
Evaluation of Gastrocnemius Motor Evoked Potentials Induced by Trans-Spinal Magnetic Stimulation Following Tibial Nerve Crush in Rats
 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
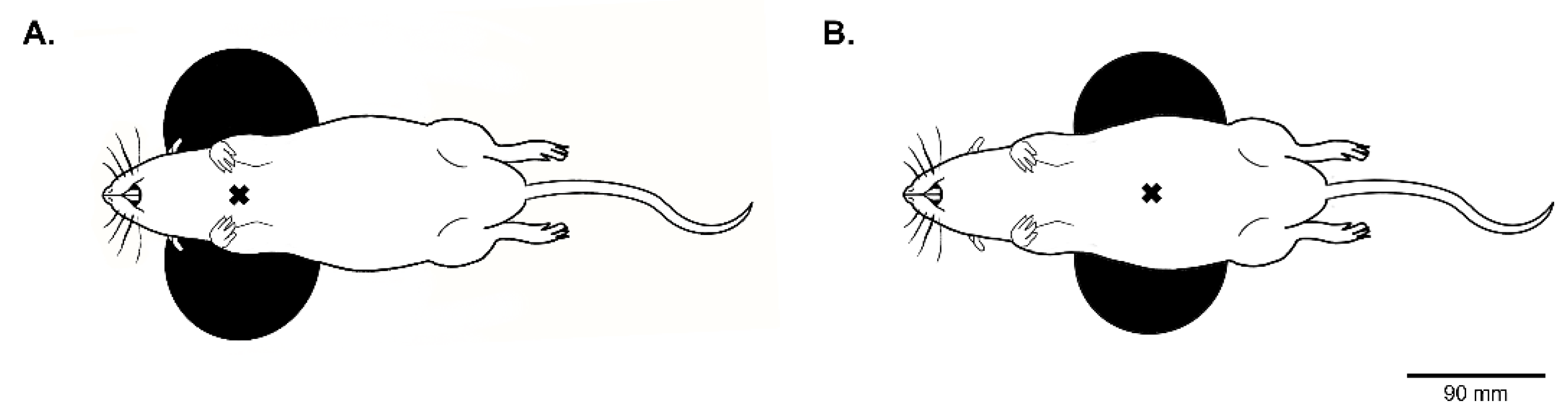
2.2. Trans-Spinal Magnetic Stimulation
2.3. Electrophysiological Recording of Gastrocnemius and Brachioradialis MEP Amplitude in Naïve Rats
2.4. Tibial Nerve (TN) Crush Surgery
2.5. Histology
2.6. Data Processing and Statistical Analyses
3. Results
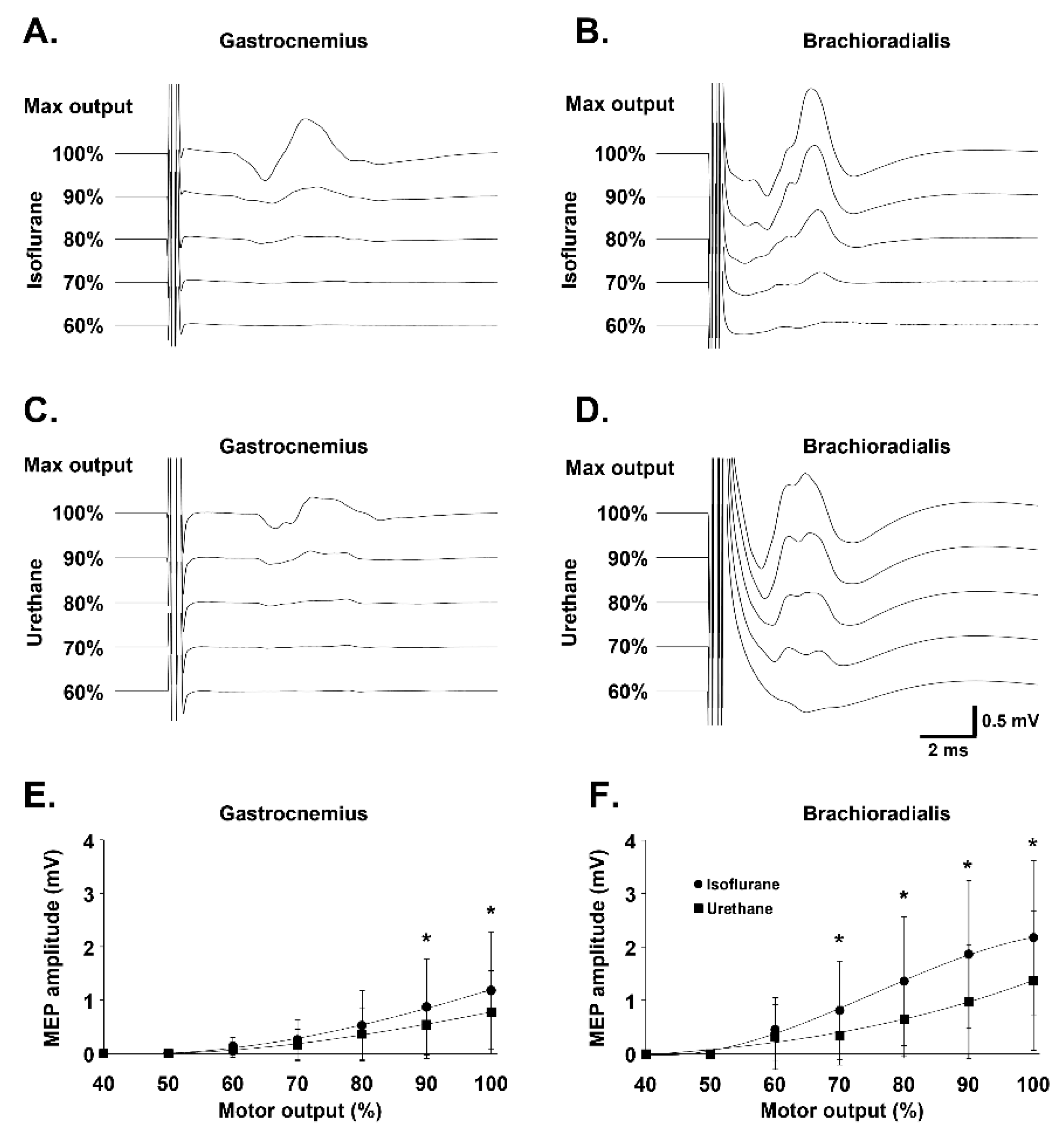
3.1. Effect of Anesthesia on Gastrocnemius and Brachioradialis MEP Amplitude
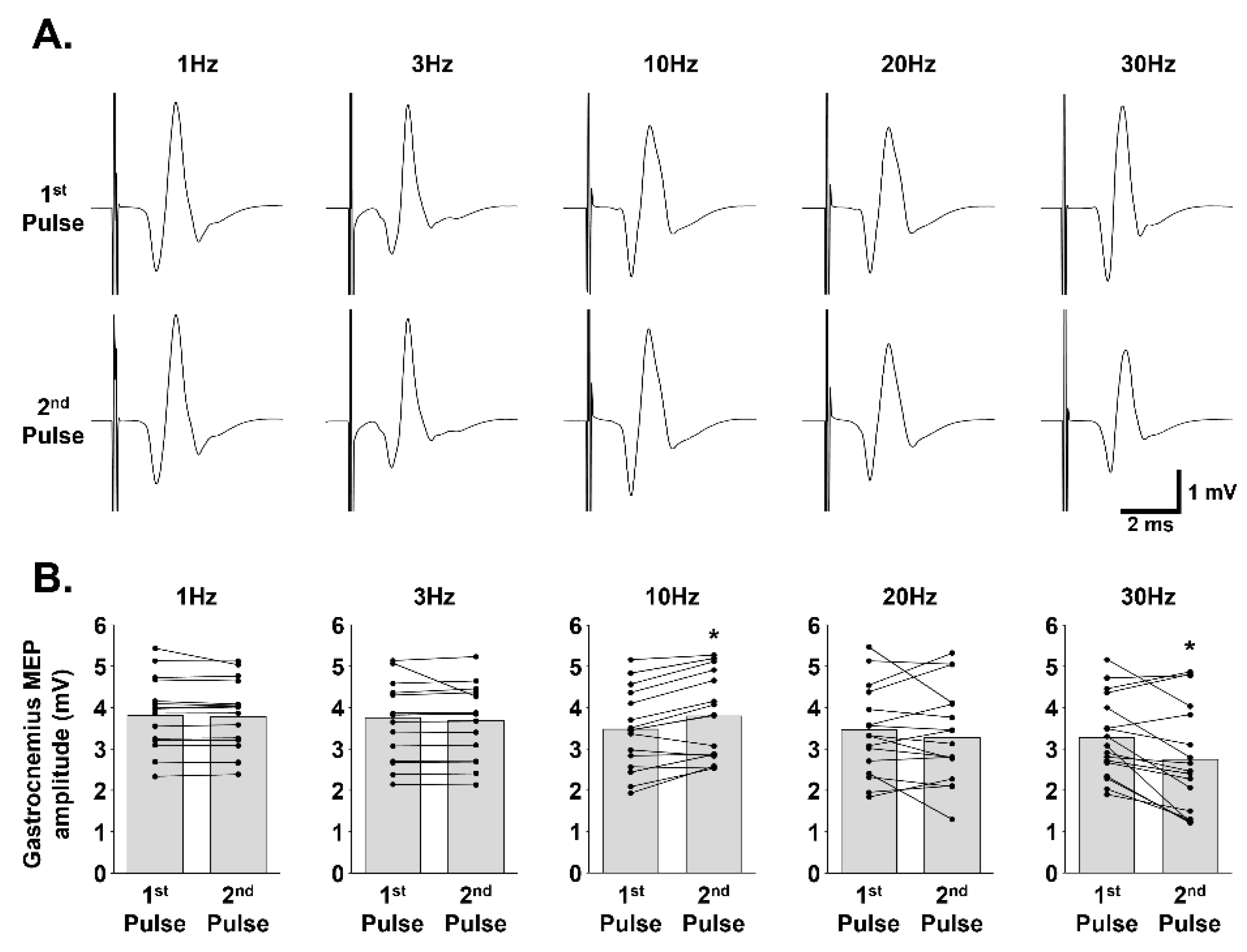
3.2. Effect of Paired-Pulse Stimulation at Different Frequencies on Gastrocnemius MEP Amplitude
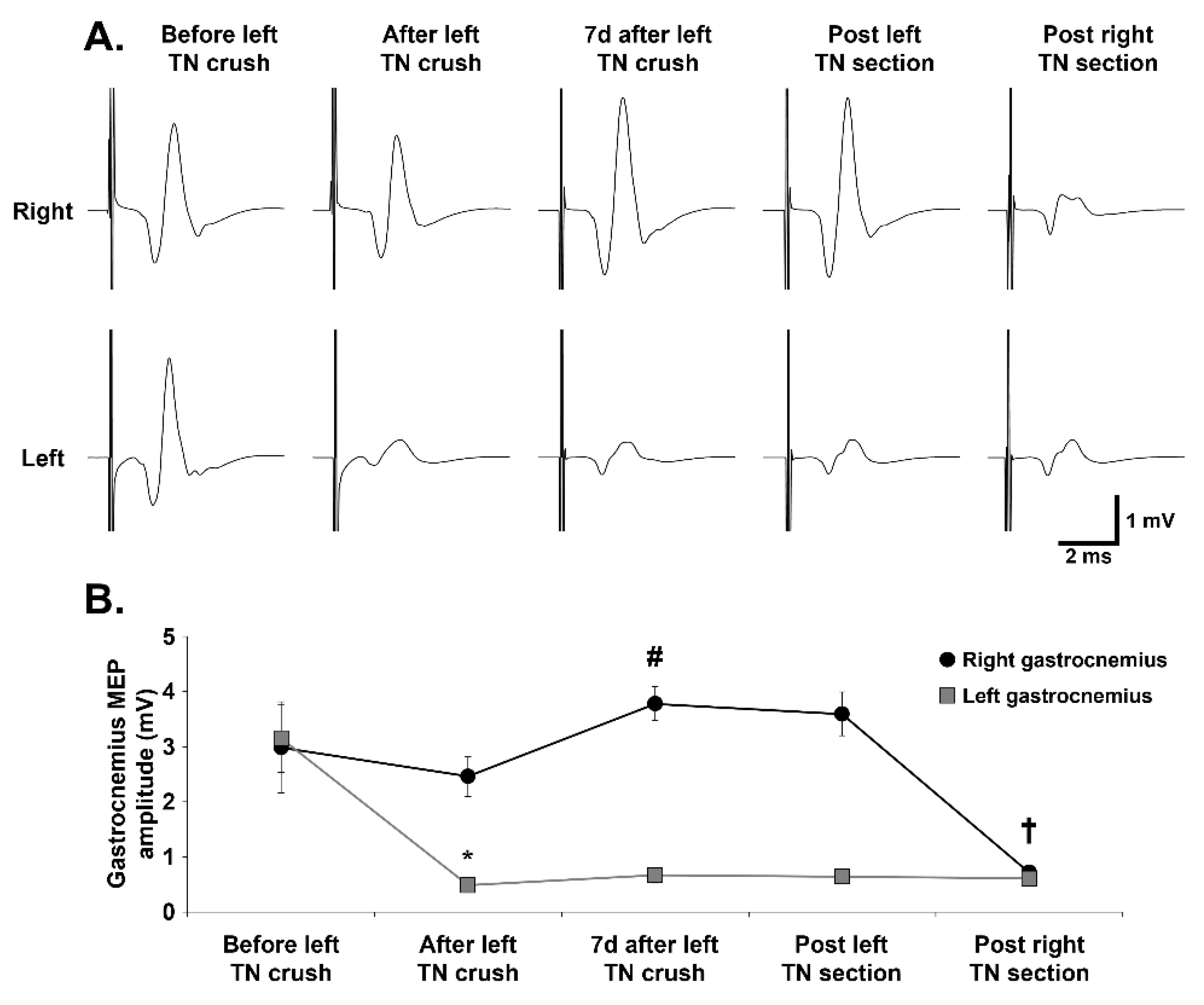
3.3. Evolution of Gastrocnemius MEPs over Time Following TN Crush and Section
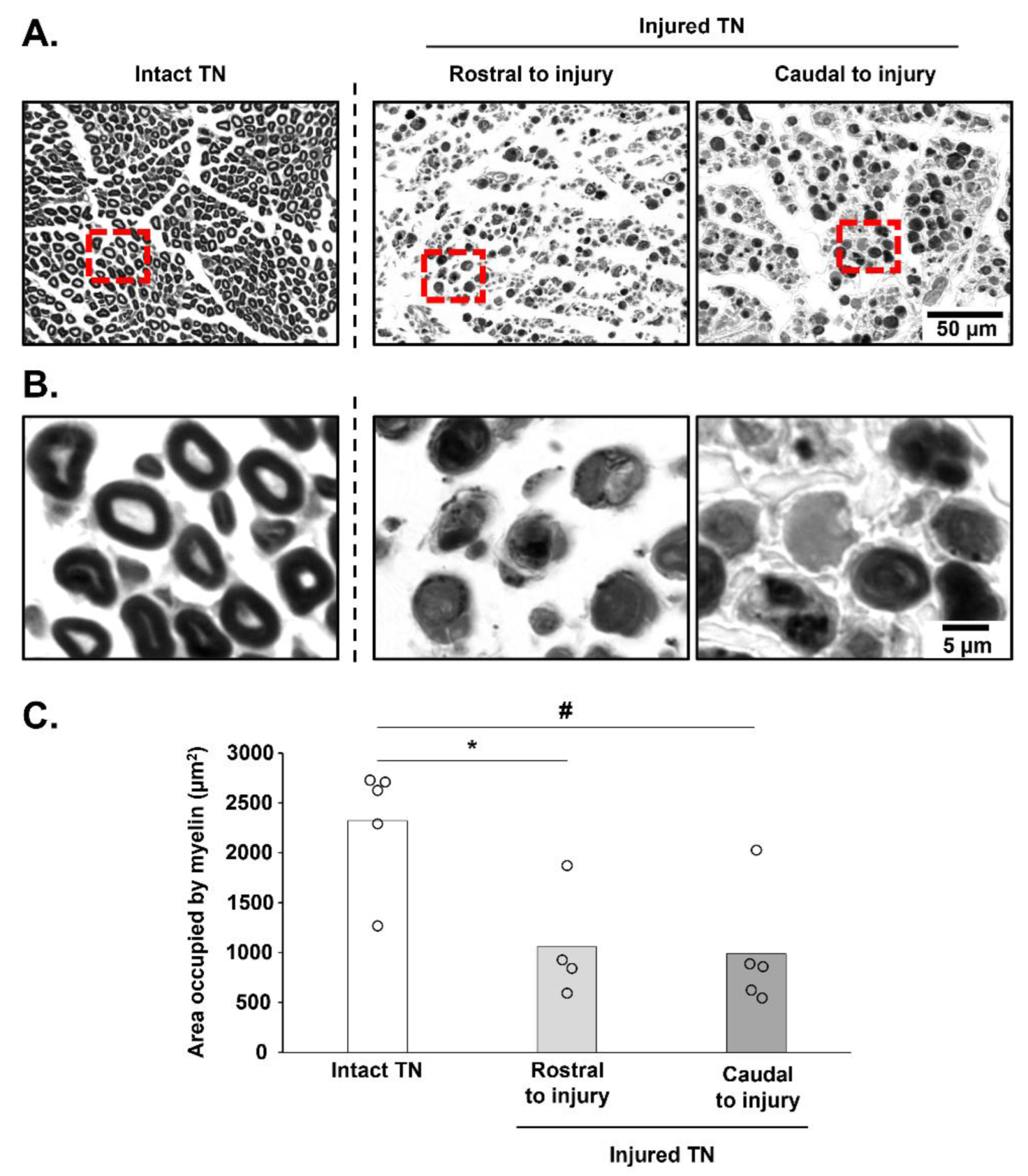
3.4. Histological Assessment of TN Integrity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antoniadis, G.; Kretschmer, T.; Pedro, M.T.; König, R.W.; Heinen, C.P.; Richter, H.P. Iatrogenic nerve injuries: Prevalence, diagnosis and treatment. Dtsch. Arztebl. Int. 2014, 111, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornwall, R.; Radomisli, T.E. Nerve injury in traumatic dislocation of the hip. Clin. Orthop. Relat. Res. 2000, 377, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Burnett, M.G.; Zager, E.L. Pathophysiology of peripheral nerve injury: A brief review. Neurosurg. Focus 2004, 16, E1. [Google Scholar] [CrossRef]
- Kurtoglu, Z.; Ozturk, A.H.; Bagdatoglu, C.; Polat, G.; Aktekin, M.; Uzmansel, D.; Camdeviren, H.; Bagdatoglu, O.; Sargon, M. Effects of trapidil after crush injury in peripheral nerve. Acta Med. Okayama 2005, 59, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Lundborg, G. A 25-year perspective of peripheral nerve surgery: Evolving neuroscientific concepts and clinical significance. J. Hand Surg. Am. 2000, 25, 391–414. [Google Scholar] [CrossRef]
- Alvites, R.; Rita Caseiro, A.; Santos Pedrosa, S.; Vieira Branquinho, M.; Ronchi, G.; Geuna, S.; Varejão, A.S.P.; Colette Maurício, A. Peripheral nerve injury and axonotmesis: State of the art and recent advances. Cogent Med. 2018, 5, 1466404. [Google Scholar] [CrossRef]
- Beer, G.M.; Steurer, J.; Meyer, V.E. Standardizing nerve crushes with a non-serrated clamp. J. Reconstr. Microsurg. 2001, 17, 531–534. [Google Scholar] [CrossRef]
- Geuna, S. The sciatic nerve injury model in pre-clinical research. J. Neurosci. Methods 2015, 243, 39–46. [Google Scholar] [CrossRef]
- Savastano, L.E.; Laurito, S.R.; Fitt, M.R.; Rasmussen, J.A.; Gonzalez Polo, V.; Patterson, S.I. Sciatic nerve injury: A simple and subtle model for investigating many aspects of nervous system damage and recovery. J. Neurosci. Methods 2014, 227, 166–180. [Google Scholar] [CrossRef]
- Elsohemy, A.; Butler, R.; Bain, J.R.; Fahnestock, M. Sensory protection of rat muscle spindles following peripheral nerve injury and reinnervation. Plast. Reconstr. Surg. 2009, 124, 1860–1868. [Google Scholar] [CrossRef]
- Bhatt, N.K.; Khan, T.R.; Mejias, C.; Paniello, R.C. Nerve transection repair using laser-activated chitosan in a rat model. Laryngoscope 2017, 127, E253–E257. [Google Scholar] [CrossRef] [PubMed]
- Monte-Raso, V.V.; Barbieri, C.H.; Mazzer, N.; Yamasita, A.C.; Barbieri, G. Is the Sciatic Functional Index always reliable and reproducible? J. Neurosci. Methods 2008, 170, 255–261. [Google Scholar] [CrossRef]
- Bain, J.R.; Mackinnon, S.E.; Hunter, D.A. Functional evaluation of complete sciatic, peroneal, and posterior tibial nerve lesions in the rat. Plast. Reconstr. Surg. 1989, 83, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J.; Mackinnon, S.E.; Evans, P.J.; Bain, J.R.; Makino, A.P.; Hunter, D.A.; Hare, G.M. Self-evaluation of walking-track measurement using a Sciatic Function Index. Microsurgery 1989, 10, 226–235. [Google Scholar] [CrossRef] [PubMed]
- de Ruiter, G.C.; Spinner, R.J.; Alaid, A.O.; Koch, A.J.; Wang, H.; Malessy, M.J.; Currier, B.L.; Yaszemski, M.J.; Kaufman, K.R.; Windebank, A.J. Two-dimensional digital video ankle motion analysis for assessment of function in the rat sciatic nerve model. J. Peripher. Nerv. Syst. 2007, 12, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ito, A.; Tajino, J.; Kuroki, H.; Aoyama, T. 3D Kinematic Analysis for the Functional Evaluation in the Rat Model of Sciatic Nerve Crush Injury. J. Vis. Exp. 2020, 156, e60267. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ito, A.; Aoyama, T.; Nakahara, R.; Nakahata, A.; Ji, X.; Zhang, J.; Kawai, H.; Kuroki, H. Functional evaluation outcomes correlate with histomorphometric changes in the rat sciatic nerve crush injury model: A comparison between sciatic functional index and kinematic analysis. PLoS ONE 2018, 13, e0208985. [Google Scholar] [CrossRef]
- Dinh, P.; Hazel, A.; Palispis, W.; Suryadevara, S.; Gupta, R. Functional assessment after sciatic nerve injury in a rat model. Microsurgery 2009, 29, 644–649. [Google Scholar] [CrossRef]
- Zhu, Y.; Jin, Z.; Luo, Y.; Wang, Y.; Peng, N.; Peng, J.; Wang, Y.; Yu, B.; Lu, C.; Zhang, S. Evaluation of the Crushed Sciatic Nerve and Denervated Muscle with Multimodality Ultrasound Techniques: An Animal Study. Ultrasound Med. Biol. 2020, 46, 377–392. [Google Scholar] [CrossRef]
- Li, X.; Shen, J.; Chen, J.; Wang, X.; Liu, Q.; Liang, B. Magnetic resonance imaging evaluation of acute crush injury of rabbit sciatic nerve: Correlation with histology. Can. Assoc. Radiol. J. 2008, 59, 123–130. [Google Scholar]
- Scipio, F.D.; Raimondo, S.; Tos, P.; Geuna, S. A simple protocol for paraffin-embedded myelin sheath staining with osmium tetroxide for light microscope observation. Microsc. Res. Tech. 2008, 71, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Carriel, V.; Garzón, I.; Alaminos, M.; Cornelissen, M. Histological assessment in peripheral nerve tissue engineering. Neural Regen. Res. 2014, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- David, G.K.; Earl, R.H.; Peter, R.M. Evaluation of Nerve Injuries by Evoked Potentials and Electromyography. J. Neurosurg. 1969, 31, 128–136. [Google Scholar] [CrossRef]
- Mathews, K.S.; Wark, H.A.; Normann, R.A. Assessment of rat sciatic nerve function following acute implantation of high density Utah slanted electrode array (25 electrodes/mm2) based on neural recordings and evoked muscle activity. Muscle Nerve 2014, 50, 417–424. [Google Scholar] [CrossRef] [PubMed]
- English, A.W.; Chen, Y.; Carp, J.S.; Wolpaw, J.R.; Chen, X.Y. Recovery of electromyographic activity after transection and surgical repair of the rat sciatic nerve. J. Neurophysiol. 2007, 97, 1127–1134. [Google Scholar] [CrossRef]
- Qu, X.; Yan, J.; Li, X.; Zhang, P.; Liu, X. Topography of Synchronization of Somatosensory Evoked Potentials Elicited by Stimulation of the Sciatic Nerve in Rat. Front. Comput. Neurosci. 2016, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Delaere, O.; Guerit, J.M.; Van Wijck, R. Sciatic nerve regeneration in the rat after frozen muscle grafting. A comparative study using somatosensory evoked potentials. J. Hand Surg. Br. 1996, 21, 53–56. [Google Scholar] [CrossRef]
- Vinit, S.; Keomani, E.; Deramaudt, T.B.; Spruance, V.M.; Bezdudnaya, T.; Lane, M.A.; Bonay, M.; Petitjean, M. Interdisciplinary approaches of transcranial magnetic stimulation applied to a respiratory neuronal circuitry model. PLoS ONE 2014, 9, e113251. [Google Scholar] [CrossRef]
- Vinit, S.; Keomani, E.; Deramaudt, T.B.; Bonay, M.; Petitjean, M. Reorganization of Respiratory Descending Pathways following Cervical Spinal Partial Section Investigated by Transcranial Magnetic Stimulation in the Rat. PLoS ONE 2016, 11, e0148180. [Google Scholar] [CrossRef] [Green Version]
- Hiwaki, O.; Ueno, S. Selective Magnetic Stimulation Of The Spinal Cord. In Proceedings of the [1990] Twelfth Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Philadelphia, PA, USA, 1–4 November 1990; pp. 436–437. [Google Scholar]
- Linden, R.D.; Zhang, Y.P.; Burke, D.A.; Hunt, M.A.; Harpring, J.E.; Shields, C.B. Magnetic motor evoked potential monitoring in the rat. J. Neurosurg. 1999, 91, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Sykes, M.; Matheson, N.A.; Brownjohn, P.W.; Tang, A.D.; Rodger, J.; Shemmell, J.B.; Reynolds, J.N. Differences in Motor Evoked Potentials Induced in Rats by Transcranial Magnetic Stimulation under Two Separate Anesthetics: Implications for Plasticity Studies. Front. Neural Circuits 2016, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.D.; Lowe, A.S.; Garrett, A.R.; Woodward, R.; Bennett, W.; Canty, A.J.; Garry, M.I.; Hinder, M.R.; Summers, J.J.; Gersner, R.; et al. Construction and Evaluation of Rodent-Specific rTMS Coils. Front. Neural Circuits 2016, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, R.; Tosolini, A.P.; Morris, R. Segmental distribution of the motor neuron columns that supply the rat hindlimb: A muscle/motor neuron tract-tracing analysis targeting the motor end plates. Neuroscience 2015, 307, 98–108. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luft, A.R.; Kaelin-Lang, A.; Hauser, T.-K.; Buitrago, M.M.; Thakor, N.V.; Hanley, D.F.; Cohen, L.G. Modulation of rodent cortical motor excitability by somatosensory input. Exp. Brain Res. 2002, 142, 562–569. [Google Scholar] [CrossRef]
- Vahabzadeh-Hagh, A.M.; Muller, P.A.; Pascual-Leone, A.; Jensen, F.E.; Rotenberg, A. Measures of cortical inhibition by paired-pulse transcranial magnetic stimulation in anesthetized rats. J. Neurophysiol. 2011, 105, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Kamida, T.; Fujiki, M.; Hori, S.; Isono, M. Conduction pathways of motor evoked potentials following transcranial magnetic stimulation: A rodent study using a “figure-8” coil. Muscle Nerve 1998, 21, 722–731. [Google Scholar] [CrossRef]
- Rotenberg, A.; Muller, P.A.; Vahabzadeh-Hagh, A.M.; Navarro, X.; López-Vales, R.; Pascual-Leone, A.; Jensen, F. Lateralization of forelimb motor evoked potentials by transcranial magnetic stimulation in rats. Clin. Neurophysiol. 2010, 121, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.H.; Huang, Y.Z.; Rotenberg, A.; Pascual-Leone, A.; Chiang, Y.H.; Wang, J.Y.; Chen, J.J. Functional Dopaminergic Neurons in Substantia Nigra are Required for Transcranial Magnetic Stimulation-Induced Motor Plasticity. Cereb. Cortex 2015, 25, 1806–1814. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.D. Anaesthetic modulation of synaptic transmission in the mammalian CNS. Br. J. Anaesth. 2002, 89, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Hara, K.; Harris, R.A. The Anesthetic Mechanism of Urethane: The Effects on Neurotransmitter-Gated Ion Channels. Anesth. Analg. 2002, 94, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Vogler, G.A. Chapter 19—Anesthesia and Analgesia. In The Laboratory Rat, 2nd ed.; Suckow, M.A., Weisbroth, S.H., Franklin, C.L., Eds.; Academic Press: Burlington, MA, USA, 2006; pp. 627–664. [Google Scholar]
- Engel, A.G. The neuromuscular junction. Handb. Clin. Neurol. 2008, 91, 103–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefaucheur, J.P. Transcranial magnetic stimulation. Handb. Clin. Neurol. 2019, 160, 559–580. [Google Scholar] [CrossRef] [PubMed]
- Van der Kloot, W.; Molgó, J. Quantal acetylcholine release at the vertebrate neuromuscular junction. Physiol. Rev. 1994, 74, 899–991. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, J.I. Repetitive Stimulation at the Mammalian Neuromuscular Junction, and the Mobilization of Transmitter. J. Physiol. 1963, 169, 641–662. [Google Scholar] [CrossRef] [Green Version]
- Lozeron, P.; Krarup, C.; Schmalbruch, H. Regeneration of unmyelinated and myelinated sensory nerve fibres studied by a retrograde tracer method. J. Neurosci. Methods 2004, 138, 225–232. [Google Scholar] [CrossRef]
- Derderian, C.; Tadi, P. Physiology, Withdrawal Response; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Eremeev, A.; Pleshchinskii, I.; Baltina, T.; Eremeev, A. The State of the Contralateral Gastrocnemius Muscle Motor Center in Rats with Unilateral Sciatic Nerve Injury. Neurosci. Behav. Physiol. 2012, 42, 822–827. [Google Scholar] [CrossRef]
- Lee, K.Z.; Liou, L.M.; Vinit, S. Diaphragm Motor-Evoked Potential Induced by Cervical Magnetic Stimulation following Cervical Spinal Cord Contusion in the Rat. J. Neurotrauma 2021, 38, 2122–2140. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Onset Latency (ms) | ||
|---|---|---|
| Anesthetic Used (90% Maximum Output) | Gastrocnemius | Brachioradialis |
| Isoflurane | 3.08 ± 0.63 | 1.71 ± 0.31 * |
| Urethane | 3.45 ± 0.92 | 1.71 ± 0.42 * |
| Onset Latency (ms) | ||
|---|---|---|
| Time Post TN Crush | Right Gastrocnemius | Left Gastrocnemius |
| Before left TN crush | 2.40 ± 0.29 | 2.42 ± 0.14 |
| After left TN crush | 2.45 ± 0.18 | 2.36 ± 0.06 |
| 7d after left TN crush | 2.37 ± 0.34 | 2.17 ± 0.12 |
| Post-left TN section | 2.29 ± 0.19 | 2.23 ± 0.16 |
| Post-right TN section | 2.21 ± 0.20 | 2.29 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michel-Flutot, P.; Jesus, I.; Mansart, A.; Bonay, M.; Lee, K.-Z.; Auré, K.; Vinit, S. Evaluation of Gastrocnemius Motor Evoked Potentials Induced by Trans-Spinal Magnetic Stimulation Following Tibial Nerve Crush in Rats. Biology 2022, 11, 1834. https://doi.org/10.3390/biology11121834
Michel-Flutot P, Jesus I, Mansart A, Bonay M, Lee K-Z, Auré K, Vinit S. Evaluation of Gastrocnemius Motor Evoked Potentials Induced by Trans-Spinal Magnetic Stimulation Following Tibial Nerve Crush in Rats. Biology. 2022; 11(12):1834. https://doi.org/10.3390/biology11121834
Chicago/Turabian StyleMichel-Flutot, Pauline, Isley Jesus, Arnaud Mansart, Marcel Bonay, Kun-Ze Lee, Karine Auré, and Stéphane Vinit. 2022. "Evaluation of Gastrocnemius Motor Evoked Potentials Induced by Trans-Spinal Magnetic Stimulation Following Tibial Nerve Crush in Rats" Biology 11, no. 12: 1834. https://doi.org/10.3390/biology11121834
APA StyleMichel-Flutot, P., Jesus, I., Mansart, A., Bonay, M., Lee, K. -Z., Auré, K., & Vinit, S. (2022). Evaluation of Gastrocnemius Motor Evoked Potentials Induced by Trans-Spinal Magnetic Stimulation Following Tibial Nerve Crush in Rats. Biology, 11(12), 1834. https://doi.org/10.3390/biology11121834