
Phylogeny and Historical Biogeography of Veronica Subgenus Pentasepalae (Plantaginaceae): Evidence for Its Origin and Subsequent Dispersal



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction, Amplification and Sequencing
2.3. Sequence Alignment, Phylogenetic Reconstruction and Dating
2.4. Ancestral Area Reconstruction
2.5. Chromosome Counting
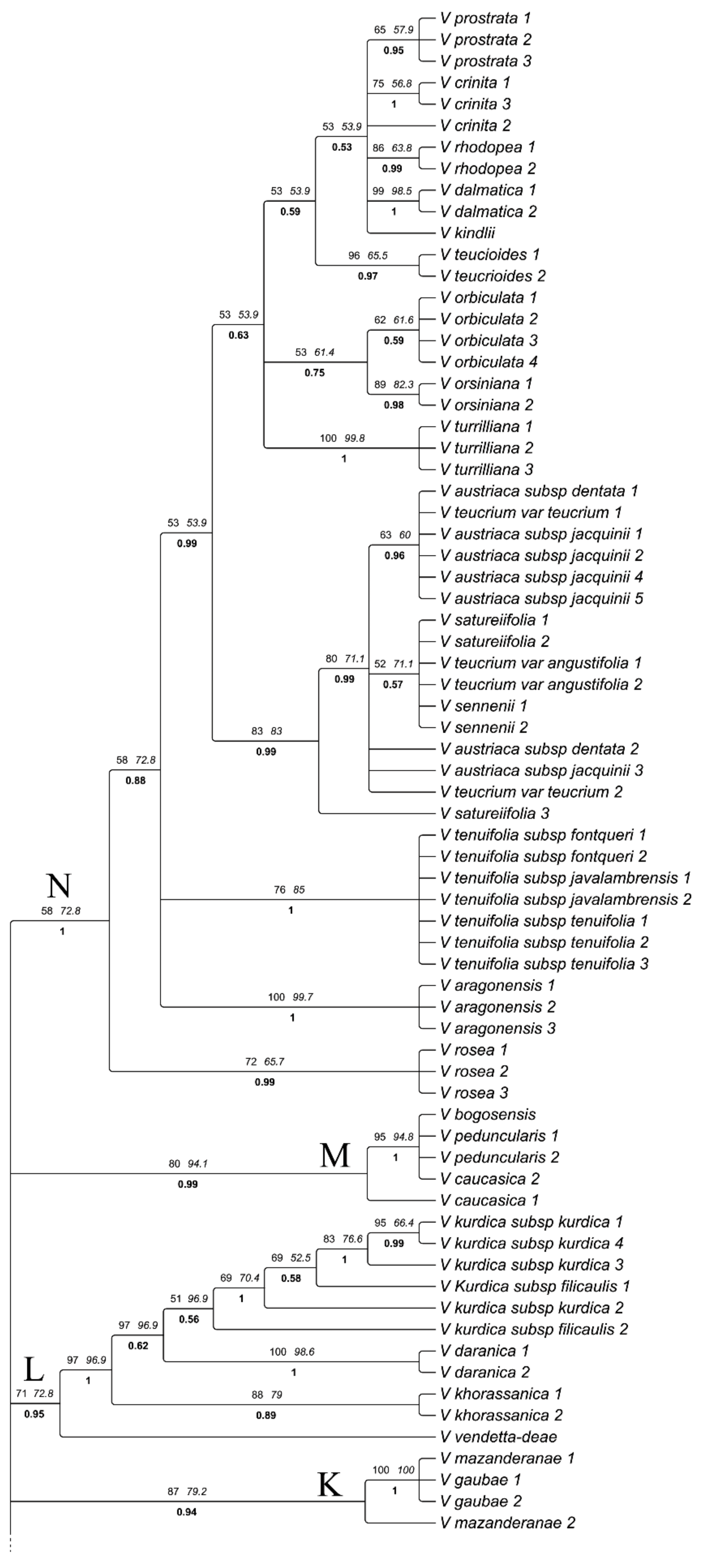
3. Results
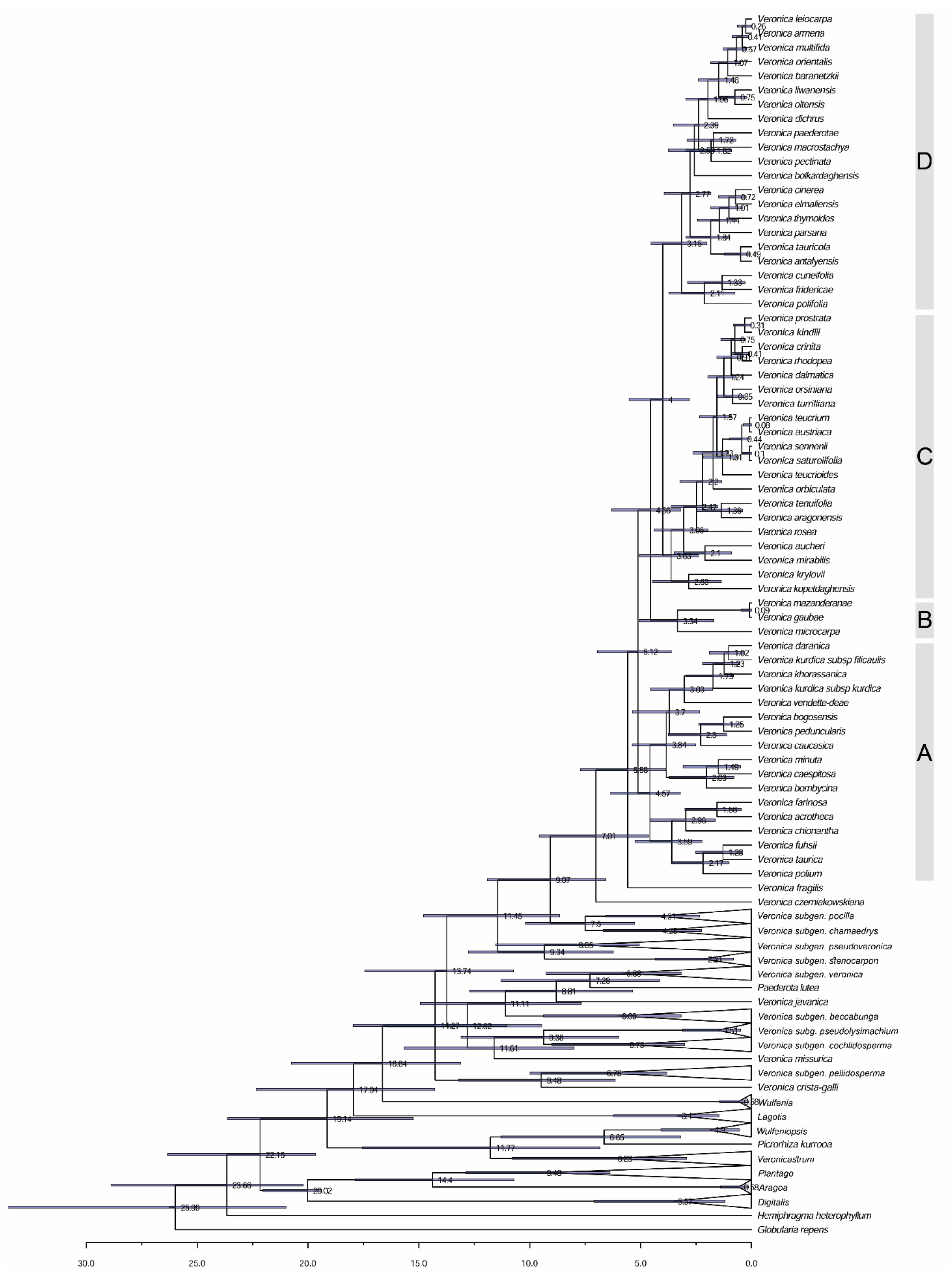
3.1. Phylogenetic Analyses and Divergence Time Estimates
3.2. Historical Biogeography Reconstructions
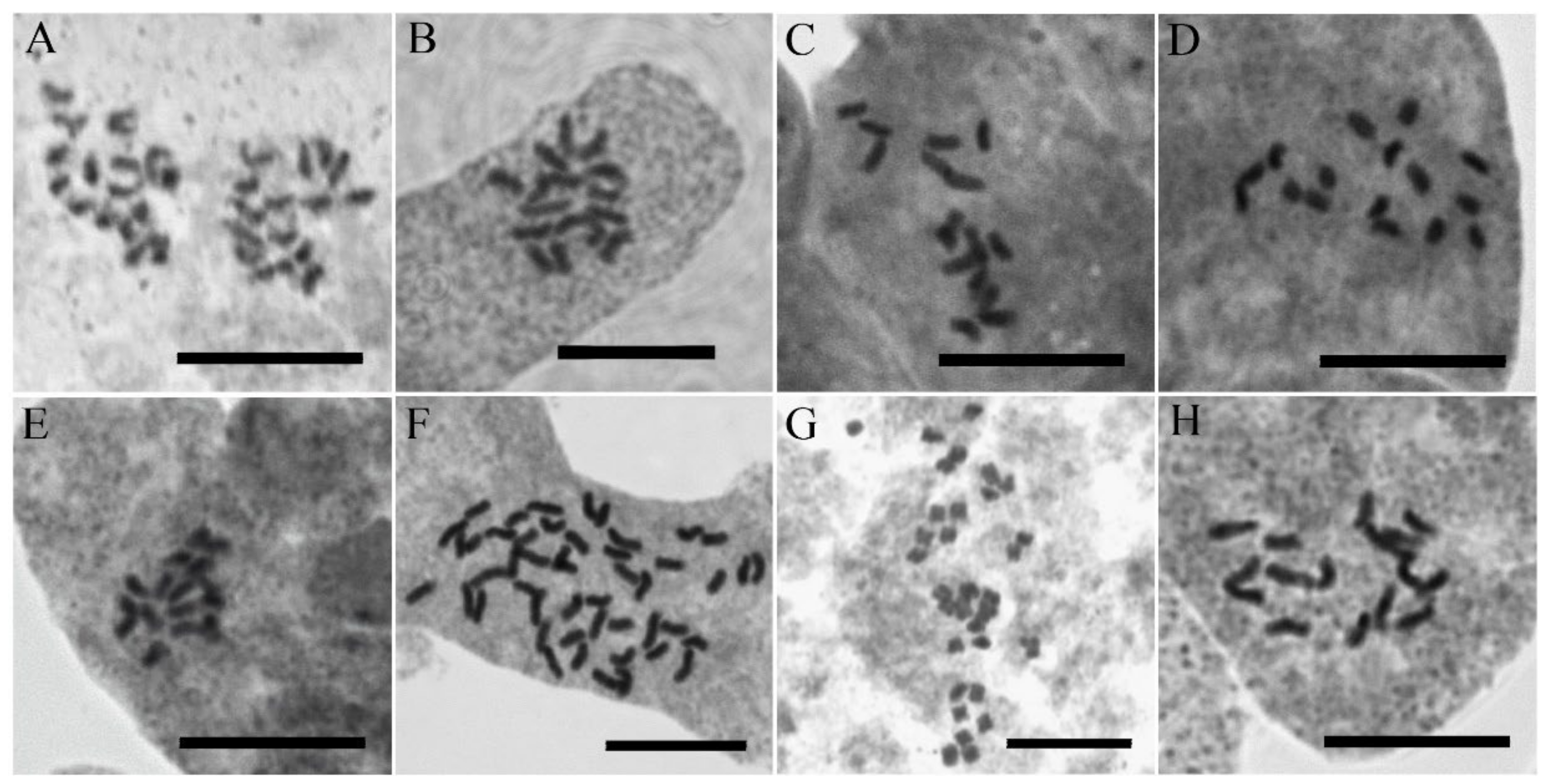
3.3. Chromosome Numbers
4. Discussion
4.1. Relict Species of Veronica in the Irano-Turanian Region with No Immediate Relatives
4.2. Phylogeny and Systematics
4.3. Origin of V. Subgenus Pentasepalae: Out of the Iranian Plateau
4.4. Dispersal and Vicariance
4.5. Taxonomic Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Vinyallonga, S.; Mehregan, I.; Garcia-Jacas, N.; Tscherneva, O.; Susanna, A.; Kadereit, J.W. Phylogeny and Evolution of the Arctium-Cousinia Complex (Compositae, Cardueae-Carduinae). Taxon 2009, 58, 153–171. [Google Scholar] [CrossRef]
- Azani, N.; Bruneau, A.; Wojciechowski, M.F.; Zarre, S. Miocene climate change as a driving force for multiple origins of annual species in Astragalus (Fabaceae, Papilionoideae). Mol. Phylogenet. Evol. 2019, 137, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Moharrek, F.; Sanmartín, I.; Kazempour-Osaloo, S.; Nieto Feliner, G. Morphological Innovations and Vast Extensions of Mountain Habitats Triggered Rapid Diversification Within the Species-Rich Irano-Turanian Genus Acantholimon (Plumbaginaceae). Front. Genet. 2019, 9, 698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabeler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Pahlevani, A.H.; Liede-Schumann, S.; Akhani, H. Diversity, distribution, endemism and conservation status of Euphorbia (Euphorbiaceae) in SW Asia and adjacent countries. Plant Syst. Evol. 2020, 306, 80. [Google Scholar] [CrossRef]
- Djamali, M.; Baumel, A.; Brewer, S.; Jackson, S.T.; Kadereit, J.W.; López-Vinyallonga, S.; Mehregan, I.; Shabanian, E.; Simakova, A. Ecological implications of Cousinia Cass. (Asteraceae) persistence through the last two glacial–interglacial cycles in the continental Middle East for the Irano-Turanian flora. Rev. Palaeobot. Palyno. 2012, 172, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Mohammadin, S.; Peterse, K.; van de Kerke, S.J.; Chatrou, L.W.; Dönmez, A.A.; Mummenhoff, K.; Pires, J.C.; Edger, P.P.; Al-Shehbaz, I.A.; Schranz, M.E. Anatolian origins and diversification of Aethionema, the sister lineage of the core Brassicaceae. Am. J. Bot. 2017, 104, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Karl, R.; Koch, M.A. A world-wide perspective on crucifer speciation and evolution: Phylogenetics, biogeography and trait evolution in tribe Arabideae. Ann. Bot. 2013, 112, 983–1001. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.L.; Sanderson, S.C.; Sun, Y.X.; Byalt, V.V.; Hao, X.L. Tertiary montane origin of the Central Asian flora, evidence inferred from cpDNA sequences of Atraphaxis (Polygonaceae). J. Integr. Plant Boil. 2014, 56, 1125–1135. [Google Scholar] [CrossRef]
- Zhang, M.L.; Wen, Z.B.; Fritsch, P.W.; Sanderson, S.C. Spatiotemporal Evolution of Calophaca (Fabaceae) Reveals Multiple Dispersals in Central Asian Mountains. PLoS ONE 2015, 10, e0123228. [Google Scholar] [CrossRef]
- Font, M.; Garcia-Jacas, N.; Vilatersana, R.; Roquet, C.; Susanna, A. Evolution and biogeography of Centaurea section Acrocentron inferred from nuclear and plastid DNA sequence analyses. Ann. Bot. 2009, 103, 985–997. [Google Scholar] [CrossRef]
- Panahi, M. Biogeographic reconstruction of the genus Ferula inferred from analyses of nrDNA and cpDNA sequences. Iran. J. Bot. 2019, 25, 79–94. [Google Scholar] [CrossRef]
- Eslami-Farouji, A.; Khodayari, H.; Assadi, M.; Çetin, Ö.; Mummenhoff, K.; Özüdoğru, B. Phylogeny and biogeography of the genus Hesperis (Brassicaceae, tribe Hesperideae) inferred from nuclear ribosomal DNA sequence data. Plant Syst. Evol. 2021, 307, 17. [Google Scholar] [CrossRef]
- Zhang, M.L.; Zeng, X.Q.; Sanderson, S.C.; Byalt, V.V.; Sukhorukov, A.P. Insight into Central Asian flora from the Cenozoic Tianshan montane origin and radiation of Lagochilus (Lamiaceae). PLoS ONE 2017, 12, e0178389. [Google Scholar] [CrossRef] [Green Version]
- Soreng, R.J.; Gillespie, L.J.; Boudko, E.A.; Cabi, E. Biogeography, Timing, and Life History Traits in the PPAM clade: Coleanthinae (syn. Puccinelliinae), Poinae, Alopecurinae superclade, Miliinae, and Avenulinae and Phleinae (Poaceae, Pooideae, Poeae). J. Syst. Evol. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Scheunert, A.; Heubl, G. Against all odds: Reconstructing the evolutionary history of Scrophularia (Scrophulariaceae) despite high levels of incongruence and reticulate evolution. Org. Divers. Evol. 2017, 17, 323–349. [Google Scholar] [CrossRef]
- Žerdoner Čalasan, A.; German, D.A.; Hurka, H.; Neuffer, B. A story from the Miocene: Clock-dated phylogeny of Sisymbrium L. (Sisymbrieae, Brassicaceae). Ecol. Evol. 2021, 11, 2573–2595. [Google Scholar] [CrossRef]
- Manafzadeh, S.; Salvo, G.; Conti, E.A. tale of migrations from east to west: The Irano-Turanian floristic region as a source of Mediterranean xerophytes. J. Biogeogr. 2014, 41, 366–379. [Google Scholar] [CrossRef]
- Malik, S.; Vitales, D.; Qasim Hayat, M.; Korobkov, A.A.; Garnatje, T.; Vallès, J. Phylogeny and biogeography of Artemisia subgenus Seriphidium (Asteraceae: Anthemideae). Taxon 2017, 66, 934–952. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.; Harpke, D.; Peterson, J.; Harpke, A.; Peruzzi, L. A pre-Miocene Irano-Turanian cradle: Origin and diversification of the species-rich monocot genus Gagea (Liliaceae). Ecol. Evol. 2019, 9, 5870–5890. [Google Scholar] [CrossRef] [Green Version]
- Albach, D.C.; Martínez-Ortega, M.M.; Chase, M.W. Veronica: Parallel morphological evolution and phylogeography in the Mediterranean. Plant Syst. Evol. 2004, 246, 177–194. [Google Scholar] [CrossRef]
- Angiosperm Phylogeny Group III. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Angiosperm phylogeny Group IV. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Fischer, E. Scrophulariaceae. In The Families and Genera of Vascular Plants; Kadereit, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 7, pp. 333–433. [Google Scholar]
- Albach, D.C.; Martínez-Ortega, M.M.; Fischer, M.A.; Chase, M.W. A new classification of the tribe Veroniceae—problems and a possible solution. Taxon 2004, 53, 429–452. [Google Scholar] [CrossRef]
- Garnock-Jones, P.; Albach, D.C.; Briggs, B.G. Botanical names in Southern Hemisphere Veronica (Plantaginaceae): Sect. Detzneria, sect. Hebe, and sect. Labiatoides. Taxon 2007, 56, 571–582. [Google Scholar] [CrossRef]
- Albach, D.C.; Martínez-Ortega, M.M.; Delgado, L.; Weiss-Schneeweiss, H.; Özgökce, F.; Fischer, M.A. Chromosome Numbers in Veroniceae (Plantaginaceae): Review and Several New Counts. Ann. Missouri Bot. Gard. 2008, 95, 543–566. [Google Scholar] [CrossRef]
- Rojas-Andrés, B.M.; Rico, E.; Martínez-Ortega, M.M. A nomenclatural treatment for Veronica subsect. Pentasepalae (Plantaginaceae sensu APG III) and typification of several names. Taxon 2016, 65, 617–627. [Google Scholar] [CrossRef]
- Padilla-García, N.; Rojas-Andrés, B.M.; López-González, N.; Castro, M.; Castro, S.; Loureiro, J.; Albach, D.C.; Machon, N.; Martínez-Ortega, M.M. The challenge of species delimitation in the diploid-polyploid complex Veronica subsection Pentasepalae. Mol. Phylogenet. Evol. 2018, 119, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Yaylaci, Ö.K.; Sezer, O.; Özgisi, K.; Özturk, D.; Erkara, İ.P.; Koyuncu, O.; Ocak, A. A new Veronica (Plantaginaceae) species from central Anatolia, Turkey. Phytotaxa 2018, 362, 55–67. [Google Scholar] [CrossRef]
- Rojas-Andrés, B.M.; Albach, D.C.; Martínez-Ortega, M.M. Exploring the intricate evolutionary history of the diploid–polyploid complex Veronica subsection Pentasepalae (Plantaginaceae). Bot. J. Linn. Soc. 2015, 179, 670–692. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Andres, B.M.; Martínez-Ortega, M.M. Taxonomic revision of Veronica subsection Pentasepalae (Veronica, Plantaginaceae sensu APG III). Phytotaxa 2016, 285, 1–100. [Google Scholar] [CrossRef]
- Martínez-Ortega, M.M.; Delgado, L.; Albach, D.C.; Elena-Rosselló, J.A.; Rico, E. Species Boundaries and Phylogeographic Patterns in Cryptic Taxa Inferred from AFLP Markers: Veronica subgen. Pentasepalae (Scrophulariaceae) in the Western Mediterranean. Syst. Bot. 2004, 29, 965–986. [Google Scholar] [CrossRef]
- Noroozi, J.; Körner, C. A bioclimatic characterization of high elevation habitats in the Alborz mountains of Iran. Alp. Bot. 2018, 128, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Albach, D.C.; Al-Gharaibeh, M. Systematics and biogeography of Veronica subg. Pentasepalae from the Levant. Willdenowia 2015, 45, 5–14. [Google Scholar] [CrossRef]
- Sonibare, M.A.; Armagan, M.; Özgökce, F.; Yaprak, A.E.; Mayland-Quellhorst, E.; Albach, D.C. Analysis of taxonomic and geographic patterns of Turkish Veronica orientalis using nuclear and plastid DNA and morphological data. Plant Syst. Evol. 2014, 300, 645–664. [Google Scholar] [CrossRef]
- Borissova, A.G. Veronica In Flora of the U.S.S.R.; Shishkin, B.K., Bobrov, E.G., Eds.; Akademii Nauk SSSR: Moscow, Russia, 1955; Volume 22, pp. 373–557. [Google Scholar]
- Fischer, M.A. Scrophulariaceae. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Edmondson, J.R., Mill, R.R., Parris, B.S., Eds.; Edinburgh University Press: Edinburgh, UK, 1978; Volume 6, pp. 458–784. [Google Scholar]
- Fischer, M.A. Veronica. In Flora Iranica 147 (Scrophulariaceae I); Rechinger, K.H., Ed.; Akadem-ische Druck- u, Verlagsanstalt: Graz, Austria, 1981; pp. 52–165. [Google Scholar]
- Saeidi-Mehrvarz, S. Veronica. In Flora of Iran 68; Assadi, M., Maassoumi, A.A., Khatamsaz, M., Mozaffarian, V., Eds.; Research Institute of Forests and Rangelands Publications: Tehran, Iran, 2011. (In Persian) [Google Scholar]
- Albach, D.C.; Martínez-Ortega, M.M.; Fischer, M.A.; Chase, M.W. Evolution of Veroniceae: A Phylogenetic Perspective. Ann. Mssouri Bot. Gard. 2004, 91, 275–302. Available online: https://www.jstor.org/stable/3298609 (accessed on 1 December 2021).
- Surina, B.; Pfanzelt, S.; Einzmann, H.J.R.; Albach, D.C. Bridging the Alps and the Middle East: Evolution, phylogeny and systematics of the genus Wulfenia (Plantaginaceae). Taxon 2014, 63, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Meudt, H.M.; Rojas-Andrés, B.M.; Prebble, J.M.; Low, E.; Garnock-Jones, P.J.; Albach, D.C. Is genome downsizing associated with diversification in polyploid lineages of Veronica? Bot. J. Linn. Soc. 2015, 178, 243–266. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Blattner, F.R. Direct Amplification of the Entire ITS Region from Poorly Preserved Plant Material Using Recombinant PCR. BioTechniques 1999, 27, 1180–1186. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Documentation of reticulated evolution in peonies (Paeonia) using internal transcribed spacer sequences of nuclear ribosomal DNA: Implications for biogeography and concerted evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 6813–6817. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Muller, K.; Muller, J.; Quandt, D. PhyDE®: Phylogenetic Data Editor, Version 0.9971. 2010. Available online: http://www.phyde.de/index.html (accessed on 1 March 2021).
- Müller, K. SeqState: Primer design and sequence statistics for phylogenetic DNA datasets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Swafford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0.b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Nixon, K.C. The Parsimony Ratchet, a New Method for Rapid Parsimony Analysis. Cladistics 1999, 15, 407–414. [Google Scholar] [CrossRef]
- Müller, K. PRAP—computation of Bremer support for large data sets. Mol. Phylogeneti. Evol. 2004, 31, 780–782. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed Phylogenetics and Dating with Confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.7. 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 20 January 2022).
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Harris, A.J.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doostmohammadi, M.; Mirtadzadini, M.; Bordbar, F. Chromosome numbers of some annual Veronica (Plantaginaceae) with report of V. tenuissima from flora of Iran. Feddes Repert. 2021, 132, 158–166. [Google Scholar] [CrossRef]
- Valente, L.M.; Savolainen, V.; Vargas, P. Unparalleled rates of species diversification in Europe. Proc. Biol. Sci. 2010, 277, 1489–1496. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.D.; Mavrodiev, E.V.; Soltis, P.S.; Calaminus, A.K.; Albach, D.C.; Cellinese, N.; Garcia-Jacas, N.; Soltis, D.E. Rapid diversification of Tragopogon and ecological associates in Eurasia. J. Evolution. Biol. 2012, 25, 2470–2480. [Google Scholar] [CrossRef]
- Richardson, J.E.; Pennington, R.T.; Pennington, T.D.; Hollingsworth, P.M. Rapid diversification of a species-rich genus of neotropical rain forest trees. Science 2001, 293, 2242–2245. [Google Scholar] [CrossRef]
- Hughes, C.; Eastwood, R. Island radiation on a continental scale: Exceptional rates of plant diversification after uplift of the Andes. Proc. Natl. Acad. Sci. USA 2006, 103, 10334–10339. [Google Scholar] [CrossRef] [Green Version]
- Sauquet, H.; Weston, P.H.; Anderson, C.L.; Barker, N.P.; Cantrill, D.J.; Mast, A.R.; Savolainen, V. Contrasted patterns of hyperdiversification in Mediterranean hotspots. Proc. Natl. Acad. Sci. USA 2009, 106, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, A.; Maassoumi, A.A.; Rahiminejad, M.R.; Brassac, J.; Blattner, F.R. Molecular phylogeny and divergence times of Astragalus section Hymenostegis: An analysis of a rapidly diversifying species group in Fabaceae. Sci. Rep. 2017, 7, 14033. [Google Scholar] [CrossRef] [Green Version]
- Axen, G.J.; Lam, P.S.; Grove, M.; Stockli, D.F.; Hassanzadeh, J. Exhumation of the west-central Alborz Mountains, Iran, Caspian subsidence, and collision-related tectonics. Geology 2001, 29, 559–562. [Google Scholar] [CrossRef]
- Lacombe, O.; Mouthereau, F.; Kargar, S.; Meyer, B. Late Cenozoic and modern stress fields in the western Fars (Iran): Implications for the tectonic and kinematic evolution of central Zagros. Tectonics 2006, 25, TC1003. [Google Scholar] [CrossRef] [Green Version]
- Mouthereau, F.; Lacombe, O.; Vergés, J. Building the Zagros collisional orogen: Timing, strain distribution and the dynamics of Arabia/Eurasia plate convergence. Tectonophysics 2012, 532–535, 27–60. [Google Scholar] [CrossRef]
- Padilla-García, N.; Machon, N.; Segarra-Moragues, J.G.; Martínez-Ortega, M.M. Surviving in southern refugia: The case of Veronica aragonensis, a rare endemic from the Iberian Peninsula. Alp. Bot. 2021, 131, 161–175. [Google Scholar] [CrossRef]
- Boissier, E. Diagnoses Plantarum Orientalum Novarum; B. Hermann: Leipzig, Germany, 1844. [Google Scholar]
- Bentham, G. Scrophulariaceae. In Prodromus Systematis Naturalis Regni Vegetabilis; de Candolle, A., Ed.; Victor Masson: Paris, France, 1846; Volume 10, pp. 448–491. [Google Scholar]
- Römpp, H. Die Verwandtschaftsverhältnisse in der Gattung Veronica. Repert. Spec. Nov. Regni Veg. 1928, 50, 1–171. [Google Scholar]
- Bornmüller, J.; Gauba, E. Florae Keredjensis fundamenta (Plantae Gaubaeanae Iranica). Supplementum. Repert. Spec. Nov. Regni Veg. 1942, 51, 209–235. [Google Scholar]
- Elenevsky, A.G. Sistematika i Geografiya Veronik SSSR i Prilezhashchikh Stran; Nauka: Moscow, Russia, 1978. [Google Scholar]
- Freyn, J. Ueber neue und bemerkenswerte orientalische Pflanzenarten (suite et fin). Bull. Herb. Boissier 1897, 5, 781–804. [Google Scholar]
- Parsa, A. Veronica L. in Flore de l’Iran; Publications du Ministere de L’Education: Teheran, Iran, 1949; Volume 4, pp. 423–466. [Google Scholar]
- Saeidi-Mehrvarz, S.; Ghahreman, A.; Assadi, M. A new species of Veronica (Scrophulariaceae) from Iran. Nord. J. Bot. 2001, 21, 581–584. [Google Scholar] [CrossRef]
- Fischer, M.A. Veronica fridericae, a new species from Eastern Turkey. Plant Syst. Evol. 1984, 144, 67–71. [Google Scholar] [CrossRef]
- Miao, Y.; Herrmann, M.; Wu, F.; Yan, X.; Yang, S. What controlled Mid–Late Miocene long-term aridification in Central Asia?—Global cooling or Tibetan Plateau uplift: A review. Earth Sci. Rev. 2012, 112, 155–172. [Google Scholar] [CrossRef]
- Kadereit, J.W.; Licht, W.; Uhink, C.H. Asian relationships of the flora of the European Alps. Plant Ecol. Divers. 2008, 1, 171–179. [Google Scholar] [CrossRef]
- Zhang, M.; Kang, Y.; Zhou, L.; Podlech, D. Phylogenetic origin of Phyllolobium with a further implication for diversification of Astragalus in China. J. Integr. Plant Boil. 2009, 51, 889–899. [Google Scholar] [CrossRef]
- Xu, T.; Abbott, R.J.; Milne, R.I.; Mao, K.; Du, F.K.; Wu, G.; Ciren, Z.; Miehe, G.; Liu, J. Phylogeography and allopatric divergence of cypress species (Cupressus L.) in the Qinghai-Tibetan Plateau and adjacent regions. BMC Evol. Biol. 2010, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.R.; Abbott, R.J.; Liu, T.L.; Mao, K.S.; Bartish, I.V.; Liu, J.Q. Out of the Qinghai-Tibet Plateau: Evidence for the origin and dispersal of Eurasian temperate plants from a phylogeographic study of Hippophaë rhamnoides (Elaeagnaceae). New Phytol. 2012, 194, 1123–1133. [Google Scholar] [CrossRef]
- Li, G.D.; Kim, C.; Zha, H.G.; Zhou, Z.; Nie, Z.L.; Sun, H. Molecular phylogeny and biogeography of the arctic-alpine genus Lagotis (Plantaginaceae). Taxon 2014, 63, 103–111. [Google Scholar] [CrossRef]
- Buhk, N.; Zhao, L.; Li, H.; Albach, D.C. Molecular systematics and morphometrics in Veronica subsect. Canae (Plantaginaceae). Plant Syst. Evol. 2015, 301, 1967–1979. [Google Scholar] [CrossRef]
- Huang, J.; Yang, L.Q.; Yu, Y.; Liu, Y.M.; Xie, D.F.; Li, J.; He, X.J.; Zhou, S.D. Molecular phylogenetics and historical biogeography of the tribe Lilieae (Liliaceae): Bi-directional dispersal between biodiversity hotspots in Eurasia. Ann. Bot. 2018, 122, 1245–1262. [Google Scholar] [CrossRef]
- Sherafati, M.; Kazempour-Osaloo, S.; Khoshsokhan-Mozaffar, M.; Esmailbegi, S.; Staedler, Y.M.; Manafzadeh, S. Diversification of Cynoglossinae genera (Cynoglosseae-Boraginaceae) in the western Irano–Turanian bioregion. Botany 2021, 99, 541–553. [Google Scholar] [CrossRef]
- Mittermeier, R.; Gil, P.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.; Lamoreux, J.; Da Fonseca, G. Hotspots Revisited: Earths Biologically Richest and Most Endangered Terrestrial Ecoregions; Cemex 315; Conservation International: Sierra Madre, CA, USA, 2005. [Google Scholar]
- Noroozi, J.; Talebi, A.; Doostmohammadi, M.; Rumpf, S.B.; Linder, H.P.; Schneeweiss, G.M. Hotspots within a global biodiversity hotspot-areas of endemism are associated with high mountain ranges. Sci. Rep. 2018, 8, 10345. [Google Scholar] [CrossRef] [Green Version]
- Noroozi, J.; Talebi, A.; Doostmohammadi, M.; Manafzadeh, S.; Asgarpour, Z.; Schneeweiss, G.M. Endemic diversity and distribution of the Iranian vascular flora across phytogeographical regions, biodiversity hotspots and areas of endemism. Sci. Rep. 2019, 9, 12991. [Google Scholar] [CrossRef] [Green Version]
- Noroozi, J.; Akhani, H.; Breckle, S.W. Biodiversity and phytogeography of the alpine flora of Iran. Biodivers. Conserv. 2008, 17, 493–521. [Google Scholar] [CrossRef]
- Doostmohammadi, M.; Talebi, A.; Noroozi, J. The Yazd–Kerman Massifs. In Plant Biogeography and Vegetation of High Mountains of Central and South-West Asia; Noroozi, J., Ed.; Springer International Publishing: Cham, Germany, 2020; pp. 151–183. [Google Scholar]
- Noroozi, J.; Pauli, H.; Grabherr, G.; Breckle, S.W. The subnival–nival vascular plant species of Iran: A unique high-mountain flora and its threat from climate warming. Biodivers. Conserv. 2011, 20, 1319–1338. [Google Scholar] [CrossRef]
- Doostmohammadi, M.; Kilian, N. Lactuca pumila (Asteraceae, Cichorieae) revisited-additional evidence for a phytogeographical link between SE Zagros and Hindu Kush. Phytotaxa 2017, 307, 133–140. [Google Scholar] [CrossRef]
- Gibbard, P.L.; Head, M.J.; Walker, M.J.C. The Subcommission on Quaternary Stratigraphy. Formal ratification of the Quaternary System/Period and the Pleistocene Series/Epoch with a base at 2.58 Ma. J. Quat. Sci. 2010, 25, 96–102. [Google Scholar] [CrossRef]
- Sanmartin, I. Dispersal vs. vicariance in the Mediterranean: Historical biogeography of the Palearctic Pachydeminae (Coleoptera, Scarabaeoidea). J. Biogeogr. 2003, 30, 1883–1897. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Collazos, E.; Sanchez-Gómez, P.; Jiménez, F.; Catalán, P. The phylogeographical history of the Iberian steppe plant Ferula loscosii (Apiaceae): A test of the abundant-centre hypothesis. Mol. Ecol. 2009, 18, 848–861. [Google Scholar] [CrossRef]
- Garciá-Aloy, S.; Daniel, V.; Cristina, R.; Isabel, S.; Pablo, V.; Julián Molero, B.; Peris, K.; Juan José, A.; María Luisa, A. North-west Africa as a source and refuge area of plant biodiversity: A case study on Campanula kremeri and Campanula occidentalis. J. Biogeogr. 2017, 44, 2057–2068. [Google Scholar] [CrossRef]
- Gil-López, M.J.; Segarra-Moragues, J.G.; Casimiro-Soriguer, R.; Ojeda, F. From the Strait of Gibraltar to northern Europe: Pleistocene refugia and biogeographic history of heather (Calluna vulgaris, Ericaceae). Bot. J. Linn. Soc. 2022, 198, 41–56. [Google Scholar] [CrossRef]
- Davis, P.H.; Hedge, I.C. Floristic links between NW Africa and SW Asia. Ann. Naturhist. Mus. Wien 1971, 73, 43–57. [Google Scholar]
- Zohary, M. Geobotanical Foundations of the Middle East; Gustav Fischer Verlag: Stuttgart, Germany, 1973; Volumes 1 and 2. [Google Scholar]
- Léonard, J. Contribution à Létude de la Flore et de la Végétation des Déserts d’Iran, Fascicule. 8 (Étude des Aires de Distribution les Phytochories, les Chorotypes) and Fascicule. 9 (Considerapays Phytogeographiques Sur les Phytochories Irano-Touranienne, Saharo-Sindienne et de la Somalie-paysMasai); Jardin Botanique National de Belgique: Meise, Belgium, 1988 and 1989. [Google Scholar]
- Xiang, K.L.; Aytac, Z.; Liu, Y.; Espinosa, F.; Jabbour, F.; Byng, J.W.; Zhang, C.F.; Erst, A.S.; Wang, W. Recircumscription of Delphinium subgen Delphinium (Ranunculaceae) and implications for its biogeography. Taxon 2017, 66, 554–566. [Google Scholar] [CrossRef]
- Rojas-Andrés, B.M.; Padilla-García, N.; de Pedro, M.; López-González, N.; Delgado, L.; Albach, D.C.; Castro, M.; Castro, S.; Loureiro, J.; Martínez-Ortega, M.M. Environmental differences are correlated with the distribution pattern of cytotypes in Veronica subsection Pentasepalae at a broad scale. Ann. Bot. 2020, 125, 471–484. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Taxon Name | Chromosome Number/Ploidy Level | Locality | Herbarium (Voucher) |
|---|---|---|---|---|
| 1 | V. acrotheca | 2n = 16/2x | Iran: Lorestan, Lake Gahar | M. Mirtadzadini and al. (3975 MIR) |
| 2 | V. khorassanica | 2n = 16/2x | Iran: Semnan, Shahrud, Northwest of Mojen Waterfall | M. Mirtadzadini (3976 MIR) |
| 3 | V. kurdica subsp. filicaulis | 2n = 16/2x | Iran: Bakhtiari, Tshoghakhor, Mt. Kallar | M. Mirtadzadini (3563 MIR) |
| 4 | V. kurdica subsp. filicaulis | 2n = 16/2x | Iran: Kerman, Mt. Bahr-Aseman | F. Rezanejad (3511 MIR) |
| 5 | V. kurdica subsp. kurdica | 2n = 16/2x | Iran: Damavand, East of Lake Tar | M. Mirtadzadini (3971 MIR) |
| 6 | V. kurdica subsp. kurdica | 2n = 16/2x | Iran: Ghazvin, West of Abe-garm, Kise-jin to Dashtak | M. Mirtadzadini (3984 MIR) |
| 7 | V. microcarpa | 2n = 16/2x | Iran: Azarbiajan, Jolfa, above St. Stephanus Church | M. Mirtadzadini (3972 MIR) |
| 8 | V. orientalis | 2n = 48/6x | Iran: Azarbaijan, near Takab | M. Doostmohammadi (3593 MIR) |
| 9 | V. rechingeri | 2n = 32/4x | Iran: Mazandaran, Kalardasht, Vandarbon to Tange-Galu | M. Doostmohammadi and A. Ghorbanalizadeh (3634 MIR) |
| 10 | V. schizostegia | 2n = 16/2x | Iran: Kordestan, South of Marivan, Dezli to Nowsud | M. Mirtadzadini (3977 MIR) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doostmohammadi, M.; Bordbar, F.; Albach, D.C.; Mirtadzadini, M. Phylogeny and Historical Biogeography of Veronica Subgenus Pentasepalae (Plantaginaceae): Evidence for Its Origin and Subsequent Dispersal. Biology 2022, 11, 639. https://doi.org/10.3390/biology11050639
Doostmohammadi M, Bordbar F, Albach DC, Mirtadzadini M. Phylogeny and Historical Biogeography of Veronica Subgenus Pentasepalae (Plantaginaceae): Evidence for Its Origin and Subsequent Dispersal. Biology. 2022; 11(5):639. https://doi.org/10.3390/biology11050639
Chicago/Turabian StyleDoostmohammadi, Moslem, Firouzeh Bordbar, Dirk C. Albach, and Mansour Mirtadzadini. 2022. "Phylogeny and Historical Biogeography of Veronica Subgenus Pentasepalae (Plantaginaceae): Evidence for Its Origin and Subsequent Dispersal" Biology 11, no. 5: 639. https://doi.org/10.3390/biology11050639
APA StyleDoostmohammadi, M., Bordbar, F., Albach, D. C., & Mirtadzadini, M. (2022). Phylogeny and Historical Biogeography of Veronica Subgenus Pentasepalae (Plantaginaceae): Evidence for Its Origin and Subsequent Dispersal. Biology, 11(5), 639. https://doi.org/10.3390/biology11050639