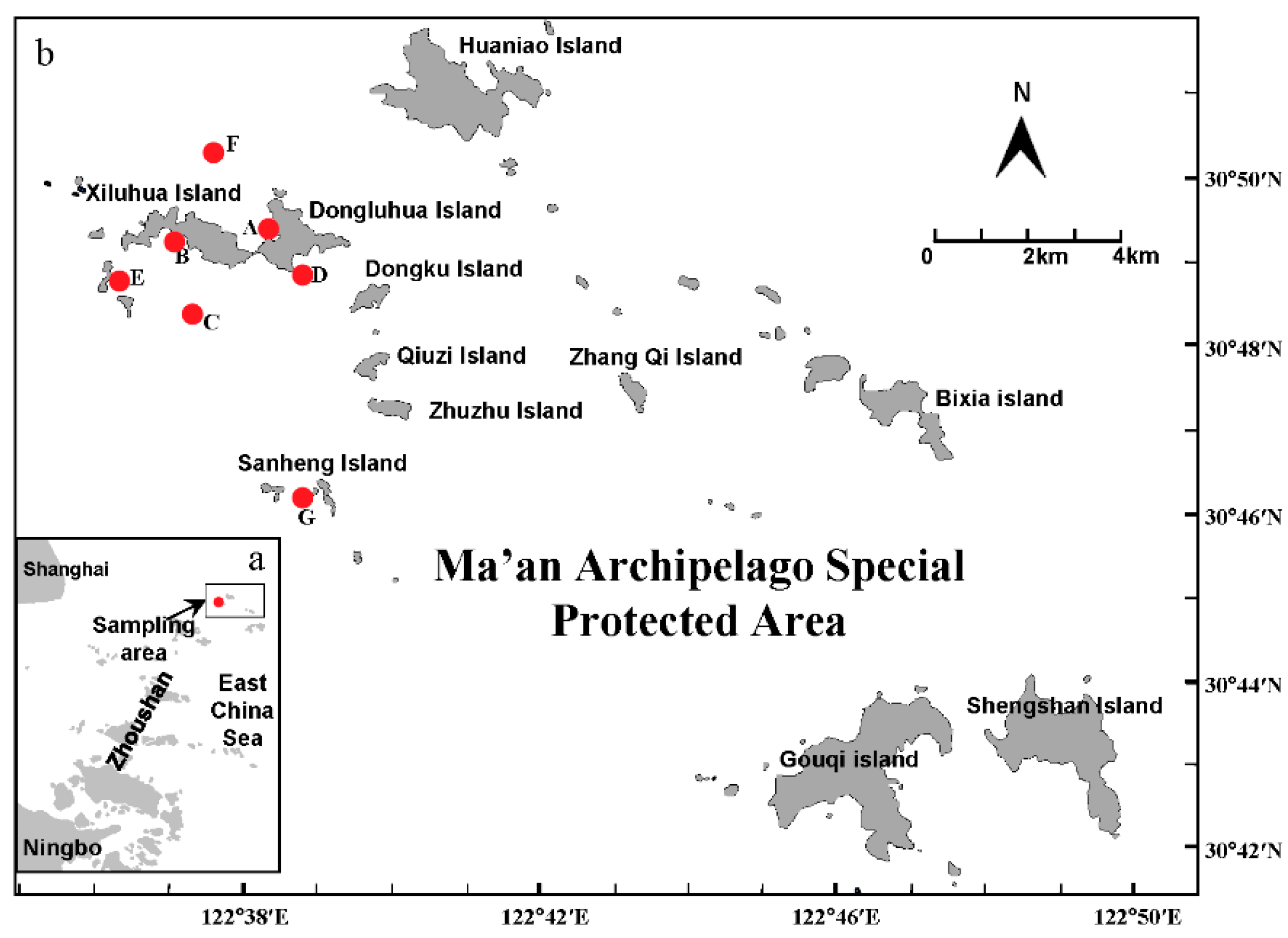
2.1. Sampling Time and Station
2.2. Water Sample Collection and Processing
2.3. Environmental DNA Testing
PCR Amplification and Sequencing
2.4. Data Analysis
2.4.1. Fish Species Determination
2.4.2. Fish Diversity Analysis
3. Analysis of Results
3.1. Results of eDNA Sequencing
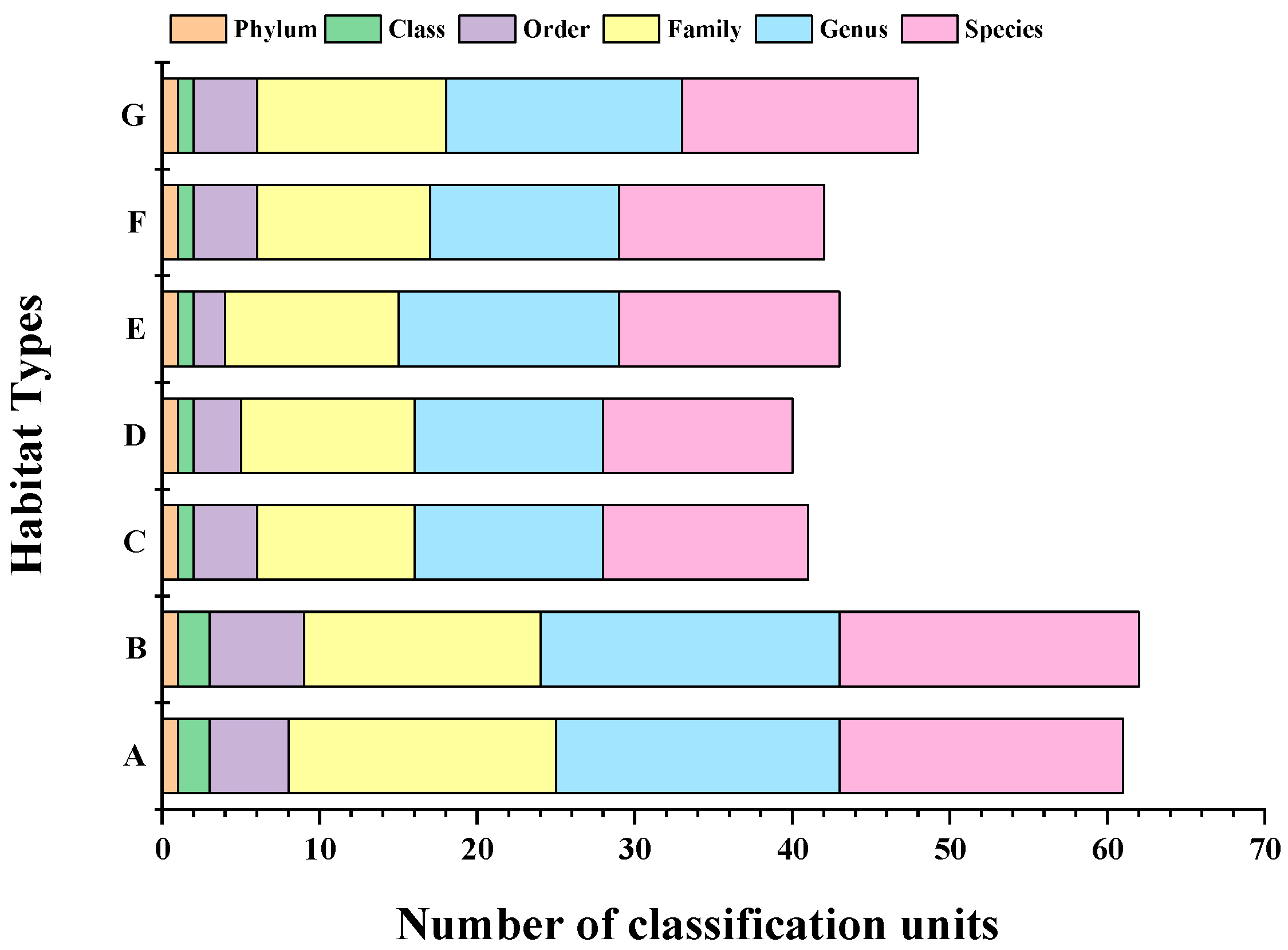
3.2. Fish Species Composition
3.2.1. Fish Species Composition in Different Habitats
3.2.2. Species Diversity Indices of Different Habitats
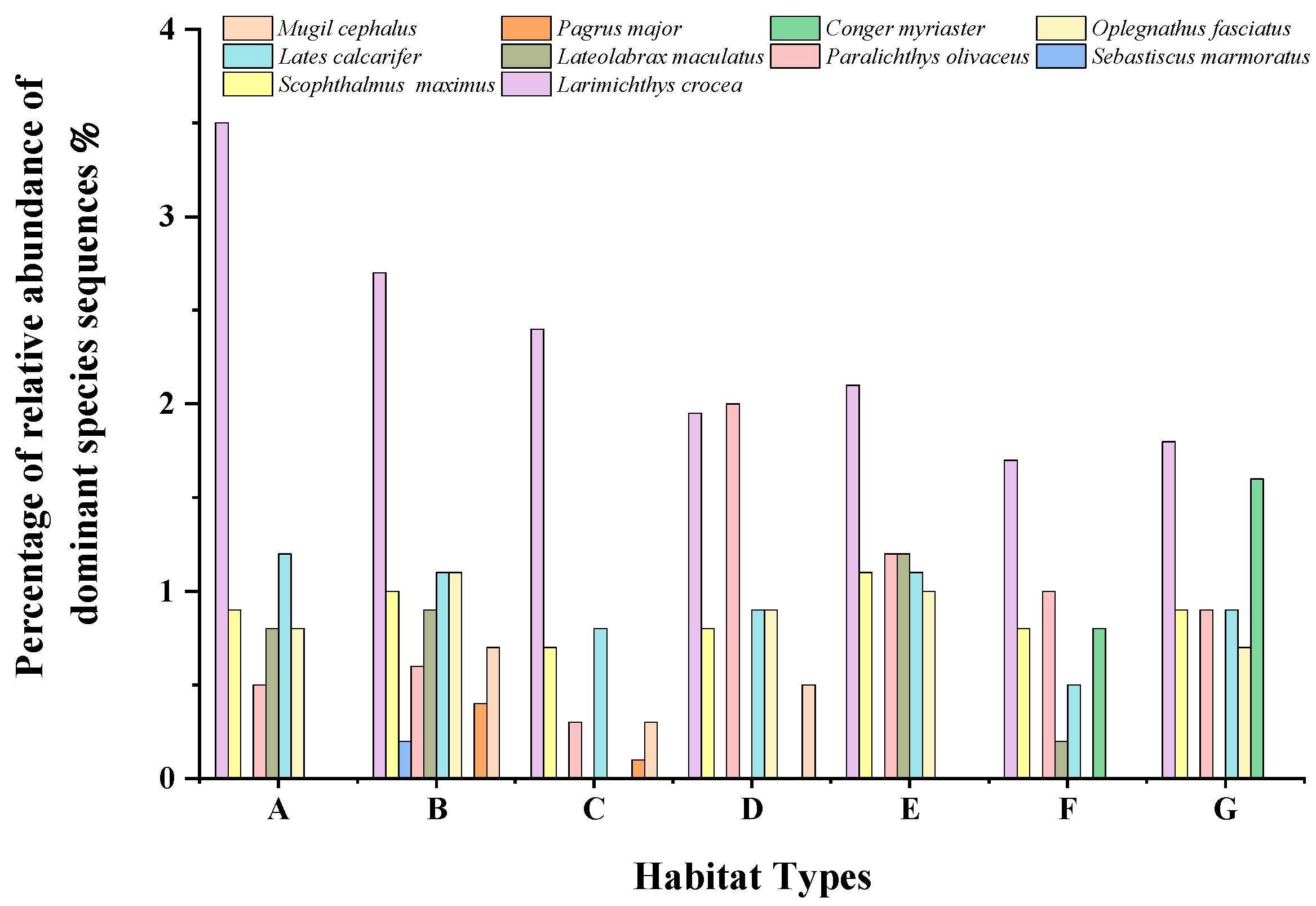
3.2.3. Composition of Dominant Fish Species
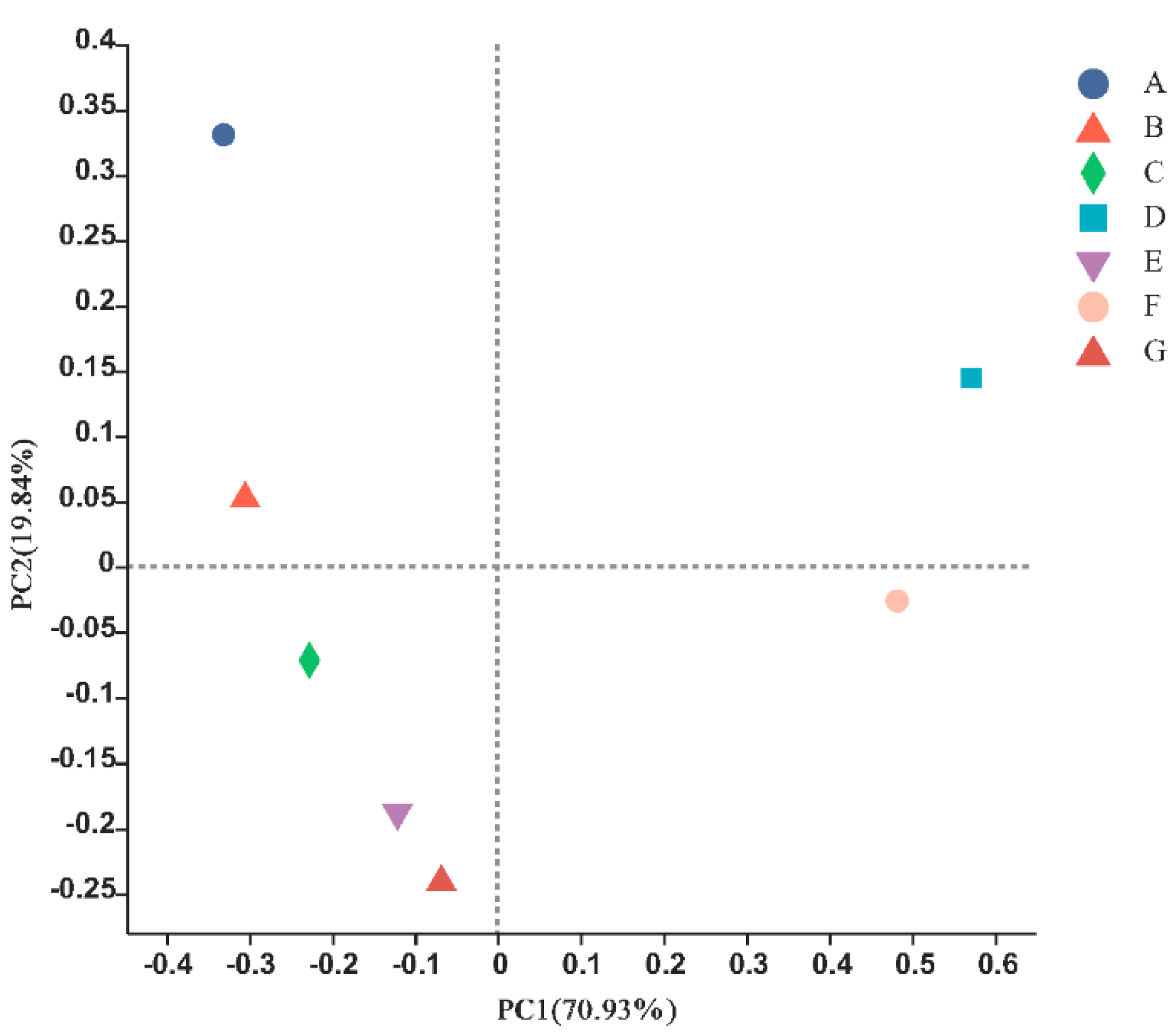
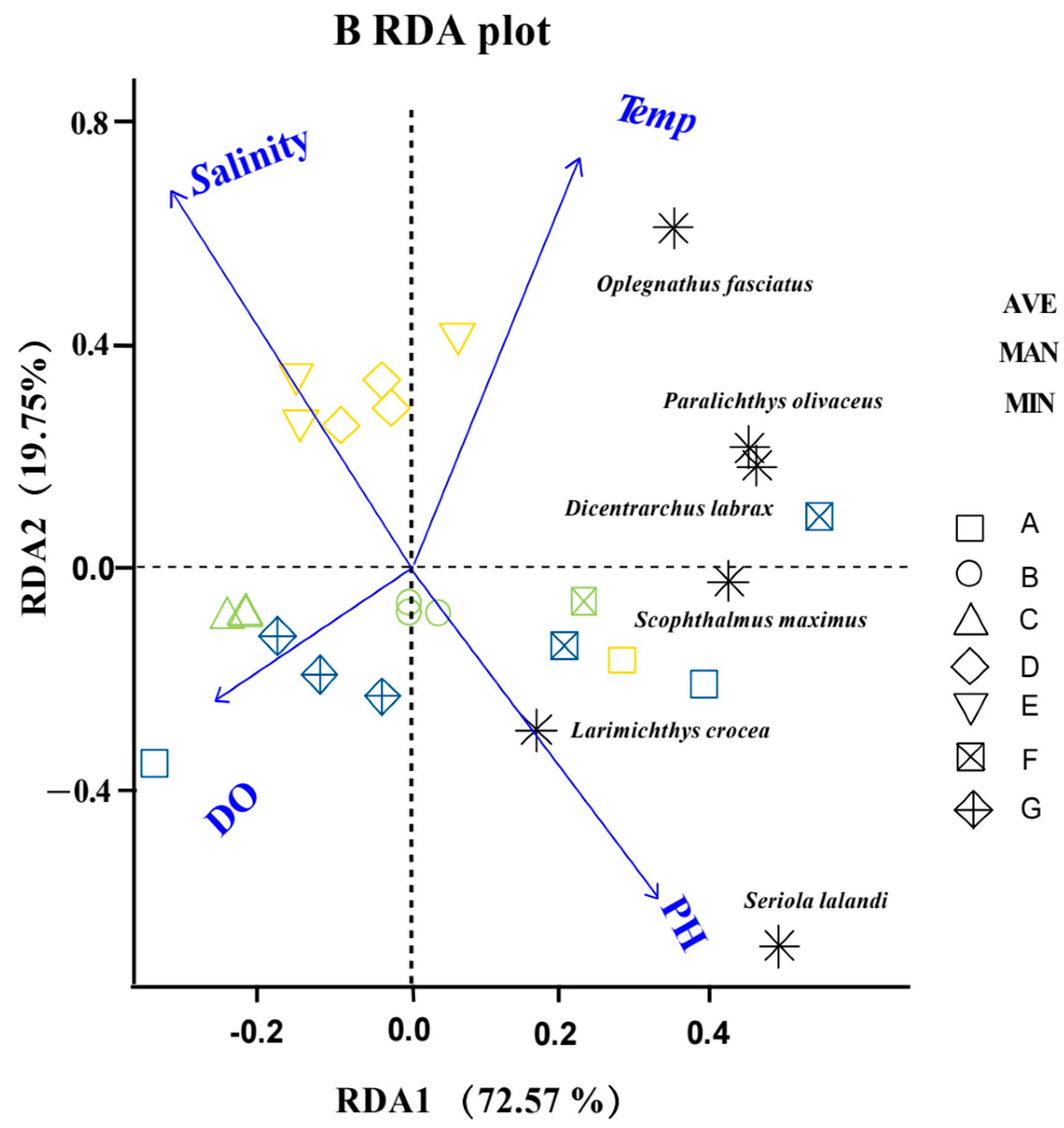
3.3. Correlation Analysis for Each Site
4. Discussion
4.1. Category Composition
4.1.1. Comparison of Fish Diversity
4.1.2. Analysis of Fish Species Composition by Different Survey Methods
4.2. Biomass Analysis
4.3. Differences between Different Habitats
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dahlgren, C.; Marr, J. Back reef systems: Important but overlooked components of tropical marine ecosystems. Bull. Mar. Sci. 2004, 75, 145–152. [Google Scholar]
- Wang, K.; Zhang, S.Y.; Wang, Z.H.; Zhao, J.; Jiang, R.J. Dietary composition and feeding strategy of Agrammus agrammus off the Ma’an Archipelago special marine reserves. J. Shanghai Ocean. Univ. 2014, 23, 251–257. [Google Scholar]
- Liu, H. Study on Bio-Resource Monitoring and Assessing Techniques and Aoolication in Offshore Island Area with Artificial Reefs; Institute of Oceanology of the Chinese Academy of Sciences: Qingdao, China, 2014. [Google Scholar]
- Huang, H.; Zhang, Y.Y.; Liu, C.Y. Coral Reef Habitat and Resources Restoration in Tropical Island Marine Ranching. Top. Discuss. 2020, 16, 225–230. [Google Scholar] [CrossRef]
- Ali, A.; Abdullah, M.R.; Safuan, C.D.M.; Afiq-Firdaus, A.M.; Bachok, Z.; Akhir, M.F.M.; Latif, R.; Muhamad, A.; Seng, T.H.; Roslee, A.; et al. Side-Scan Sonar Coupled with Scuba Diving Observation for Enhanced Monitoring of Benthic Artificial Reefs along the Coast of Terengganu, Peninsular Malaysia. J. Mar. Sci. Eng. 2022, 10, 1309. [Google Scholar] [CrossRef]
- Wang, Z.H.; Gong, F.X.; Wu, Z.L.; Yuan, X.B.; Zhang, S.Y. Efficiency of multi-mesh gillnets and multi-mesh trammel nets in collecting fish from rocky reef and sandy beach habitats. Chin. J. Ecol. 2013, 32, 507–514. [Google Scholar] [CrossRef]
- Xu, S.N.; Wang, Z.H.; Liang, J.L.; Zhang, S.Y. Use of different sampling tools for comparison of fish-aggregating effects along horizontal transect at two artificial reef sites in Shengs. J. Fish. China 2016, 40, 820–831. [Google Scholar]
- Baldigo, B.P.; Sporn, L.A.; George, S.D.; Ball, J.A. Efficacy of Environmental DNA to Detect and Quantify Brook Trout Populations in Headwater Streams of the Adirondack Mountains, New York. Trans. Am. Fish. Soc. 2017, 146, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Van Denderen, P.D.; Bolam, S.G.; Hiddink, J.G.; Jennings, S.; Kenny, A.; Rijnsdorp, A.D.; van Kooten, T. Similar effects of bottom trawling and natural disturbance on composition and function of benthic communities across habitats. Mar. Ecol. Prog. Ser. 2015, 541, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.J.; Li, M.; Wang, W. Application of Environmental DNA Technology in Aquatic Ecosystem. Prog. Fish. Sci. 2018, 39, 23–26. [Google Scholar] [CrossRef]
- Dejean, T.; Valentini, A.; Duparc, A.; Pellier-Cuit, S.; Pompanon, F.; Taberlet, P.; Miaud, C. Persistence of Environmental DNA in Freshwater Ecosystems. PLoS ONE 2011, 6, e23398. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, S.Y.; Zhou, X.J.; Chen, Q.M. Comparative analysis of two sampling gillnets of rocky reef area in Gouqi Islands, Shengsi, Zhejiang. J. Fish. China 2013, 37, 311–320. [Google Scholar] [CrossRef]
- Tong, W.J.; Zhang, S.Y. Preliminary study on distribution of fishery resources in LvHua-SanHeng sea area based on Kriging. J. Biol. 2011, 28, 62–67. [Google Scholar] [CrossRef]
- Li, M.; Wei, T.T.; Shi, B.Y.; Hao, X.Y.; Xu, X.Y.; Sun, H.Y. Biodiversity monitoring of freshwater benthic macroinvertebrates using environmental DNA. Biodivers. Sci. 2019, 27, 480–490. [Google Scholar] [CrossRef]
- Doi, H.; Takahara, T.; Minamoto, T.; Matsuhashi, S.; Uchii, K.; Yamanaka, H. Droplet Digital Polymerase Chain Reaction (PCR) Outperforms Real-Time PCR in the Detection of Environmental DNA from an Invasive Fish Species. Environ. Sci. Technol. 2015, 49, 5601–5608. [Google Scholar] [CrossRef]
- Kelly, R.P.; Port, J.A.; Yamahara, K.M.; Crowder, L.B. Using Environmental DNA to Census Marine Fishes in a Large Mesocosm. PLoS ONE 2014, 9, e86175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahara, T.; Ikebuchi, T.; Doi, H.; Minamoto, T. Using environmental DNA to estimate the seasonal distribution and habitat preferences of a Japanese basket clam in Lake Shinji, Japan. Estuar. Coast. Shelf Sci. 2019, 221, 15–20. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- West, K.M.; Stat, M.; Harvey, E.S.; Skepper, C.L.; DiBattista, J.D.; Richards, Z.T.; Travers, M.J.; Newman, S.J.; Bunce, M. eDNA metabarcoding survey reveals fine-scale coral reef community variation across a remote, tropical island ecosystem. Mol. Ecol. 2020, 29, 1069–1086. [Google Scholar] [CrossRef]
- Aglieri, G.; Baillie, C.; Mariani, S.; Cattano, C.; Calo, A.; Turco, G.; Spatafora, D.; Di Franco, A.; Di Lorenzo, M.; Guidetti, P.; et al. Environmental DNA effectively captures functional diversity of coastal fish communities. Mol. Ecol. 2021, 30, 3127–3139. [Google Scholar] [CrossRef]
- Wang, Z.H. Fish Community Patterns in Meta-Habitat: A Case Study from Ma’an Archipelago; Shanghai Ocean University: Shanghai, China, 2011. [Google Scholar]
- Wang, Z.H.; Zhao, J.; Wang, K.; Zhang, S.Y. Fish community ecology in rocky reef habitat of Ma’ an Archipelago Ⅱ. Spatio-temporal patterns of community structure. Acta Ecol. Sin. 2013, 33, 6218–6226. [Google Scholar] [CrossRef]
- Wang, Z.H.; Zhang, S.Y.; Chen, Q.M.; Xu, Q.; Wang, K. Fish community ecology in rocky reef habitat of Ma’an Archipelago. I. Species composition and diversity. Biodivers. Sci. 2012, 20, 41–50. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, S.Y.; Wang, Z.; Wang, K. Analysis on community structure and diversity of fish and macroinvertebrate in Shengsi artificial reef area. J. Fish. Sci. China 2010, 17, 1045–1056. [Google Scholar]
- Han, X.D.; Zhang, S.Y.; Wang, Z.H.; Wang, K.; Lin, J.; Deng, M.Q.; Wu, X.C. Fish community structure and its relationship with environmental factors in the Ma’an Archipelago and its eastern waters. J. Fish. China 2019, 43, 1483–1497. [Google Scholar] [CrossRef]
- Boussarie, G.; Bakker, J.; Wangensteen, O.S.; Mariani, S.; Bonnin, L.; Juhel, J.-B.; Kiszka, J.J.; Kulbicki, M.; Manel, S.; Robbins, W.D.; et al. Environmental DNA illuminates the dark diversity of sharks. Sci. Adv. 2018, 4, eaap9661. [Google Scholar] [CrossRef] [PubMed]
- Sigsgaard, E.E.; Nielsen, I.B.; Carl, H.; Krag, M.A.; Knudsen, S.W.; Xing, Y.; Holm-Hansen, T.H.; Moller, P.R.; Thomsen, P.F. Seawater environmental DNA reflects seasonality of a coastal fish community. Mar. Biol. 2017, 164, 128. [Google Scholar] [CrossRef]
- Stat, M.; Huggett, M.J.; Bernasconi, R.; DiBattista, J.D.; Berry, T.E.; Newman, S.J.; Harvey, E.S.; Bunce, M. Ecosystem biomonitoring with eDNA: Metabarcoding across the tree of life in a tropical marine environment. Sci. Rep. 2017, 7, 12240. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Fang, C.; Zhi-Gang, W.; Fan, X.; Dan, Y.; YongDe, C. AeDNA: Aquatic Envionment DNA Database. Acta Hydrobiol. Sin. 2022, 46, 1741–1747. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Benson, D.A.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2014, 42, D32–D37. [Google Scholar] [CrossRef] [Green Version]
- Lamy, T.; Pitz, K.J.; Chavez, F.P.; Yorke, C.E.; Miller, R.J. Environmental DNA reveals the fine-grained and hierarchical spatial structure of kelp forest fish communities. Sci. Rep. 2021, 11, 14439. [Google Scholar] [CrossRef]
- Polanco Fernández, A.; Marques, V.; Fopp, F.; Juhel, J.B.; Borrero-Pérez, G.H.; Cheutin, M.C.; Dejean, T.; González Corredor, J.D.; Acosta-Chaparro, A. Comparing environmental DNA metabarcoding and underwater visual census to monitor tropical reef fishes. Environ. DNA 2021, 3, 142–156. [Google Scholar] [CrossRef]
- Wang, Z.H.; Zhang, S.Y.; Wang, K. Fish and macroinvertebrates community structure in artificial habitat around Sanheng Isle, Shengsi, China. Acta Ecol. Sin. 2010, 30, 2026–2035. [Google Scholar]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Le, Z.Y.; Tang, Y.W.; Wang, Z.Y. Reason analysis and resource protection suggestions on the frequent catch of wild large yellow croaker in Zhejiang Province sea area. Chin. Fish. Econ. 2022, 40, 53–60. [Google Scholar]
- Klymus, K.E.; Richter, C.A.; Chapman, D.C.; Paukert, C. Quantification of eDNA shedding rates from invasive bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. Biol. Conserv. 2015, 183, 77–84. [Google Scholar] [CrossRef]
- Ebisawa, A.; Kanashiro, K.; Ohta, I.; Uehara, M.; Nakamura, H. Changes of group construction accompanying with growth and maturity in blue-barred parrotfish (Scarus ghobban), and influences of the fishing targeting the immature group to the stock. Reg. Stud. Mar. Sci. 2016, 7, 32–42. [Google Scholar] [CrossRef]
- Wang, Z.H.; Liang, J.L.; Zhang, S.Y. Comparison of pelagic and benthic fish assemblages in mussel farming habitat. J. Biol. 2015, 34, 753–759. [Google Scholar] [CrossRef]
- Lin, J.; Deng, M.X.; Zhang, S.Y.; Yan, Q. Seasonal variation of surface water temperature and its ecological impacts in a mussel aquaculture farm. J. Shanghai Ocean. Univ. 2015, 25, 918–926. [Google Scholar] [CrossRef]
- Lin, Y.; Li, J.; Wang, Z.; Zhang, S.; Wang, K.; Li, X. A Comparison of Fish Diversity in Rocky Reef Habitats by Multi-Mesh Gillnets and Environmental DNA Metabarcoding. Front. Ecol. Evol. 2022, 10, 874558. [Google Scholar] [CrossRef]
- Wang, Z.H.; Shen, H.; Lin, J.; Zhang, S.Y.; Zhong, J.M.; Chen, Y.F.; Liu, Z.B. Spatial and temporal distribution of small yellow croaker (Larimichthys polyactis) in Eastern Ma’an Archipelago. J. Fish. China 2022, 46, 959–972. [Google Scholar] [CrossRef]
- Zamani, N.P.; Zuhdi, M.F.; Madduppa, H. Environmental DNA biomonitoring reveals seasonal patterns in coral reef fish community structure. Environ. Biol. Fishes 2022, 105, 971–991. [Google Scholar] [CrossRef]
- Ling, J.Z.; Jiang, Y.Z.; Sun, P.; Yuan, X.W.; Zhang, H.; Tang, B.J. Application and evaluation of environmental DNA technology in fish diversity research in Xiangshan Bay. J. Fish. Sci. China 2021, 28, 205–214. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.M.; Tang, J.H.; Zhang, H.; Zhong, F.; Wu, L.; Gao, Y.S. Temporal andspatial distributionof Miichthysmiiuy in Jiangsucoastal waters. Haiyang Xuebao 2010, 32, 95–106. [Google Scholar]
- Luo, B.Z. Seasonal growth the large yellow croaker, Pseudosciaena crocea (rich.), off chekiang. Oceanol. Limnol. Sin. 1966, 8, 121–139. [Google Scholar]
- Xu, Z.L.; Chen, J.J. Analysis of migratory route Larimichthys crocea in the East China Sea and Yellow Sea. J. Fish. China 2011, 35, 429–437. [Google Scholar] [CrossRef]
- Wang, X.Y.; Lu, G.Q.; Zhao, L.L.; Du, X.Q.; Gao, T.X. Assessment of fishery resources using environmental DNA: The large yellow croaker (Larimichthys crocea) in the East China Sea. Fish. Res. 2022, 235, 105813. [Google Scholar] [CrossRef]
- Lou, B.; Mao, G.M.; Shi, H.L.; Luo, J.; Jian, X.; Zheng, D.M. Experiment on Artifi cial Domestication of Paralichthys olivaces (Temminck et Schlegel) Captured from Natural Waters along the Coast of Zhoushan. Fish. Inf. Strategy 2008, 23, 15–17. [Google Scholar]
- Sato, M.; Inoue, N.; Nambu, R.; Furuichi, N.; Imaizumi, T.; Ushio, M. Quantitative assessment of multiple fish species around artificial reefs combining environmental DNA metabarcoding and acoustic survey. Sci. Rep. 2021, 11, 19477. [Google Scholar] [CrossRef]
- Yu, N.J.; Yu, C.G.; Xu, Y.J.; Zheng, J.; Liu, K.; Zhang, P.Y. Fish community structure and biodiversity in the offshore waters of Zhoushan Islands in spring and autumn. J. Fish. China 2021, 45, 1374–1383. [Google Scholar] [CrossRef]
- Xu, Y.; Ma, L.; Sun, Y.; Li, X.Z.; Wang, H.F.; Zhang, H.M. Spatial variation of demersal fish diversity and distribution in the East China Sea: Impact of the bottom branches of the Kuroshio Current. J. Sea Res. 2019, 144, 22–32. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Liu, S.R.; Zhou, X.J.; Wang, Z.H.; Wang, K. Ecological function of seaweed-formed habitat and discussion of its application to sea ranching. J. Fish. China 2019, 43, 2004–2014. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Sun, H.C. Research progress on seaweed bed ecosystem and its engineering. Chin. J. Appl. Ecol. 2007, 18, 1647–1653. [Google Scholar] [CrossRef]
- Komatsu, T.; Mikami, A.; Sultana, S.; Ishida, K.; Hiraishi, T.; Tatsukawa, K.-I. Hydro-acoustic methods as a practical tool for cartography of seagrass beds. Otsuchi Mar. Sci. 2003, 28, 72–79. [Google Scholar]
- Wang, K.; Zhang, S.Y.; Wang, Z.H.; Zhao, J.; Xu, M.; Lin, J. Dietary composition and food competition of six main fish species in rocky reef habitat off Gouqi Island. Chin. J. Appl. Ecol. 2012, 23, 536–544. [Google Scholar] [CrossRef]
- Diamant, A.; Tuvia, A.B.; Baranes, A.; Golani, D. An analysis of rocky coastal eastern Mediterranean fish assemblages and a comparison with an adjacent small artificial reef. J. Exp. Mar. Biol. Ecol. 1986, 97, 269–285. [Google Scholar] [CrossRef]
- Guo, J.; Wang, T.; Defang, C.; Yong, L.; Quan, Q.; Wang, J. Feeding habits of Acanthopagrus schlegeli in the Daya Bay. J. Fish. Sci. China 2021, 28, 1041–1050. [Google Scholar] [CrossRef]
- Dong, S.; Xiuqing, B.; Chengqing, Y. Analysis of Induced Corrosion by Fouling Organisms on Offshore Platform and Its Research Progress. Mater. Prot. 2018, 51, 116–124. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Xu, M.; Wang, Z.H. Review of artificial reef and stock enhancement. Fish. Mod. 2010, 37, 55–58. [Google Scholar]
- Komyakova, V.; Chamberlain, D.; Swearer, S.E. A multi-species assessment of artificial reefs as ecological traps. Ecol. Eng. 2021, 171, 106394. [Google Scholar] [CrossRef]
- Granneman, J.E.; Steele, M.A. Effects of reef attributes on fish assemblage similarity between artificial and natural reefs. Ices J. Mar. Sci. 2015, 72, 2385–2397. [Google Scholar] [CrossRef]
- Zhang, R.L.; Sun, D.Y.; Hou, C.W.; Zhao, J.M. Characteristics of benthic fishery community at natural reefs and artificial reefs located in oddshore area. Oceanol. Limnol. Sin. 2021, 52, 697–707. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, S.Y.; Cheng, X.P.; Lin, J. Acoustic estimation of fisheries resources off Ma’an Archipelago. J. Fish. China 2020, 44, 1695–1706. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Type | Sample Label | Total Number of Bases Sequenced | Accounted for | Effective Sequence |
|---|---|---|---|---|
| A | A1 | 4,457,369 | 94.35 | 52,440 |
| A2 | 4,563,778 | 93.97 | 53,692 | |
| A3 | 5,254,263 | 94.15 | 61,815 | |
| B | B1 | 5,784,349 | 94.40 | 68,051 |
| B2 | 4,042,183 | 94.74 | 47,555 | |
| B3 | 4,118,932 | 93.73 | 48,458 | |
| C | C1 | 4,325,293 | 95.41 | 50,886 |
| C2 | 4,301,465 | 94.62 | 50,605 | |
| C3 | 3,877,771 | 93.90 | 45,621 | |
| D | D1 | 4,330,038 | 93.58 | 50,942 |
| D2 | 4,178,398 | 94.54 | 49,158 | |
| D3 | 4,064,025 | 93.18 | 47,812 | |
| E | E1 | 3,753,436 | 93.70 | 44,158 |
| E2 | 3,845,085 | 95.75 | 45,236 | |
| E3 | 4,239,784 | 93.25 | 49,880 | |
| F | F1 | 5,016,051 | 94.58 | 59,012 |
| F2 | 4,285,021 | 94.87 | 50,412 | |
| F3 | 4,031,488 | 94.86 | 47,429 | |
| G | G1 | 5,361,963 | 94.83 | 63,082 |
| G2 | 4,028,498 | 94.05 | 47,394 | |
| G3 | 4,428,378 | 94.31 | 52,099 |
| Order and Species | Habitat Type | Ecological Types | ||||||
|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | ||
| Anguilliformes | ||||||||
| 1 Conger myriaster | + | + | + | + | **▲● | |||
| Clupeiformes | ||||||||
| 2 Engraulis japonicus | + | + | + | + | **▲● | |||
| 3 Coilia nasus | + | *△● | ||||||
| 4 Ilisha elongata | + | ***▲● | ||||||
| Perciformes | ||||||||
| 5 Johnius belangerii | + | + | + | *◆○ | ||||
| 6 Larimichthys polyactis | + | + | + | *▲● | ||||
| 7 Larimichthys crocea | + | + | + | + | + | + | + | *▲● |
| 8 Nibea albiflora | + | *▲● | ||||||
| 9 Miichthys miiuy | + | + | + | + | + | *▲● | ||
| 10 Pagrus major | + | + | + | + | ***◆○ | |||
| 11 Acanthopagrus schlegelii | + | + | + | + | + | □◆○ | ||
| 12 Oplegnathus fasciatus | + | + | + | + | + | + | + | *◆○ |
| 13 Scomberomorus niphonius | + | *◆○ | ||||||
| 14 Psenopsis anomala | + | *◆○ | ||||||
| 15 Zoarces viviparus | + | ****◆○ | ||||||
| 16 Lateolabrax maculatus | + | + | + | + | + | □▲● | ||
| 17 Mugil cephalus | + | + | + | + | + | □◆○ | ||
| 18 Lates calcarifer | + | + | + | + | + | + | + | *▲◎ |
| 19 Dicentrarchus labrax | + | + | + | + | + | + | + | ◎ |
| 20 Sparus aurata | + | + | + | + | + | + | + | □◆◎ |
| 21 Seriola lalandi | + | + | + | + | + | + | + | *▲● |
| 22 Scarus ghobban | + | ◎ | ||||||
| 23 Siganus fuscescens | + | □◆○ | ||||||
| Scorpaeniformes | ||||||||
| 24 Sebastiscus marmoratus | + | + | + | *◆○ | ||||
| Pleuronectiformes | ||||||||
| 25 Paralichthys olivaceus | + | + | + | + | + | + | + | ***◆○ |
| 26 Scophthalmus maximus | + | + | + | + | + | + | + | *****◆○ |
| Tetraodontiformes | ||||||||
| 27 Takifugu niphobles | + | + | *◆○ | |||||
| Number of categories | 18 | 19 | 13 | 12 | 14 | 13 | 18 | |
| Special Species | 3 | 2 | 1 | 0 | 0 | 0 | 2 | |
| Habitat Type | Alpha Diversity Indices | |||
|---|---|---|---|---|
| Shannon–Wiener Diversity Index | Simpson Diversity Index | Chao1 Index | Pielou Evenness Index | |
| A | 2.274 ± 0.163 | 0.839 ± 0.034 | 86.671 ± 7.178 | 0.786 ± 0.029 |
| B | 2.352 ± 0.161 | 0.868 ± 0.037 | 95.912 ± 7.581 | 0.799 ± 0.031 |
| C | 1.865 ± 0.127 | 0.757 ± 0.036 | 74.141 ± 4.997 | 0.727 ± 0.033 |
| D | 1.993 ± 0.102 | 0.826 ± 0.018 | 76.544 ± 4.657 | 0.802 ± 0.017 |
| E | 2.106 ± 0.100 | 0.846 ± 0.019 | 81.781 ± 4.462 | 0.798 ± 0.020 |
| F | 2.000 ± 0.122 | 0.816 ± 0.023 | 77.505 ± 5.425 | 0.779 ± 0.024 |
| G | 2.245 ± 0.113 | 0.863 ± 0.013 | 88.352 ± 4.415 | 0.829 ± 0.038 |
| Habitat Type | eDNA * | Bottom Trawling [21] | Bottom Trawling (Unpublished Data) | Bottom Trawling [42] | Multiple Gillnet [22] | ||||
|---|---|---|---|---|---|---|---|---|---|
| Species * | Species | Specific Name | Species | Specific Name | Species | Specific Name | Species | Specific Name | |
| Oceanic ranch | 18 | 9 | N. lbiflora L. polyacti S. marmoratus J. belangerii Hexagrammos indicus Chelidonichthys kumu P. olivaceus T. kammalensis C. nasus | 7 | Engraulis japonicus J. belangerii M. miiuy P. anomala Pampus argenteus L. polyacti C. lucidu | 5 | J. belangerii L. polyacti S. marmoratus Stephanolepis cirrhifer P. olivaceus | 4 | S. marmoratus T. kammalensis M. miiuy A. schlegelii |
| Seaweed beds | 18 | 6 | N. albiflora S. marmoratus L. maculatus A. schlegelii Hexagrammos otakii Agrammus agrammus | - | - | 7 | Trichiurus lepturus J. belangerii M. miiuy P. anomala P. argenteus L. polyacti C. lucidu Muraenesocis cinereus | ||
| Cage culture area and nearby rock reefs | 12 | 5 | S. marmoratus N. albiflora C. myriaster L. maculatus A. agrammus | 2 | N. albiflora Cynoglossus joyneri | - | - | ||
| Mussel culture area | 19 | 4 | S. marmoratus J. belangerii L. polyacti N. albiflora | - | - | - | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, X.; Zhao, X.; Chen, J.; Wang, Z.; Chen, L.; Zhang, S.; Wang, K. Assessment of Fish Diversity in the Ma’an Archipelago Special Protected Area Using Environmental DNA. Biology 2022, 11, 1832. https://doi.org/10.3390/biology11121832
Wang Y, Li X, Zhao X, Chen J, Wang Z, Chen L, Zhang S, Wang K. Assessment of Fish Diversity in the Ma’an Archipelago Special Protected Area Using Environmental DNA. Biology. 2022; 11(12):1832. https://doi.org/10.3390/biology11121832
Chicago/Turabian StyleWang, Yuqing, Xunmeng Li, Xu Zhao, Jianqu Chen, Zhenhua Wang, Lili Chen, Shouyu Zhang, and Kai Wang. 2022. "Assessment of Fish Diversity in the Ma’an Archipelago Special Protected Area Using Environmental DNA" Biology 11, no. 12: 1832. https://doi.org/10.3390/biology11121832
APA StyleWang, Y., Li, X., Zhao, X., Chen, J., Wang, Z., Chen, L., Zhang, S., & Wang, K. (2022). Assessment of Fish Diversity in the Ma’an Archipelago Special Protected Area Using Environmental DNA. Biology, 11(12), 1832. https://doi.org/10.3390/biology11121832