
Anguillid Eels as a Model Species for Understanding Endocrinological Influences on the Onset of Spawning Migration of Fishes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
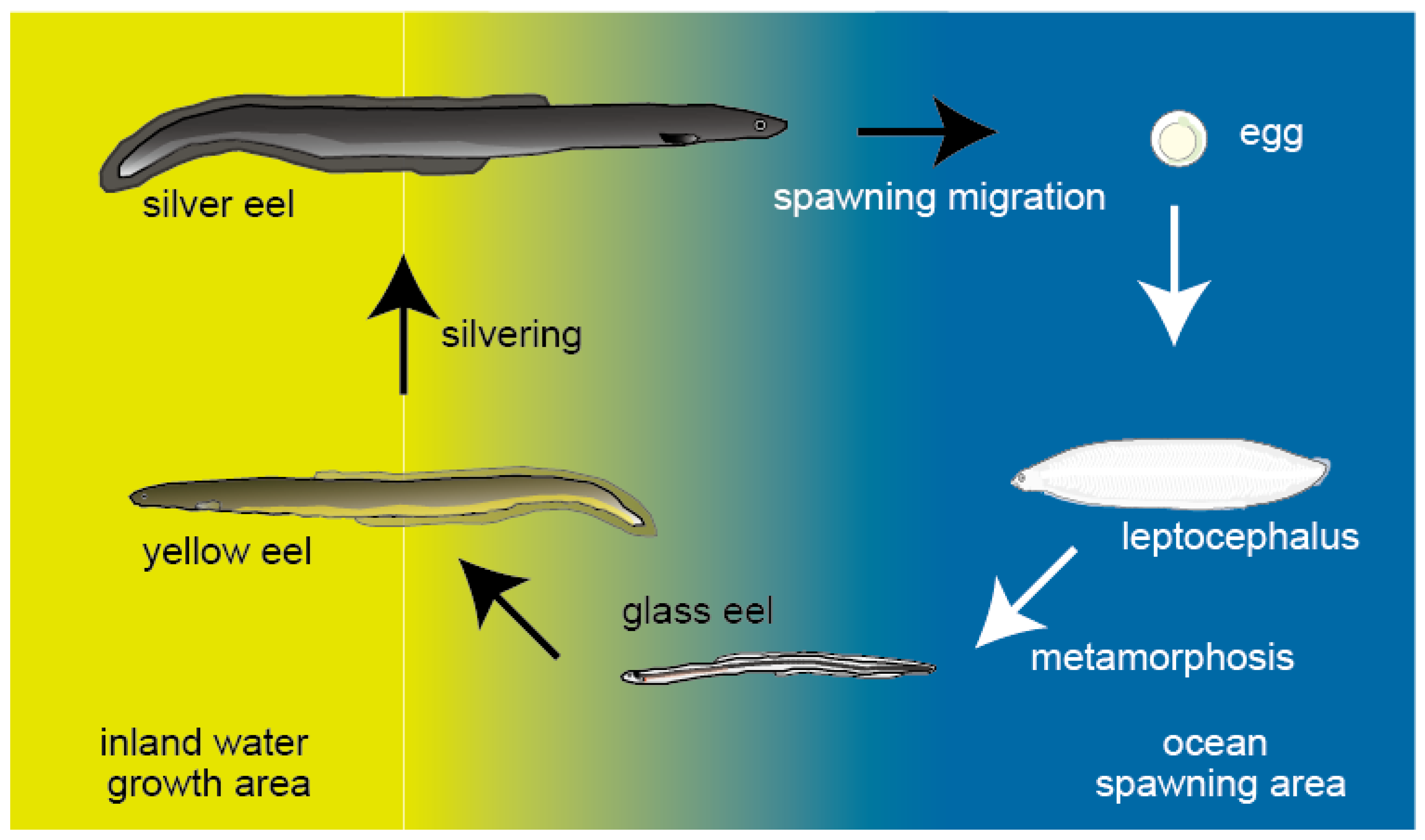
2. Silvering
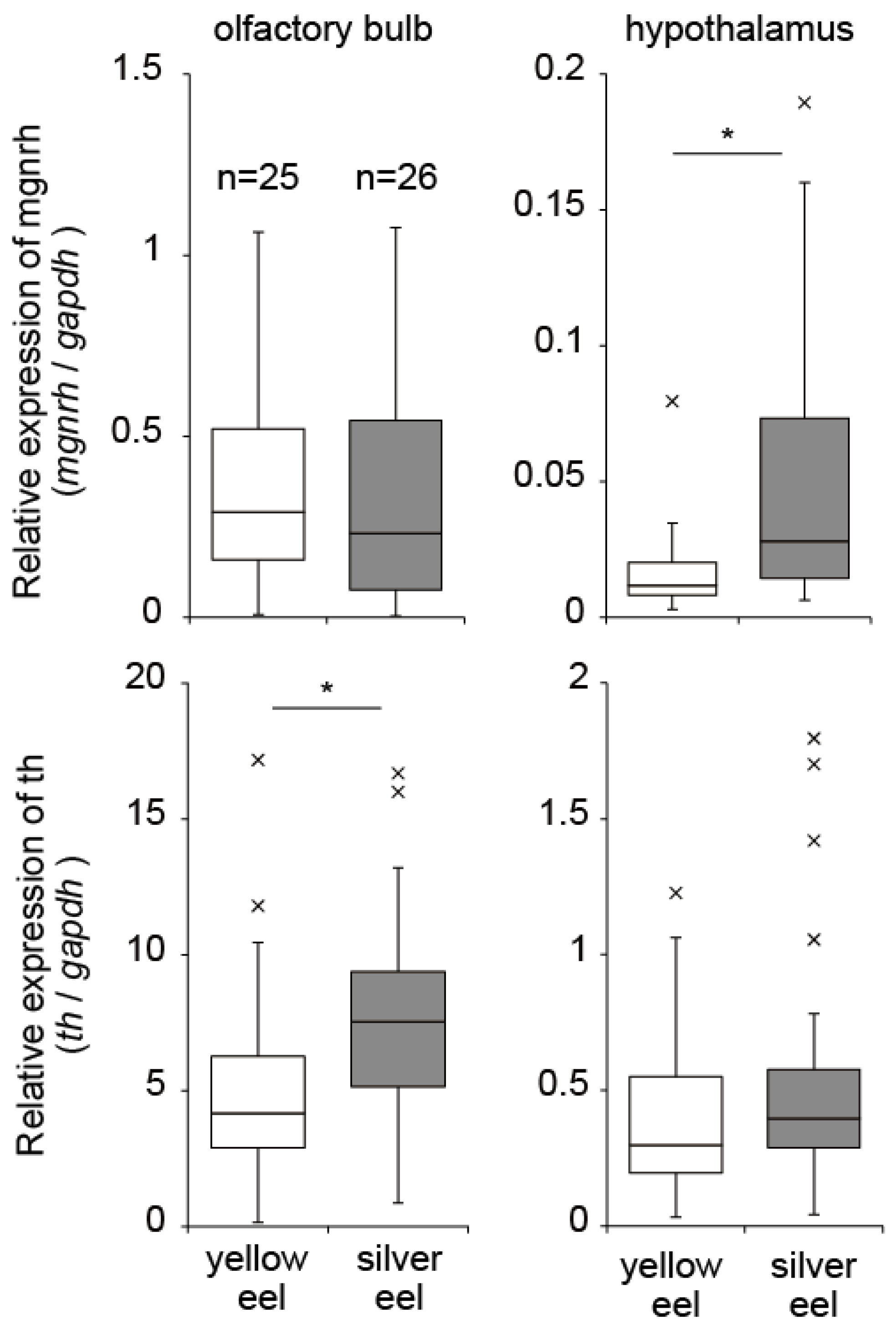
3. Neurohormones
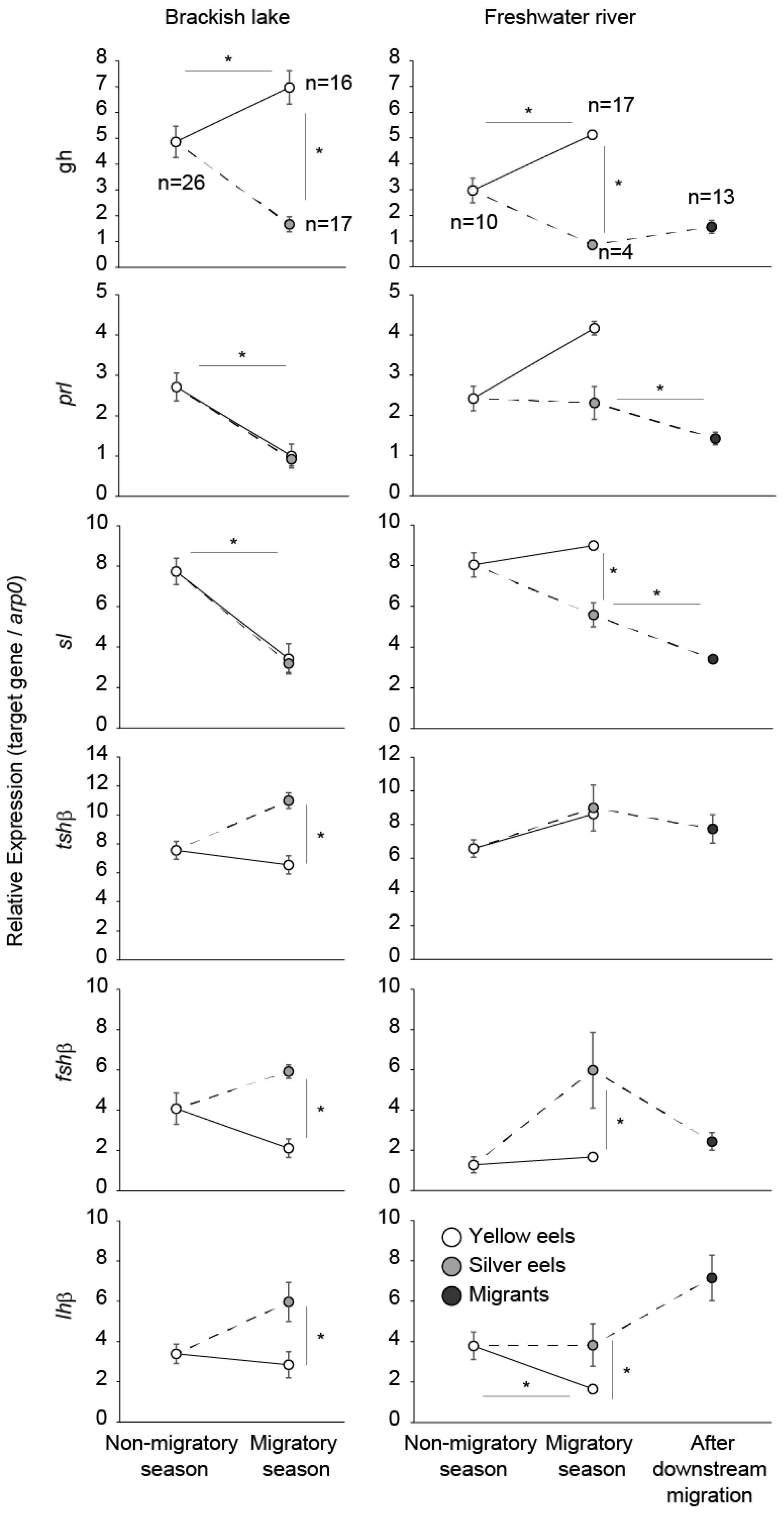
4. Growth Hormone, Prolactin, Somatolactin
5. Thyroid Hormones and Thyroid-Stimulating Hormone
6. Gonadotropins
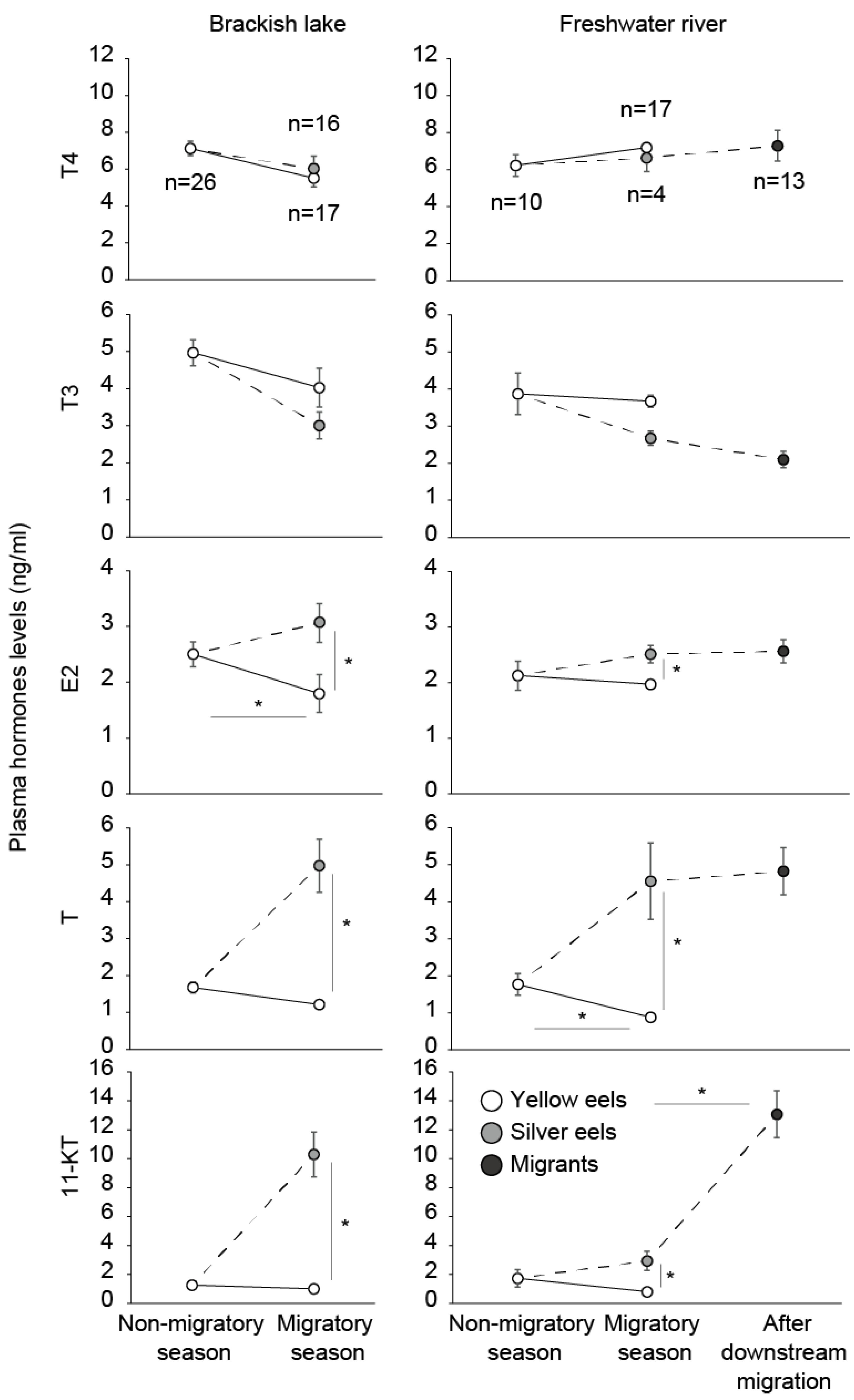
7. Sex Steroids
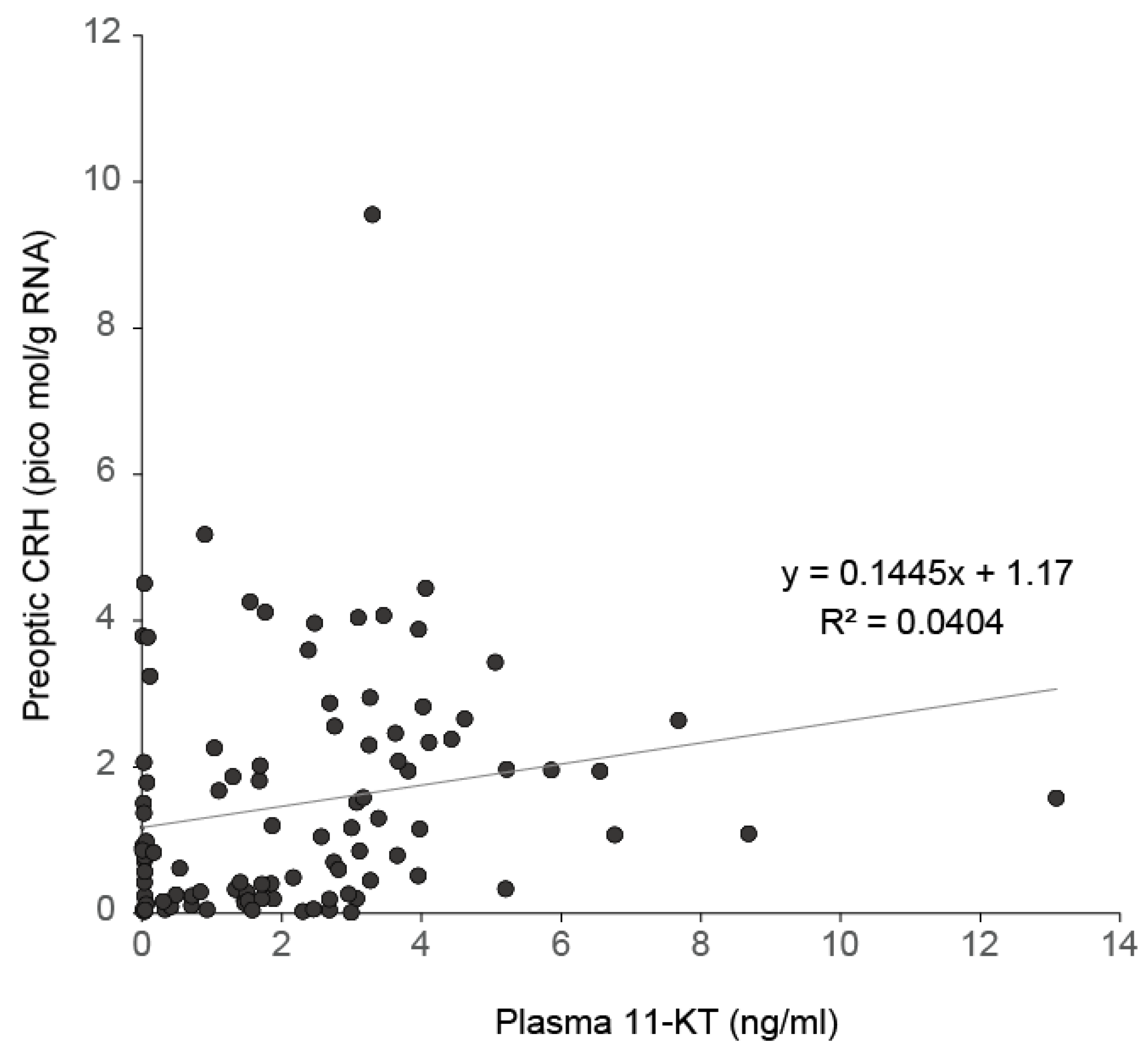
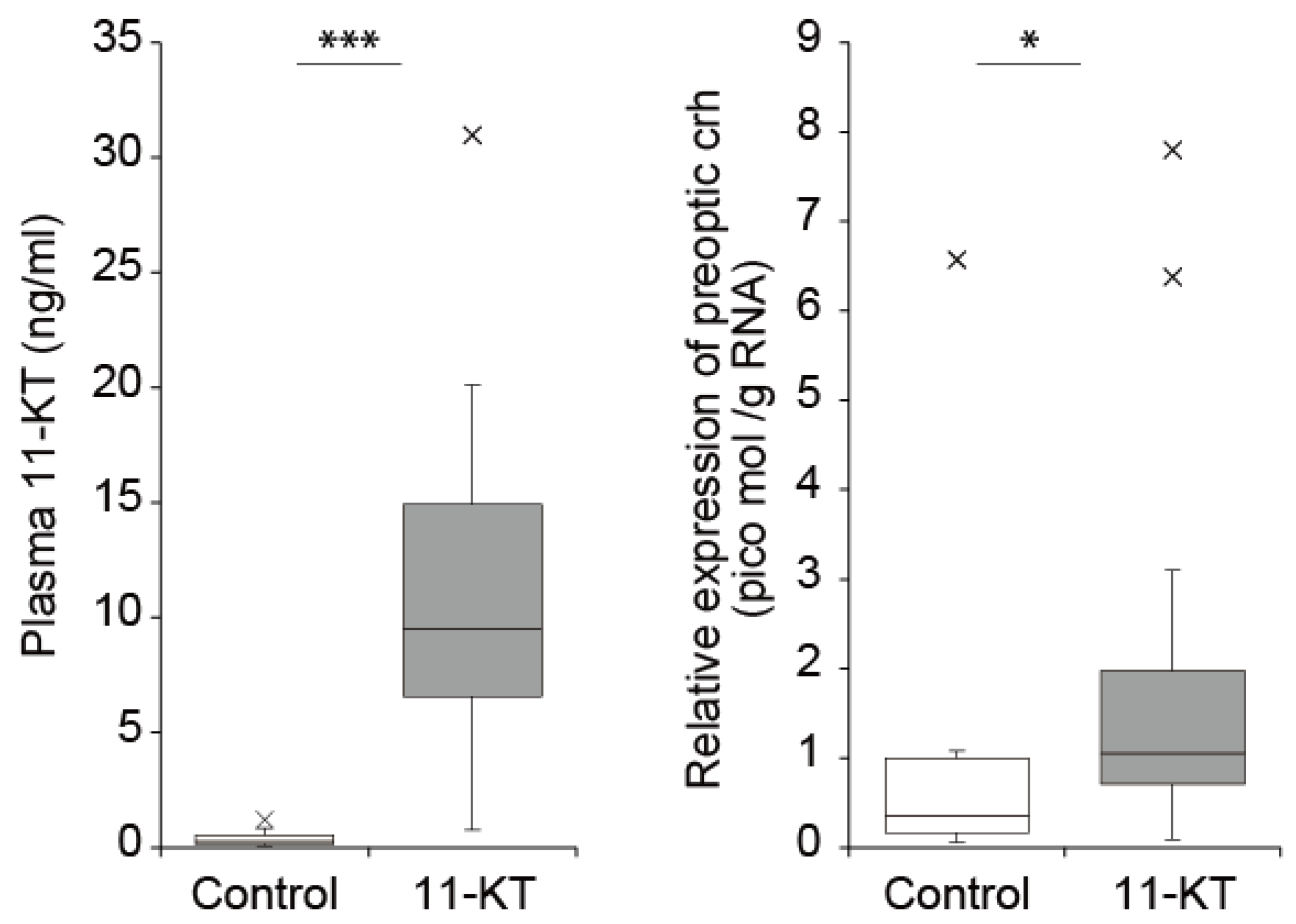
8. Role of Androgen in the Onset of Spawning Migration
9. Interaction of Hormones
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsukamoto, K.; Miller, M.J.; Kotake, A.; Aoyama, J.; Uchida, K. The origin of fish migration: The random escapement hypothesis. In Challenges for Diadromous Fishes in a Dynamic Global Environment. American Fisheries Society Symposium 69; Haro, A., Smith, K.L., Rulifson, R.A., Moffitt, C.M., Klauda, R.J., Dadswell, M.J., Cunjak, R.A., Cooper, J.E., Beal, K.L., Avery, T.S., Eds.; The American Fisheries Society: Bethesda, MD, USA, 2009; pp. 45–61. [Google Scholar]
- Dickhoff, W.W.; Beckman, B.R.; Larsen, D.A.; Duan, C.; Moriyama, S. The role of growth in endocrine regulation of salmon smoltification. Fish Physiol. Biochem. 1997, 17, 231–236. [Google Scholar] [CrossRef]
- Nagae, M.; Fuda, H.; Hara, A.; Saneyoshi, M.; Yamauchi, K. Changes in serum concentrations of immunoglobulin M (IgM), cortisol and thyroxine (T4) during smoltification in the masu salmon Oncorhynchus masou. Fish. Sci. 1994, 60, 241–242. [Google Scholar] [CrossRef] [Green Version]
- Prunet, P.; Boeuf, G.; Bolton, J.P.; Young, G. Smoltification and seawater adaptation in Atlantic salmon (Salmo salar): Plasma prolactin, growth hormone, and thyroid hormones. Gen. Comp. Endocrinol. 1989, 74, 355–364. [Google Scholar] [CrossRef]
- Young, G.; Björnsson, B.T.H.; Prunet, P.; Lin, R.J.; Bern, H.A. Smoltification and seawater adaptation in coho salmon (Oncorhynchus kisutch): Plasma prolactin, growth hormone, thyroid hormones, and cortisol. Gen. Comp. Endocrinol. 1989, 74, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Grau, E.G.; Dickhoff, W.W.; Nishioka, R.S.; Bern, H.A.; Folmar, L.C. Lunar phasing of the thyroxine surge preparatory to seaward migration of salmonid. Fish. Sci. 1981, 211, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Ban, M.; Kasahara, N.; Izumi, T.; Kojima, H.; Harako, T. Physiological and behavioral changes occurring during smoltification in the masu salmon Oncorhynchus masou. Aquaculture 1985, 45, 227–235. [Google Scholar] [CrossRef]
- Ikuta, K. Effects of steroid hormones on migration of salmonid fishes. Bull. Natl. Inst. Aquacult. Suppl. 1994, 2, 23–27. [Google Scholar]
- Tsukamoto, K. Migratory mechanisms and behavioral characateristics in ayu. In Ichthyology Currents; Ueno, T., Okiyama, M., Eds.; Asakurashoten: Tokyo, Japan, 1988; pp. 100–133. [Google Scholar]
- Van Ginneken, V.; Durif, C.; Balm, S.P.; Boot, R.; Verstegen, M.W.A.; Antonissen, E.; van den Thillart, G. Silvering of European eel (Anguilla anguilla L.): Seasonal changes of morphological and metabolic parameters. Anim. Biol. 2007, 57, 63–77. [Google Scholar]
- Okamura, A.; Yamada, Y.; Yokouchi, K.; Horie, N.; Mikawa, N.; Utoh, T.; Tanaka, S.; Tsukamoto, K. A silvering index for the Japanese eel Anguilla japonica. Environ. Biol. Fishes 2007, 80, 77–89. [Google Scholar] [CrossRef]
- Durif, C.; Dufour, S.; Elie, P. The silvering process of Anguilla anguilla: A new classification from the yellow resident to the silver migrating stage. J. Fish Biol. 2005, 66, 1025–1043. [Google Scholar] [CrossRef]
- Han, Y.S.; Liao, I.C.; Huang, Y.S.; He, J.T.; Chang, C.W.; Tzeng, W.N. Synchronous changes of morphology and gonadal development of silvering Japanese eel Anguilla japonica. Aquaculture 2003, 219, 783–796. [Google Scholar] [CrossRef]
- Pankhurst, N.W. Relation of Visual Changes to the onset of sexual maturation in the European eel Anguilla anguilla (L). J. Fish Biol. 1982, 21, 127–140. [Google Scholar] [CrossRef]
- Kleckner, R.C. Swim bladder wall guanine enhancement related to migratory depth in silver phase Anguilla rostrata. Comp. Biochem. Physiol. A 1980, 65, 351–354. [Google Scholar] [CrossRef]
- Kleckner, R.C. Swim bladder volume maintenance related to initial oceanic migratory depth in silver-phase Anguilla rostrata. Science 1980, 208, 1481–1482. [Google Scholar] [CrossRef]
- Yamada, Y.; Zhang, H.; Okamura, A.; Tanaka, S.; Horie, N.; Mikawa, N.; Utoh, T.; Oka, H.P. Morphological and histological changes in the swim bladder during maturation of the Japanese eel. J. Fish Biol. 2001, 58, 804–814. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Sorensen, P.W. Degeneration of the alimentary tract in sexually maturing European eel Anguilla anguilla (L.) and American eel Anguilla rostrata (LeSeuer). Can. J. Zool. 1984, 62, 1143–1148. [Google Scholar] [CrossRef]
- Todd, P.R. Timing of periodicity of migrating New Zealand freshwater eels (Anguilla spp.). N. Z. J. Mar. Freshw. Res. 1981, 15, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Chow, S.; Kurogi, H.; Katayama, S.; Ambe, D.; Okazaki, M.; Watanabe, T.; Ichikawa, T.; Kodama, M.; Aoyama, J.; Shinoda, A.; et al. Japanese eel Anguilla japonica do not assimilate nutrition during the oceanic spawning migration: Evidence from stable isotope analysis. Mar. Ecol. Prog. Ser. 2010, 402, 233–238. [Google Scholar] [CrossRef]
- Lokman, P.M.; Wass, R.T.; Suster, H.C.; Scott, S.G.; Judge, K.F.; Young, G. Changes in steroid hormone profiles and ovarian histology during salmon pituitary-induced vitellogenesis and ovulation in female New Zealand longfinned eels, Anguilla diffenbachii Gray. J. Exp. Biol. 2001, 289, 119–129. [Google Scholar] [CrossRef]
- Svedäng, H.; Wickström, H. Low fat contents in female silver eels: Indications of insufficient energetic stores for migration and gonadal development. J. Fish Biol. 1997, 50, 475–486. [Google Scholar] [CrossRef]
- Ueda, H. Physiology of imprinting and homing migration of Pacific salon. In Physiology and Ecology of Fish Migration; Ueda, H., Tsukamoto, K., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–27. [Google Scholar]
- Wendelaar, B.S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Gorissen, M.; Flik, G. The endocrinology of the stress response in fish: An adaptation-physiological view. In Biology of Stress in Fish; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Amsterdam, The Netherlands, 2016; pp. 75–111. [Google Scholar]
- Clements, S.; Schreck, C.B.; Larsen, D.A.; Dickhoff, W.W. Central administration of corticotropin-releasing hormone stimulates locomotor activity in juvenile chinook salmon (Oncorhynchus tshawytscha). Gen. Comp. Endocrinol. 2002, 125, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Clements, S.; Schreck, C.B. Central administration of corticotropin-releasing hormone alters downstream movement in an artificial stream in juvenile chinook salmon (Oncorhynchus tshawytscha). Gen. Comp. Endocrinol. 2004, 137, 1–8. [Google Scholar] [CrossRef]
- Backström, T.; Pettersson, A.; Johansson, V.; Winberg, S. CRF and urotensin I effects on aggression and anxiety-like behavior in rainbow trout. J. Exp. Biol. 2011, 214, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balment, R.J.; Lu, W.; Weybourne, E.; Warne, J.M. Arginine vasotocin a key hormone in fish physiology and behavior: A review with insights from mammalian models. Gen. Com. Endocrinol. 2006, 147, 9–16. [Google Scholar] [CrossRef]
- Iwata, E.; Nagai, Y.; Sasaki, H. Social rank modulates brain arginine vasotocin immunoreactivity in false clown anemonefish (Amphiprion ocellaris). Fish Physiol. Biochem. 2010, 36, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.; Gozdowska, M.; EwaKulczykowska, E.; Oliveira, R.F. Brain levels of arginine–vasotocin and isotocin in dominant and subordinate males of a cichlid fish. Horm. Behav. 2012, 61, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramallo, R.M.; Grober, M.; Cánepa, M.M.; Morandini, L.; Pandolfi, M. Arginine-vasotocin expression and participation in reproduction and social behavior in males of the cichlid fish Cichlasoma dimerus. Gen. Comp. Endocrinol. 2012, 179, 221–231. [Google Scholar] [CrossRef]
- Yaeger, C.; Ros, A.M.; Cross, V.; DeAngelis, R.A.; Stobaugh, D.J.; Rhodes, J.S. Blockade of arginine vasotocin signaling reduces aggressive behavior and c-Fos expression in the preoptic area and periventricular nucleus of the posterior tuberculum in male Amphiprion ocellaris. Neuroscience 2014, 267, 205–218. [Google Scholar] [CrossRef]
- Berton, O.; Nestler, E.J. New approaches to antidepressant drug discovery: Beyond monoamines. Nat. Rev. Neurosci. 2006, 7, 137–151. [Google Scholar] [CrossRef]
- Narvaes, R.; Martins de Almeida, R.M. Aggressive behavior and three neurotransmitters: Dopamine, GABA, and serotonin—A review of the last 10 years. Psychol. Neurosci. 2014, 7, 601–607. [Google Scholar] [CrossRef]
- Øverli, Ø.; Winberg, S.; Pottinger, T.G. Behavioral and neuroendocrine correlates of Selection for stress responsiveness in rainbow trout—A review. Integr. Comp. Biol. 2005, 45, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paula, J.R.; Messias, J.P.; Grutter, A.S.; Bshary, R.; Soares, M.C. The role of serotonin in the modulation of cooperative behavior. Behav. Ecol. 2015, 26, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Messias, J.P.M.; Paula, J.R.; Grutter, A.S.; Bshary, R.; Soares, M.C. Dopamine disruption increases negotiation for cooperative interactions in a fish. Sci. Rep. 2016, 6, 20817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winberg, S.; Thörnqvist, P.-O. Role of brain serotonin in modulating fish behavior. Curr. Zool. 2016, 62, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Montero, M.; Vidal, B.; King, J.A.; Tramu, G.; Vamdesande, F.; Dufour, S.; Kah, O. Immunocytochemical localization of mammalian GnRH (gonadotropin-releasing hormone) and chicken GnRH-II in the brain of the European silver eel (Anguilla anguilla L.). J. Chem. Neuroanat. 1994, 7, 227–241. [Google Scholar] [CrossRef]
- Okubo, K.; Suetake, H.; Aida, K. Expression of two gonadotropin-releasing hormone (GnRH) precursor genes in various tissues of the Japanese eel and evolution of GnRH. Zool. Sci. 1999, 16, 471–478. [Google Scholar] [CrossRef]
- Sudo, R.; Suetake, H.; Suzuki, Y.; Utoh, T.; Tanaka, S.; Aoyama, J.; Tsukamoto, K. Dynamics of reproduction hormones during downstream migration in females of the Japanese eels, Anguilla japonica. Zool. Sci. 2011, 28, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Rand-Weaver, M.; Kawauchi, H. Growth hormone, prolactin and somatolactin: A structural overview. In The Biochemistry and Molecular Biology of Fishes; Hochachka, P.W., Mommsen, T.P., Eds.; Elsevier Press: Amsterdam, The Netherlands, 1993; pp. 39–59. [Google Scholar]
- Bjornsson, B.T. The biology of salmon growth hormone: From daylight to dominance. Fish Physiol. Biochem. 1997, 17, 9–24. [Google Scholar] [CrossRef]
- Perez-Sanchez, J. The involvement of growth hormone in growth regulation, energy homeostasis and immune function in the gilthead sea bream (Sparus aurata): A short review. Fish Physiol. Biochem. 2000, 22, 135–144. [Google Scholar] [CrossRef]
- Picha, M.M.; Turano, M.J.; Beckman, B.R.; Borski, R.J. Endocrine biomarkers of growth and applications to aquaculture: A minireview of growth hormone, insulin-like growth factor (IGF)-I, and IGF-binding proteins as potential growth indicators in fish. N. Am. J. Aquac. 2008, 70, 196–211. [Google Scholar] [CrossRef]
- Sakamoto, T.; McCormick, S.D. Prolactin and growth hormone in fish osmoregulation. Gen. Comp. Endocrinol. 2006, 147, 24–30. [Google Scholar] [CrossRef]
- Hirano, T. Osmoregulatory roles of prolactin and growth hormone in teleosts. Zool. Sci. 1987, 4, 1133. [Google Scholar]
- Kawauchi, H.; Sower, S.A. The dawn and evolution of hormones in the adenohypophysis. Gen. Comp. Endocrinol. 2006, 148, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Planas, J.V.; Swanson, P.; Rand-Weaver, M.; Dickhoff, W.W. Somatolactin stimulates in vitro gonadal steroidogenesis in coho salmon, Oncorhynchus kisutch. Gen. Comp. Endocrinol. 1992, 87, 1–5. [Google Scholar] [CrossRef]
- Rand-Weaver, M.; Swanson, P.; Kawauchi, H.; Dickhoff, W.W. Somatolactin, a novel pituitary protein - purification and plasma levels during reproductive maturation of coho salmon. J. Endocrinol. 1992, 133, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Le Gac, F.; Blaise, O.; Fostier, A.; Le Bail, P.Y.; Loir, M.; Mourot, B.; Weil, C. Growth hormone (GH) and reproduction: A review. Fish Physiol. Biochem. 1993, 11, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Cavaco, J.E.B.; Santos, C.R.A.; Ingleton, P.M.; Canario, A.V.M.; Power, D.M. Quantification of prolactin (PRL) and PRL receptor messenger RNA in gilthead seabream (Sparus aurata) after treatment with estradiol-17b. Biol. Reprod. 2003, 68, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T. Cell biology of somatolactin. Int. Rev. Cytol. 1996, 169, 1–24. [Google Scholar]
- Kakizawa, S.; Kaneko, T.; Ogasawara, T.; Hirano, T. Changes in plasma somatolactin levels during spawning migration of chum salmon (Oncorhynchus keta). Fish Physiol. Biochem. 1995, 14, 93–101. [Google Scholar] [CrossRef]
- Power, D.M. Developmental ontogeny of prolactin and its receptor in fish. Gen. Comp. Endocrinol. 2005, 142, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Munakata, A.; Amano, M.; Ikuta, K.; Kitamura, S.; Aida, K. Effects of growth hormone and cortisol on the downstream migratory behavior in masu salmon, Oncorhynchus masou. Comp. Biochem. Physiol. A 2013, 164, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Sudo, R.; Suetake, H.; Suzuki, U.; Aoyama, J.; Tsukamoto, K. Profiles of mRNA expression for prolactin, growth hormone, and somatolactin in Japanese eel, Anguilla japonica: The effect of salinity, silvering and seasonal change. Comp. Biochem. Physiol. A 2013, 164, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Aroua, S.; Schmitz, M.; Baloche, S.; Vidal, B.; Rousseau, K.; Dufour, S. Endocrine evidence that silvering, a secondary metamorphosis in the eel, is a pubertal rather than a metamorphic event. Neuroendocrinology 2005, 82, 221–232. [Google Scholar] [CrossRef]
- Van Ginneken, V.; Durif, C.; Dufour, S.; Sbaihi, M.; Boot, R.; Noorlander, K.; Doornbos, J.; Murk, A.J.; van den Thillart, G. Endocrine profiles during silvering of the European eel (Anguilla anguilla L.) living in saltwater. Anim. Biol. 2007, 57, 453–465. [Google Scholar]
- Björnsson, B.T.; Hemre, G.I.; Björnevik, M.; Hansen, T. Photoperiod regulation of plasma growth hormone levels during induced smoltification of underyearling Atlantic salmon. Gen. Comp. Endocrinol. 2000, 119, 17–25. [Google Scholar] [CrossRef]
- Iwata, M.; Yamauchi, K.; Nishioka, R.S.; Lin, R.; Bern, H.A. EVects of thyroxine, growth hormone and cortisol on salinity preference of juvenile coho salmon (Oncorhynchus kisutch). Mar. Behav. Physiol. 1990, 17, 191–201. [Google Scholar] [CrossRef]
- Boeuf, G.; Marc, A.M.; Prunet, P.; Bail, P.Y.L.; Smal, J. Stimulation of parr-smolt transformation by hormonal treatment in Atlantic salmon (Salmo salar L.). Aquaculture 1994, 121, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Johnsson, J.I.; Petersson, E.; Jonsson, E.; Bjornsson, B.T.; Jarvi, T. Domestication and growth hormone alter antipredator behaviour and growth patterns in juvenile brown trout, Salmo trutta. Can. J. Fish Aquat. Sci. 1996, 53, 1546–1554. [Google Scholar] [CrossRef]
- Rankin, M.A. Endocrine effects on migration. Am. Zool. 1991, 31, 217–230. [Google Scholar] [CrossRef]
- Yada, T.; Iguchi, K.; Yamamoto, S.; Sakano, H.; Takasawa, H.; Katsura, K.; Abe, N.; Aawata, S.; Uchida, K. Prolactin and upstream migration of the amphidromous teleost, Ayu Plecoglossus altivelis. Zool. Sci. 2014, 31, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Yu, J.Y.L.; Liao, I.C.; Tzeng, W.N. Salinity preference of silvering Japanese eel Anguilla japonica: Evidence from pituitary prolactin mRNA levels and otolith Sr:Ca ratios. Mar. Ecol. Prog. Ser. 2003, 259, 253–261. [Google Scholar] [CrossRef]
- Kozaka, Y.; Fujii, Y.; Ando, M. Central effects of various ligands on drinking behavior in eels acclimated to seawater. J. Exp. Biol. 2003, 206, 687–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onuma, T.A.; Ban, M.; Makino, K.; Katsumata, H.; Hu, W.W.; Ando, H.; Fukuwaka, M.; Azumaya, T.; Urano, A. Changes in gene expression for GH/PCR/SL family hormones in the pituitary of homing chum salmon during ocean migration through upstream migration. Gen. Comp. Endocrinol. 2010, 166, 537–548. [Google Scholar] [CrossRef]
- Tata, J.R. Amphibian metamorphosis as a model for studying the developmental actions of thyroid hormone. Cell Res. 1998, 8, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Inui, Y.; Miwa, S. Thyroid hormone induces metamorphosis of flounder larvae. Gen. Comp. Endocrinol. 1985, 60, 450–454. [Google Scholar] [CrossRef]
- Hoar, W.S. The physiology of smolting salmonids. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA, 1988; Volume 2, pp. 275–343. [Google Scholar]
- Tsukamoto, K.; Aida, K.; Otake, T. Plasma thyroxine concentration and upstream migratory behavior of juvenile Ayu. Nippon. Suisan Gakkaishi 1988, 54, 1687–1693. [Google Scholar] [CrossRef] [Green Version]
- Castonguay, M.; Dutil, J.D.; Audet, C.; Miller, R. Locomotor activity and concentration of thyroid hormones in migratory and sedentary juvenile American eels. Trans. Am. Fish Soc. 1990, 119, 946–956. [Google Scholar] [CrossRef]
- Edeline, E.; Bardonnet, A.; Bolliet, V.; Dufour, S.; Elie, P. Endocrine control of Anguilla anguilla glass eel dispersal: Effect of thyroid hormones on locomotor activity and rheotactic behavior. Horm. Behav. 2005, 48, 53–63. [Google Scholar] [CrossRef]
- Marchelidon, J.; Le Belle, N.; Hardy, A.; Vidal, B.; Sbaihi, M.; Buruzawa-Gerard, E.; Schmitz, M.; Dufour, S. Etude des variations de parametres anatomiques et endocriniens chez 1′anguille europeenne (Anguilla anguilla) femelle, sedentaire et d’avalaison: Application a la caracterisation du stade argente. Bull. Fr. Peche Piscic. 1999, 355, 349–368. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Liao, I.C.; Tzeng, W.N.; Yu, J.Y.L. Cloning of the cDNA for thyroid stimulating hormone b subunit and changes in activity of the pituitary-thyroid axis during silvering of the Japanese eel, Anguilla japonica. J. Mol. Endocrinol. 2004, 32, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Sudo, R.; Tosaka, R.; Ijiri, S.; Adachi, S.; Aoyama, J.; Tsukamoto, K. 11-ketotestosterone synchronously induces oocyte development and silvering related changes in the Japanese eel, Anguilla japonica. Zool. Sci. 2012, 29, 254–259. [Google Scholar] [CrossRef]
- Gharib, S.D.; Wierman, M.E.; Shupnik, M.A.; Chin, W.W. Molecular biology of the pituitary gonadotropins. Endocr. Rev. 1990, 11, 177–199. [Google Scholar] [CrossRef]
- Kazeto, Y.; Kohara, M.; Miura, T.; Miura, C.; Yamaguchi, S.; Trant, J.M.; Adachi, S.; Yamauchi, K. Japanese eel follicle-stimulating hormone (Fsh) and luteinizing hormone (Lh): Production of biologically active recombinant Fsh and Lh by Drosophila S2 cells and their differential actions on the reproductive biology. Biol. Reprod. 2008, 79, 938–946. [Google Scholar] [CrossRef] [Green Version]
- Kazeto, Y.; Suzuki, H.; Ozaki, Y.; Gen, K. C-terminal peptide (hCTP) of human chorionic gonadotropin enhances in vivo biological activity of recombinant Japanese eel follicle-stimulating hormone and luteinizing hormone produced in FreeStyle 293-F cell lines. Gen. Comp. Endocrinol. 2021, 306, 113731. [Google Scholar] [CrossRef]
- Munakata, A. Migratory behaviors in masu salmon (Oncorhynchus masou) and the influence of endocrinological factors. Aqua-BioSci. Monogr. 2012, 5, 29–65. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Liao, I.C.; Huang, Y.S.; Tzeng, W.N.; Yu, J.Y.L. Profiles of PGH-alpha, GTH-beta, and GTH II-beta mRNA transcript levels at different ovarian stages in the wild female Japanese eel Anguilla japonica. Gen. Comp. Endocrinol. 2003, 133, 8–16. [Google Scholar] [CrossRef]
- Hagihara, S.; Aoyama, J.; Sudo, R.; Limbong, D.; Ijiri, S.; Adachi, S.; Tsukamoto, K. Reproductive physiological characteristics of tropical Celebes eels Anguilla celebesensis in relation to downstream migration and ovarian development. J. Fish Biol. 2020, 96, 558–569. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Chia, J.H.Z.; Kazeto, Y.; Lokman, P.M. Expression of gonadotropin subunit and gonadotropin receptor genes in wild female New Zealand shortfinned eel (Anguilla australis) during yellow and silver stages. Gen. Comp. Endocrinol. 2019, 272, 83–92. [Google Scholar] [CrossRef]
- Sparks, R.T.; Ron, B.; Shepherd, B.S.; Shimoda, S.K.; Iwama, G.K.; Grau, E.G. Effects of environmental salinity and 17alpha-methyltestosterone on growth and oxygen consumption in the tilapia, Oreochromis mossambicus. Comp. Biochem. Physiol. B 2003, 136, 657–665. [Google Scholar] [CrossRef]
- Dasmahapatra, A.K.; Medda, A.K. Effect of estradiol dipropionate and testosterone propionate on the glycogen, lipid, and water contents of liver, muscle, and gonad of male and female (vitellogenic and non-vitellogenic) singi fish (Heteropneustes fossilis Bloch). Gen. Comp. Endocrinol. 1982, 48, 476–484. [Google Scholar] [CrossRef]
- Mommsen, T.P.; Walsh, P.J. Vitellogenesis and oocyte assembly. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA, 1988; pp. 347–406. [Google Scholar]
- Sangiao-Alvarellos, S.; Polakof, S.; Arjona, F.J.; García-Lopez, A.; Martín del Rio, M.P.; Martínez-Rodriguez, G.; Míguez, J.M.; Mancera, J.M.; Soengas, J.L. Influence of testosterone administration on osmoregulation and energy metabolism of gilthead sea bream Sparus auratus. Gen. Comp. Endocrinol. 2006, 149, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Munakata, A.; Kobayashi, M. Endocrine control of sexual behavior in teleost fish. Gen. Comp. Endocrinol. 2010, 165, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Lokman, P.M.; Vermeulen, G.J.; Lambert, J.G.D.; Young, G. Gonad histology and plasma steroid profiles in wild New Zealand freshwater eels (Anguilla dieffenbachii and A. australis) before and at the onset of the natural spawning migration. I. Females. Fish Physiol. Biochem. 1998, 19, 325–338. [Google Scholar] [CrossRef]
- Lokman, P.M.; Young, G. Gonad histology and plasma steroid profiles in wild New Zealand freshwater eels (Anguilla dieffenbachii and A. australis) before and at the onset of the natural spawning migration. II. Males. Fish Physiol Biochem. 1998, 19, 339–347. [Google Scholar] [CrossRef]
- Han, Y.S.; Liao, I.C.; Tzeng, W.N.; Huang, Y.S.; Yu, J.Y.L. Serum estradiol-17 and testosterone levels during silvering in wild Japanese eel Anguilla japonica. Comp. Biochem. Physiol. B 2003, 136, 913–920. [Google Scholar] [CrossRef]
- Olivereau, M.; Olivereau, J. Effects of 17 a-methyltestosterone on the skin and gonads of freshwater male silver eels. Gen. Comp. Endocrinol. 1985, 57, 64–71. [Google Scholar] [CrossRef]
- Böetius, I.; Larsen, L.O. Effects of testosterone on eye size and spermiation in silver eels, Anguilla anguilla. Gen. Comp. Endocrinol. 1991, 82, 238. [Google Scholar]
- Rohr, D.H.; Lokman, P.M.; Davie, P.S.; Young, G. 11-Ketotestosterone induces silvering-related changes in immature female short-finned eels, Anguilla australis. Comp. Biochem. Physiol. A 2001, 130, 701–714. [Google Scholar] [CrossRef]
- Vøllestad, L.A. Temperature dependent activity of brackish water yellow eels Anguilla anguilla. Aquacult. Fish. Manag. 1986, 17, 201–206. [Google Scholar] [CrossRef]
- Durif, C.M.F.; Elie, P. Predicting downstream migration of silver eels in a large river catchment based on commercial fishery data. Fish. Manag. Ecol. 2008, 15, 127–137. [Google Scholar] [CrossRef]
- Haro, A.; Castro-Santos, T.; McLaughlin, L.; Whalen, K.; Wipplehauser, G. Stimulation of the influence of hydroelectric project operation on mortality of American eels. In Biology, Management, and Protection of Catadromous Eels, American Fisheries Society Symposium 33; Dixon, D., Ed.; The American Fisheries Society: Bethesda, MD, USA, 2002; pp. 357–365. [Google Scholar]
- Okamura, A.; Yamada, Y.; Tanaka, S.; Horie, N.; Utoh, T.; Mikawa, N.; Akazawa, A.; Oka, P.O. Atmospheric depression as the final trigger for the seaward migration of the Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 2002, 234, 281–288. [Google Scholar] [CrossRef]
- Sudo, R.; Okamura, A.; Fukuda, N.; Miller, M.J.; Tsukamoto, K. Environmental factors affecting the onset of spawning migrations of Japanese eels (Anguilla japonica) in Mikawa Bay Japan. Envrion. Biol. Fish. 2017, 100, 237–249. [Google Scholar] [CrossRef]
- Sudo, R.; Tosaka, R.; Ijiri, S.; Adachi, S.; Suetake, H.; Suzuki, Y.; Horie, N.; Tanaka, S.; Aoyama, J.; Tsukamoto, K. The effect of temperature decrease on oocyte development, sex steroids and gonadotropin-subunit mRNA expression levels in female Japanese eels Anguilla japonica. Fish. Sci. 2011, 77, 575–582. [Google Scholar] [CrossRef]
- Helm, B. Zugunruhe of migratory and non-migratory birds in a circannual context. J. Avian Biol. 2006, 37, 533–540. [Google Scholar] [CrossRef]
- Sudo, R.; Tsukamoto, K. Migratory restlessness and the role of androgen for increasing behavioral drive in the spawning migration of the Japanese eel. Sci. Rep. 2015, 5, 17430. [Google Scholar] [CrossRef] [Green Version]
- Setiawan, A.N.; Wylie, M.J.; Forbes, E.L.; Lokman, P.M. The effect of 11-ketotesotrerone on occupation of downstream location and seawater in the New Zealand Shortfinned eel, Anguilla australis. Zool. Sci. 2012, 29, 1–5. [Google Scholar] [CrossRef]
- Weltzien, F.A.; Pasqualini, C.; Sebert, M.E.; Vidal, B.; Le Belle, N.; Kah, O.; Vernier, P.; Dufour, S. Androgen-dependent stimulation of brain dopaminergic systems in the female European eel (Anguilla anguilla). Endocrinology 2006, 147, 2964–2973. [Google Scholar] [CrossRef] [Green Version]
- Aida, K.; Kato, T.; Awaji, M. Effects of castration on the smoltification of precocious male masu salmon Oncorhynchus masou. Bull. Jpn. Soc. Fish. 1984, 50, 565–571. [Google Scholar] [CrossRef]
- Machidori, S.; Katou, F. Spawning populations and marine life of masu salmon (Oncorhynchus masou). Int. North Pac. Fish Comm. Bull. 1984, 43, 138. [Google Scholar]
- Kiso, K. The life history of masu salmon Oncorhynchus masou originated from rivers of the Pacific coast of northern Honshu, Japan. Bull. Natl. Res. Inst. Fish Sci. 1995, 7, 1–188. [Google Scholar]
- Berglund, I.; Lundqvist, H.; Fängstam, H. Downstream migration of immature salmon (Salmo salar) smolts blocked by implantation of androgen 11-ketoandrostenedione. Aquaculture 1994, 121, 269–276. [Google Scholar] [CrossRef]
- Munakata, A.; Amano, M.; Ikuta, K.; Kitamura, S.; Aida, K. Inhibitory effects of testosterone on downstream migratory behavior in masu salmon, Oncorhynchus masou. Zool. Sci. 2000, 17, 863–870. [Google Scholar] [CrossRef] [Green Version]
- de Jesus, E.G.; Hirano, T.; Inui, Y. Changes in cortisol and thyroid hormone concentrations during early development and metamorphosis in the Japanese eel flounder, Paralichthys olivaceus. Gen. Comp. Endocrinol. 1991, 82, 369–376. [Google Scholar] [CrossRef]
- Yada, T.; Abe, M.; Kaifu, K.; Yokouchi, K.; Fukuda, N.; Kodama, S.; Hakoyama, H.; Ogoshi, M.; Kaiya, H.; Sakamoto, T.; et al. Ghrelin and food acquisition in wild and cultured Japanese eel (Anguilla japonica). Comp. Biochem. Physiol. A 2020, 245, 110700. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudo, R.; Yada, T. Anguillid Eels as a Model Species for Understanding Endocrinological Influences on the Onset of Spawning Migration of Fishes. Biology 2022, 11, 934. https://doi.org/10.3390/biology11060934
Sudo R, Yada T. Anguillid Eels as a Model Species for Understanding Endocrinological Influences on the Onset of Spawning Migration of Fishes. Biology. 2022; 11(6):934. https://doi.org/10.3390/biology11060934
Chicago/Turabian StyleSudo, Ryusuke, and Takashi Yada. 2022. "Anguillid Eels as a Model Species for Understanding Endocrinological Influences on the Onset of Spawning Migration of Fishes" Biology 11, no. 6: 934. https://doi.org/10.3390/biology11060934
APA StyleSudo, R., & Yada, T. (2022). Anguillid Eels as a Model Species for Understanding Endocrinological Influences on the Onset of Spawning Migration of Fishes. Biology, 11(6), 934. https://doi.org/10.3390/biology11060934