
Effect of Bis(maltolato)oxovanadium(IV) on Zinc, Copper, and Manganese Homeostasis and DMT1 mRNA Expression in Streptozotocin-Induced Hyperglycemic Rats

 ,
, 
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
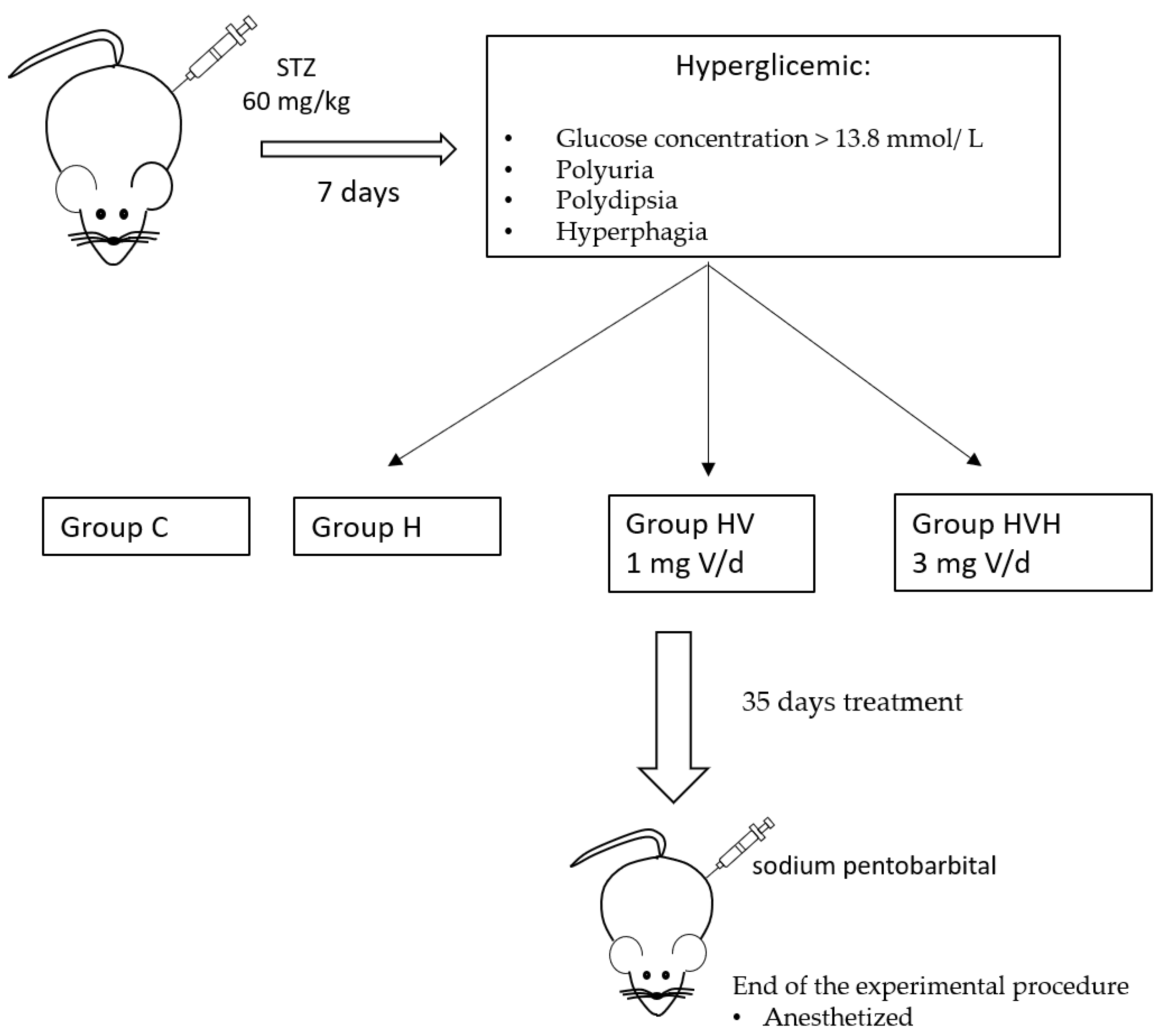
2.1. Animals and Diets
2.2. Determination of Total Metals
2.3. Analysis of DMT1 Gene Expression in Liver Samples by RT-qPCR
2.4. Determination of Antioxidant Enzyme Activity and Proinflammatory Parameters
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, D.T.; Hamilton, K.; Burnand, R.; Smith, C.P.; Tomlinson, D.R.; Riccardi, D. Altered Expression of Iron Transport Proteins in Streptozotocin-Induced Diabetic Rat Kidney. Biochim. Biophys. Acta—Mol. Basis Dis. 2005, 1740, 79–84. [Google Scholar] [CrossRef]
- El-Maraghy, S.A.; Mehana, N.A. Modulatory Effects of L-Arginine and Soy Enriched Diet on Bone Homeostasis Abnormalities in Streptozotocin-Induced Diabetic Rats. Chem. Biol. Interact. 2015, 229, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and Characterization of a Mammalian Proton-Coupled Metal-Ion Transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, S.; Adulcikas, J.; Sohal, S.S.; Myers, S. Zinc Stimulates Glucose Oxidation and Glycemic Control by Modulating the Insulin Signaling Pathway in Human and Mouse Skeletal Muscle Cell Lines. PLoS ONE 2018, 13, e0191727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazi, T.G.; Afridi, H.I.; Kazi, N.; Jamali, M.K.; Arain, M.B.; Jalbani, N.; Kandhro, G.A. Copper, Chromium, Manganese, Iron, Nickel, and Zinc Levels in Biological Samples of Diabetes Mellitus Patients. Biol. Trace Elem. Res. 2008, 122, 1–18. [Google Scholar] [CrossRef]
- Chimienti, F. Zinc, Pancreatic Islet Cell Function and Diabetes: New Insights into an Old Story. Nutr. Res. Rev. 2013, 26, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chabosseau, P.; Rutter, G.A. Zinc and Diabetes. Arch. Biochem. Biophys. 2016, 611, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Haase, H.; Maret, W. Protein Tyrosine Phosphatases as Targets of the Combined Insulinomimetic Effects of Zinc and Oxidants. Biometals 2005, 18, 333–338. [Google Scholar] [CrossRef]
- Lu, J.; Ji, W.; Zhao, M.; Wang, M.; Yan, W.; Chen, M.; Ren, S.; Yuan, B.; Wang, B.; Chen, L. Protamine Zinc Insulin Combined with Sodium Selenite Improves Glycometabolism in the Diabetic KKAy Mice. Sci. Rep. 2016, 6, 26563. [Google Scholar] [CrossRef] [Green Version]
- Kretsinger, R.H.; Uversky, V.N.; Permyakov, E.A. (Eds.) Encyclopedia of Metalloproteins; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-1532-9. [Google Scholar]
- Zhou, L.; Cao, L.; Wang, X.; Wang, M.; Wang, H.; Zhong, Z.; Xu, Z.; Chen, H.; Li, L.; Li, M.; et al. Metal Adaptation Strategies of Deep-Sea Bathymodiolus Mussels from a Cold Seep and Three Hydrothermal Vents in the West Pacific. Sci. Total Environ. 2020, 707, 136046. [Google Scholar] [CrossRef]
- Agil, A.; Chayah, M.; Visiedo, L.; Navarro-Alarcon, M.; Rodríguez Ferrer, J.M.; Tassi, M.; Reiter, R.J.; Fernández-Vázquez, G. Melatonin Improves Mitochondrial Dynamics and Function in the Kidney of Zücker Diabetic Fatty Rats. JCM 2020, 9, 2916. [Google Scholar] [CrossRef] [PubMed]
- Domingo, J.L.; Gómez, M. Vanadium Compounds for the Treatment of Human Diabetes Mellitus: A Scientific Curiosity? A Review of Thirty Years of Research. Food Chem. Toxicol. 2016, 95, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-González, C.; Rivas-García, L.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J.; Aranda, P.; Montes-Bayón, M.; Llopis, J. Vanadium Decreases Hepcidin MRNA Gene Expression in STZ-Induced Diabetic Rats, Improving the Anemic State. Nutrients 2021, 13, 1256. [Google Scholar] [CrossRef] [PubMed]
- Ścibior, A.; Wnuk, E.; Gołębiowska, D. Wild Animals in Studies on Vanadium Bioaccumulation—Potential Animal Models of Environmental Vanadium Contamination: A Comprehensive Overview with a Polish Accent. Sci. Total Environ. 2021, 785, 147205. [Google Scholar] [CrossRef]
- Aureliano, M. Decavanadate Toxicology and Pharmacological Activities: V10 or V1, Both or None? Oxidative Med. Cell. Longev. 2016, 2016, 6103457. [Google Scholar] [CrossRef] [Green Version]
- Treviño, S.; Díaz, A.; Sánchez-Lara, E.; Sanchez-Gaytan, B.L.; Perez-Aguilar, J.M.; González-Vergara, E. Vanadium in Biological Action: Chemical, Pharmacological Aspects, and Metabolic Implications in Diabetes Mellitus. Biol. Trace Elem. Res. 2019, 188, 68–98. [Google Scholar] [CrossRef] [Green Version]
- Ścibior, A.; Pietrzyk, Ł.; Plewa, Z.; Skiba, A. Vanadium: Risks and Possible Benefits in the Light of a Comprehensive Overview of Its Pharmacotoxicological Mechanisms and Multi-Applications with a Summary of Further Research Trends. J. Trace Elem. Med. Biol. 2020, 61, 126508. [Google Scholar] [CrossRef]
- Thompson, K.H.; Lichter, J.; LeBel, C.; Scaife, M.C.; McNeill, J.H.; Orvig, C. Vanadium Treatment of Type 2 Diabetes: A View to the Future. J. Inorg. Biochem. 2009, 103, 554–558. [Google Scholar] [CrossRef]
- Rivas-García, L.; Quiles, J.L.; Varela-López, A.; Arredondo, M.; Lopez, P.; Diéguez, A.R.; Montes-Bayon, M.; Aranda, P.; Llopis, J.; Sánchez-González, C. In Vitro Study of the Protective Effect of Manganese against Vanadium-Mediated Nuclear and Mitochondrial DNA Damage. Food Chem. Toxicol. 2020, 135, 110900. [Google Scholar] [CrossRef]
- Oster, M.H.; Llobet, J.M.; Domingo, J.L.; Bruce German, J.; Keen, C.L. Vanadium Treatment of Diabetic Sprague-Dawley Rats Results in Tissue Vanadium Accumulation and pro-Oxidant Effects. Toxicology 1993, 83, 115–130. [Google Scholar] [CrossRef]
- Ścibior, A.; Adamczyk, A.; Gołębiowska, D.; Niedźwiecka, I. Effect of 12-Week Vanadate and Magnesium Co-Administration on Chosen Haematological Parameters as Well as on Some Indices of Iron and Copper Metabolism and Biomarkers of Oxidative Stress in Rats. Environ. Toxicol. Pharmacol. 2012, 34, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Ścibior, A.; Kurus, J. Vanadium and Oxidative Stress Markers—In Vivo Model: A Review. CMC 2019, 26, 5456–5500. [Google Scholar] [CrossRef] [PubMed]
- Krośniak, M.; Kowalska, J.; Francik, R.; Gryboś, R.; Blusz, M.; Kwiatek, W.M. Influence of Vanadium–Organic Ligands Treatment on Selected Metal Levels in Kidneys of STZ Rats. Biol. Trace Elem. Res. 2013, 153, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krośniak, M.; Francik, R.; Kowalska, J.; Gryboś, R.; Blusz, M.; Kwiatek, W.M. Effects of Vanadium Complexes Supplementation on V, Fe, Cu, Zn, Mn, Ca and K Concentration in STZ Diabetic Rat’s Spleen. Acta Pol. Pharm. 2013, 70, 71–77. [Google Scholar]
- Thompson, K.H.; Tsukada, Y.; Xu, Z.; Battell, M.; McNeill, J.H.; Orvig, C. Influence of Chelation and Oxidation State on Vanadium Bioavailability, and Their Effects on Tissue Concentrations of Zinc, Copper, and Iron. BTER 2002, 86, 31–44. [Google Scholar] [CrossRef]
- Ścibior, A.; Adamczyk, A.; Gołębiowska, D.; Niedźwiecka, I.; Fornal, E. The Influence of Combined Magnesium and Vanadate Administration on the Level of Some Elements in Selected Rat Organs: V–Mg Interactions and the Role of Iron-Essential Protein (DMT-1) in the Mechanism Underlying Altered Tissues Iron Level. Metallomics 2014, 6, 907–920. [Google Scholar] [CrossRef]
- Ścibior, A.; Gołębiowska, D.; Adamczyk, A.; Kurus, J.; Staniszewska, M.; Sadok, I. Evaluation of Lipid Peroxidation and Antioxidant Defense Mechanisms in the Bone of Rats in Conditions of Separate and Combined Administration of Vanadium (V) and Magnesium (Mg). Chem. Biol. Interact. 2018, 284, 112–125. [Google Scholar] [CrossRef]
- Sánchez-González, C.; Rivas-García, L.; López-Chaves, C.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J.; Gómez-Aracena, J.; Vera-Ramírez, L.; Montes-Bayon, M.; Sanz-Medel, A.; et al. Exposure to Bis(Maltolato)Oxovanadium(IV) Increases Levels of Hepcidin MRNA and Impairs the Homeostasis of Iron but Not That of Manganese. Food Chem. Toxicol. 2014, 73, 113–118. [Google Scholar] [CrossRef]
- McNeill, J.H.; Yuen, V.G.; Hoveyda, H.R.; Orvig, C. Bis(Maltolato)Oxovanadium(IV) Is a Potent Insulin Mimic. J. Med. Chem. 1992, 35, 1489–1491. [Google Scholar] [CrossRef]
- Giampieri, F.; Quiles, J.L.; Orantes-Bermejo, F.J.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Sánchez-González, C.; Llopis, J.; Rivas-García, L.; Afrin, S.; Varela-López, A.; et al. Are By-Products from Beeswax Recycling Process a New Promising Source of Bioactive Compounds with Biomedical Properties? Food Chem. Toxicol. 2018, 112, 126–133. [Google Scholar] [CrossRef]
- Sanchez-Gonzalez, C.; Moreno, L.; Lopez-Chaves, C.; Nebot, E.; Pietschmann, P.; Rodriguez-Nogales, A.; Galvez, J.; Montes-Bayon, M.; Sanz-Medel, A.; Llopis, J. Effect of Vanadium on Calcium Homeostasis, Osteopontin MRNA Expression, and Bone Microarchitecture in Diabetic Rats. Metallomics 2017, 9, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gonzalez, C.; Bermudez-Peña, C.; Guerrero-Romero, F.; Trenzado, C.E.; Montes-Bayon, M.; Sanz-Medel, A.; Llopis, J. Effect of Bis(Maltolato)Oxovanadium (IV) (BMOV) on Selenium Nutritional Status in Diabetic Streptozotocin Rats. Br. J. Nutr. 2012, 108, 893–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-González, C.; López-Chaves, C.; Trenzado, C.E.; Aranda, P.; López-Jurado, M.; Gómez-Aracena, J.; Montes-Bayón, M.; Sanz-Medel, A.; Llopis, J. Changes in Iron Metabolism and Oxidative Status in STZ-Induced Diabetic Rats Treated with Bis(Maltolato) Oxovanadium (IV) as an Antidiabetic Agent. Sci. World J. 2014, 2014, 706074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C.; Torres, M.; Bermúdez-Peña, M.C.; Aranda, P.; Montes-Bayón, M.; Sanz-Medel, A.; Llopis, J. Bioavailability, Tissue Distribution and Hypoglycaemic Effect of Vanadium in Magnesium-Deficient Rats. Magnes. Res. 2011, 24, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-Peña, M.C.; López-Chaves, C.; Llopis, J.; Guerrero-Romero, F.; Montes-Bayón, M.; Sanz-Medel, A.; Sánchez-González, C. Aggravation by Vanadium of Magnesium Deficiency in STZ-Induced Diabetic Rats. Magnes. Res. 2013, 26, 74–82. [Google Scholar] [CrossRef]
- Thompson, K.H.; Orvig, C. Vanadium in Diabetes: 100 Years from Phase 0 to Phase I. J. Inorg. Biochem. 2006, 100, 1925–1935. [Google Scholar] [CrossRef]
- Wang, X.; Karvonen-Gutierrez, C.A.; Herman, W.H.; Mukherjee, B.; Harlow, S.D.; Park, S.K. Urinary Metals and Incident Diabetes in Midlife Women: Study of Women’s Health Across the Nation (SWAN). BMJ Open Diab. Res. Care 2020, 8, e001233. [Google Scholar] [CrossRef]
- Liu, B.; Feng, W.; Wang, J.; Li, Y.; Han, X.; Hu, H.; Guo, H.; Zhang, X.; He, M. Association of Urinary Metals Levels with Type 2 Diabetes Risk in Coke Oven Workers. Environ. Pollut. 2016, 210, 1–8. [Google Scholar] [CrossRef]
- Mackenzie, B.; Takanaga, H.; Hubert, N.; Rolfs, A.; Hediger, M.A. Functional Properties of Multiple Isoforms of Human Divalent Metal-Ion Transporter 1 (DMT1). Biochem. J. 2007, 403, 59–69. [Google Scholar] [CrossRef]
- Ghio, A.J.; Stonehuerner, J.; Soukup, J.M.; Dailey, L.A.; Kesic, M.J.; Cohen, M.D. Iron Diminishes the In Vitro Biological Effect of Vanadium. J. Inorg. Biochem. 2015, 147, 126–133. [Google Scholar] [CrossRef]
- Karganov, M.Y.; Alchinova, I.B.; Tinkov, A.A.; Medvedeva, Y.S.; Lebedeva, M.A.; Ajsuvakova, O.P.; Polyakova, M.V.; Skalnaya, M.G.; Burtseva, T.I.; Notova, S.V.; et al. Streptozotocin (STZ)-Induced Diabetes Affects Tissue Trace Element Content in Rats in a Dose-Dependent Manner. Biol. Trace Elem. Res. 2020, 198, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, J.; Ognik, K.; Stępniowska, A.; Zduńczyk, Z.; Kozłowski, K. The Effect of the Source and Dose of Manganese on the Performance, Digestibility and Distribution of Selected Minerals, Redox, and Immune Status of Turkeys. Poult. Sci. 2019, 98, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xiao, J.; Zhang, J.; Chen, H.; Li, D.; Li, L.; Cao, J.; Xie, L.; Luo, Y. Effects of Dietary Cu and Zn on the Accumulation, Oxidative Stress and the Expressions of Immune-Related Genes in the Livers of Nile Tilapia (Oreochromis Niloticus). Fish Shellfish Immunol. 2020, 100, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, C.; Li, Z.; Li, J.; Chen, Y.; Wang, T.; Wang, C. Effects of Zinc Oxide Nanoparticles on Growth, Intestinal Barrier, Oxidative Status and Mineral Deposition in 21-Day-Old Broiler Chicks. Biol. Trace Elem. Res. 2022, 200, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, K.S.; Sivagaami, P.; Ramkumar, A.; Murugadas, A.; Srinivasan, V.; Arun, S.; Senthil Kumar, P.; Thirumurugan, R. Bio-Functionalized Zinc Oxide Nanoparticles: Potential Toxicity Impact on Freshwater Fish Cyprinus Carpio. Chemosphere 2022, 290, 133220. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, J.; Lin, G.; Yu, D.; Xiong, Y.L. Upregulation of Antioxidant Enzymes by Organic Mineral Co-Factors to Improve Oxidative Stability and Quality Attributes of Muscle from Laying Hens. Food Res. Int. 2019, 125, 108575. [Google Scholar] [CrossRef]
- Illán-Cabeza, N.A.; Jiménez-Pulido, S.B.; Hueso-Ureña, F.; Ramírez-Expósito, M.J.; Martínez-Martos, J.M.; Moreno-Carretero, M.N. Relationship between the Antiproliferative Properties of Cu(II) Complexes with the Schiff Base Derived from Pyridine-2-Carboxaldehyde and 5,6-Diamino-1,3-Dimethyluracil and the Redox Status Mediated by Antioxidant Defense Systems on Glioma Tumoral Cells. J. Inorg. Biochem. 2020, 207, 111053. [Google Scholar] [CrossRef]
- Atli, G. How Metals Directly Affect the Antioxidant Status in the Liver and Kidney of Oreochromis Niloticus? An in Vitro Study. J. Trace Elem. Med. Biol. 2020, 62, 126567. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Eweedah, N.M.; Moustafa, E.M.; El-Sharawy, M.E.; Soliman, A.A.; Amer, A.A.; Atia, M.H. Copper Nanoparticles Mitigate the Growth, Immunity, and Oxidation Resistance in Common Carp (Cyprinus Carpio). Biol. Trace Elem. Res. 2020, 198, 283–292. [Google Scholar] [CrossRef]
- Gao, W.; Huang, L.; Zhang, X.; Ma, X.; Wang, W.; Zheng, Y.; Geng, W.; Liu, C.; Wei, S.; Yang, L.; et al. Effect of Maternal Marginal Zinc Deficiency on Development, Redox Status, and Gene Expression Related to Oxidation and Apoptosis in an Avian Embryo Model. Oxidative Med. Cell. Longev. 2021, 2021, 9013280. [Google Scholar] [CrossRef]
- Tok, A.; Özer, A.; Baylan, F.A.; Kurutaş, E.B. Copper/Zinc Ratio Can Be a Marker to Diagnose Ectopic Pregnancy and Is Associated with the Oxidative Stress Status of Ectopic Pregnancy Cases. Biol. Trace Elem. Res. 2021, 199, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Faghfouri, A.H.; Zarezadeh, M.; Aghapour, B.; Izadi, A.; Rostamkhani, H.; Majnouni, A.; Abu-Zaid, A.; Kord Varkaneh, H.; Ghoreishi, Z.; Ostadrahimi, A. Clinical Efficacy of Zinc Supplementation in Improving Antioxidant Defense System: A Comprehensive Systematic Review and Time-Response Meta-Analysis of Controlled Clinical Trials. Eur. J. Pharmacol. 2021, 907, 174243. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.H.; Guimaraes, I.; Menta, P.R.; Fernandes, L.; Paiva, D.; Ribeiro, T.L.; Celestino, M.L.; Netto, A.S.; Ballou, M.A.; Machado, V.S. Effect of Injectable Trace Mineral Supplementation on Peripheral Polymorphonuclear Leukocyte Function, Antioxidant Enzymes, Health, and Performance in Dairy Cows in Semi-Arid Conditions. J. Dairy Sci. 2022, 105, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Låg, M.; Øvrevik, J.; Totlandsdal, A.I.; Lilleaas, E.M.; Thormodsæter, A.; Holme, J.A.; Schwarze, P.E.; Refsnes, M. Air Pollution-Related Metals Induce Differential Cytokine Responses in Bronchial Epithelial Cells. Toxicol. Vitr. 2016, 36, 53–65. [Google Scholar] [CrossRef]
- Li, S.; Zhao, H.; Wang, Y.; Shao, Y.; Li, J.; Liu, J.; Xing, M. The Inflammatory Responses in Cu-Mediated Elemental Imbalance Is Associated with Mitochondrial Fission and Intrinsic Apoptosis in Gallus Gallus Heart. Chemosphere 2017, 189, 489–497. [Google Scholar] [CrossRef]
- Das, D.C.; Jahan, I.; Uddin, M.d.G.; Hossain, M.d.M.; Chowdhury, M.A.Z.; Fardous, Z.; Rahman, M.d.M.; Kabir, A.K.M.H.; Deb, S.R.; Siddique, M.A.B.; et al. Serum CRP, MDA, Vitamin C, and Trace Elements in Bangladeshi Patients with Rheumatoid Arthritis. Biol. Trace Elem. Res. 2021, 199, 76–84. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | C (n = 9) | H (n = 8) | HV (n = 8) | HVH (n = 8) | PANOVA |
|---|---|---|---|---|---|
| Zn | |||||
| Intake (I) (µg/day) | 609 ± 80 | 1342 ± 95 a | 1090 ± 81 a,b | 560 ± 45 b,c | p < 0.001 |
| Fecal excretion (F) (µg/day) | 294± 49 | 606 ± 177 a | 473 ± 107 a | 259 ± 62 b,c | p < 0.001 |
| Urinary excretion (U) (µg/day) | 3.8 ± 1.3 | 40 ± 5 a | 34 ± 2 a,b | 5.6 ± 2.9 b,c | p < 0.001 |
| Absorbed (A); A = I − F (µg/day) | 323 ± 85 | 735 ± 145 a | 615 ± 126 a | 302 ± 57 b,c | p < 0.001 |
| %Absorbed; %A = (A × 100/I) | 52 ± 9 | 55 ± 12 | 56 ± 10 | 54 ± 10 | NS |
| Retained (R); R = [I − (F/U)] (µg/day) | 318 ± 84 | 681 ± 136 a | 586 ± 127 a | 297 ± 59 b,c | p < 0.001 |
| %Retained; %R = (R × 100/I) | 51 ± 9 | 51 ± 11 | 54 ± 10 | 53 ± 10 | NS |
| Cu | |||||
| Intake (I) (µg/day) | 93 ± 12 | 206 ± 14 a | 168 ± 12 a,b | 86 ± 7 b,c | p < 0.001 |
| Fecal excretion (F) (µg/day) | 55± 10 | 109 ± 10 a | 83 ± 20 a,b | 44 ± 9 b,c | p < 0.001 |
| Urinary excretion (U) (µg/day) | 3.4 ± 0.9 | 4.1 ± 1.3 | 2.l ±1. 2 | 8 ± 6 a,c | p < 0.01 |
| Absorbed (A); A = I − F (µg/day) | 39 ± 9 | 98 ± 19 a | 85 ± 21 a | 41 ± 8 b,c | p < 0.001 |
| %Absorbed; %A = (A × 100/I) | 41 ± 9 | 47 ± 7 | 50 ± 11 | 48 ± 9 | NS |
| Retained (R); R = [I − (F/U)] (µg/day) | 35 ± 9 | 93 ± 19 a | 82 ± 21 a | 33 ± 9,b,c | p < 0.001 |
| %Retained; %R = (R × 100/I) | 38 ± 9 | 45 ± 7 | 49 ± 10a | 38 ± 13 | NS |
| Mn | |||||
| Intake (I) (µg/day) | 192 ± 25 | 415 ± 183 a | 290 ± 101 a,b | 179 ± 33 b,c | p < 0.001 |
| Fecal excretion (F) (µg/day) | 124 ± 25 | 248 ± 73 a | 188 ± 47 a | 95 ± 17 b,c | p < 0.001 |
| Urinary excretion (U) (µg/day) | 0.08 ± 0.2 | 0.33 ± 0.20 a | 0.37 ± 0.22 a | 0.13 ± 0.11 b,c | p < 0.001 |
| Absorbed (A); A = I − F (µg/day) | 68 ± 17 | 181 ± 61 a | 152 ± 41 a | 84 ± 19 b,c | p < 0.001 |
| % Absorbed; %A = (A × 100/I) | 35 ± 9 | 42 ± 16 | 44 ± 12 | 46 ± 9 | NS |
| Retained (R); R = [I − (F/U)] (µg/day) | 67 ± 17 | 180 ± 61 a | 151 ± 41 a | 83 ± 19 b,c | p < 0.001 |
| %Retained; %R = (R × 100/I) | 35 ± 9 | 42 ± 16 | 44 ± 12 | 46 ± 9 | NS |
| Control (n = 9) | H (n = 8) | HV (n = 8) | HVH (n = 8) | PANOVA | |
|---|---|---|---|---|---|
| Serum | |||||
| V | 2.4 ± 0.5 | 6.2 ± 1.3 | 385 ± 96 a,b | 766 ± 202 a,b,c | p < 0.001 |
| Zn | 1505 ± 139 | 1729 ± 82 a | 1783 ± 191 a | 1331 ± 162 b,c | p < 0.001 |
| Cu | 851 ± 117 | 1020 ± 190 a | 1027 ± 132 a | 948 ± 95 | p < 0.05 |
| Mn | 1.6 ± 0.4 | 1.9 ± 0.3 | 1.9 ± 0.9 | 1.0 ± 0.3 b,c | p < 0.05 |
| Kidney | |||||
| Zn | 101 ± 12 | 140 ± 17 a | 128 ± 19 a | 79 ± 10 a,b,c | p < 0.001 |
| Cu | 23 ± 6,3 | 159 ± 28 a | 162 ± 20 a | 69 ± 15 a,b,c | p < 0.001 |
| Mn | 3.5 ± 0.48 | 3.6 ± 0.36 | 3.6 ± 0.37 | 3.3 ± 0.30 | NS |
| Liver | |||||
| Zn | 121 ± 18 | 148 ± 19 a | 141 ± 18 | 69 ± 7 a,b,c | p < 0.001 |
| Cu | 15 ± 1.4 | 24 ± 5,6 a | 17 ± 2.1 b | 15 ± 1.7 b | p < 0.001 |
| Mn | 8.2 ± 1.2 | 8.4 ± 1.2 | 9.8 ± 2.2 | 6.1 ± 0.83 b,c | p < 0.01 |
| Skeletal muscle | |||||
| Zn | 36 ± 4.8 | 46 ± 8.4 | 57 ± 24 a | 39 ± 4 | p < 0.01 |
| Cu | 2.8 ± 0.45 | 2.9 ± 0.45 | 2.7 ± 0.19 | 2.7 ± 0.33 | NS |
| Mn | 0.29 ± 0.08 | 0.32 ± 0.10 | 0.26 ± 0.09 | 0.26 ± 0.05 | NS |
| Spleen | |||||
| Zn | 76 ± 3.8 | 80 ± 3.4 | 77 ± 5 | 75 ± 10 | NS |
| Cu | 1.9 ± 0.11 | 2.2 ± 0.11 | 2.2 ± 0.30 | 2.0 ± 0.22 | p < 0.05 |
| Mn | 0.63 ± 0.05 | 0.65 ± 0.04 | 0.61 ± 0.09 | 0.56 ± 0.06 | NS |
| Heart | |||||
| Zn | 67 ± 8.4 | 65 ± 9.0 | 63 ± 3 | 61 ± 5 | NS |
| Cu | 12 ± 1.5 | 12 ± 2.0 | 12 ± 0.82 | 11 ± 1.4 | NS |
| Mn | 1.2 ± 0.14 | 1.1 ± 0.22 | 0.97 ± 0.05 a | 1.1 ± 0.14 | p < 0.05 |
| Femur | |||||
| Zn | 66 ± 2.7 | 82 ± 8.7 a | 72 ± 16 | 69 ± 10 b | p < 0.05 |
| Cu | 0.42 ± 0.04 | 0.52 ± 008 a | 0.45 ± 0.10 | 0.44 ± 0.04 | p < 0.05 |
| Mn | 0.26 ± 0.02 | 0.28 ± 0.04 | 0.24 ± 0.07 | 0.25 ± 0.04 | NS |
| Tissue | Food Intake | |
|---|---|---|
| Serum | V | −0.390 a |
| Zn | 0.712 a | |
| Cu | 0.423 a | |
| Mn | 0.482 a | |
| Kidney | Zn | 0.788 a |
| Cu | 0.876 a | |
| Mn | NS | |
| Liver | Zn | 0.703 a |
| Cu | 0.664 a | |
| Mn | 0.430 a | |
| Spleen | Zn | NS |
| Cu | 0.499 a | |
| Mn | NS | |
| Skeletal muscle | Zn | 0.375 a |
| Cu | NS | |
| Mn | NS | |
| Heart | Zn | NS |
| Cu | NS | |
| Mn | NS | |
| Femur | Zn | 0.510 a |
| Cu | 0.415 a | |
| Mn | NS |
| Liver Antioxidant Enzymes Activity | Liver Zn | Liver Cu | Liver Mn |
|---|---|---|---|
| GPx | 0.798 a | 0.658 a | 0.529 a |
| GX | NS | NS | NS |
| GST | NS | NS | NS |
| SOD | NS | NS | NS |
| CAT | −0.796 a | −0.446 a | −0.497 a |
| NQO1 | 0.536 a | NS | 0.475 a |
| Protein carbonyl groups | NS | NS | NS |
| MDA | −0.666 a | NS | −0.451 a |
| Serum Proinflammatory Cytokines | Serum Zn | Serum Cu | Serum Mn |
|---|---|---|---|
| Leptin | NS | −0.468 a | NS |
| CRP | NS | NS | NS |
| IL β | NS | NS | NS |
| IL-6 | NS | NS | NS |
| TNF α | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-González, C.; Moreno, L.; Aranda, P.; Montes-Bayón, M.; Llopis, J.; Rivas-García, L. Effect of Bis(maltolato)oxovanadium(IV) on Zinc, Copper, and Manganese Homeostasis and DMT1 mRNA Expression in Streptozotocin-Induced Hyperglycemic Rats. Biology 2022, 11, 814. https://doi.org/10.3390/biology11060814
Sánchez-González C, Moreno L, Aranda P, Montes-Bayón M, Llopis J, Rivas-García L. Effect of Bis(maltolato)oxovanadium(IV) on Zinc, Copper, and Manganese Homeostasis and DMT1 mRNA Expression in Streptozotocin-Induced Hyperglycemic Rats. Biology. 2022; 11(6):814. https://doi.org/10.3390/biology11060814
Chicago/Turabian StyleSánchez-González, Cristina, Laura Moreno, Pilar Aranda, María Montes-Bayón, Juan Llopis, and Lorenzo Rivas-García. 2022. "Effect of Bis(maltolato)oxovanadium(IV) on Zinc, Copper, and Manganese Homeostasis and DMT1 mRNA Expression in Streptozotocin-Induced Hyperglycemic Rats" Biology 11, no. 6: 814. https://doi.org/10.3390/biology11060814
APA StyleSánchez-González, C., Moreno, L., Aranda, P., Montes-Bayón, M., Llopis, J., & Rivas-García, L. (2022). Effect of Bis(maltolato)oxovanadium(IV) on Zinc, Copper, and Manganese Homeostasis and DMT1 mRNA Expression in Streptozotocin-Induced Hyperglycemic Rats. Biology, 11(6), 814. https://doi.org/10.3390/biology11060814