
In Vitro Model of Human Skeletal Muscle Tissue for the Study of Resident Macrophages and Stem Cells

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Specimens
2.2. Maintenance and Stimulation of Human Skeletal Muscle Tissue
2.3. Hematoxylin and Eosin (HE) Staining
2.4. Immunofluorescence and DAPI Staining
2.5. Microscopy and Imaging
2.6. Statistical Analyses
3. Results
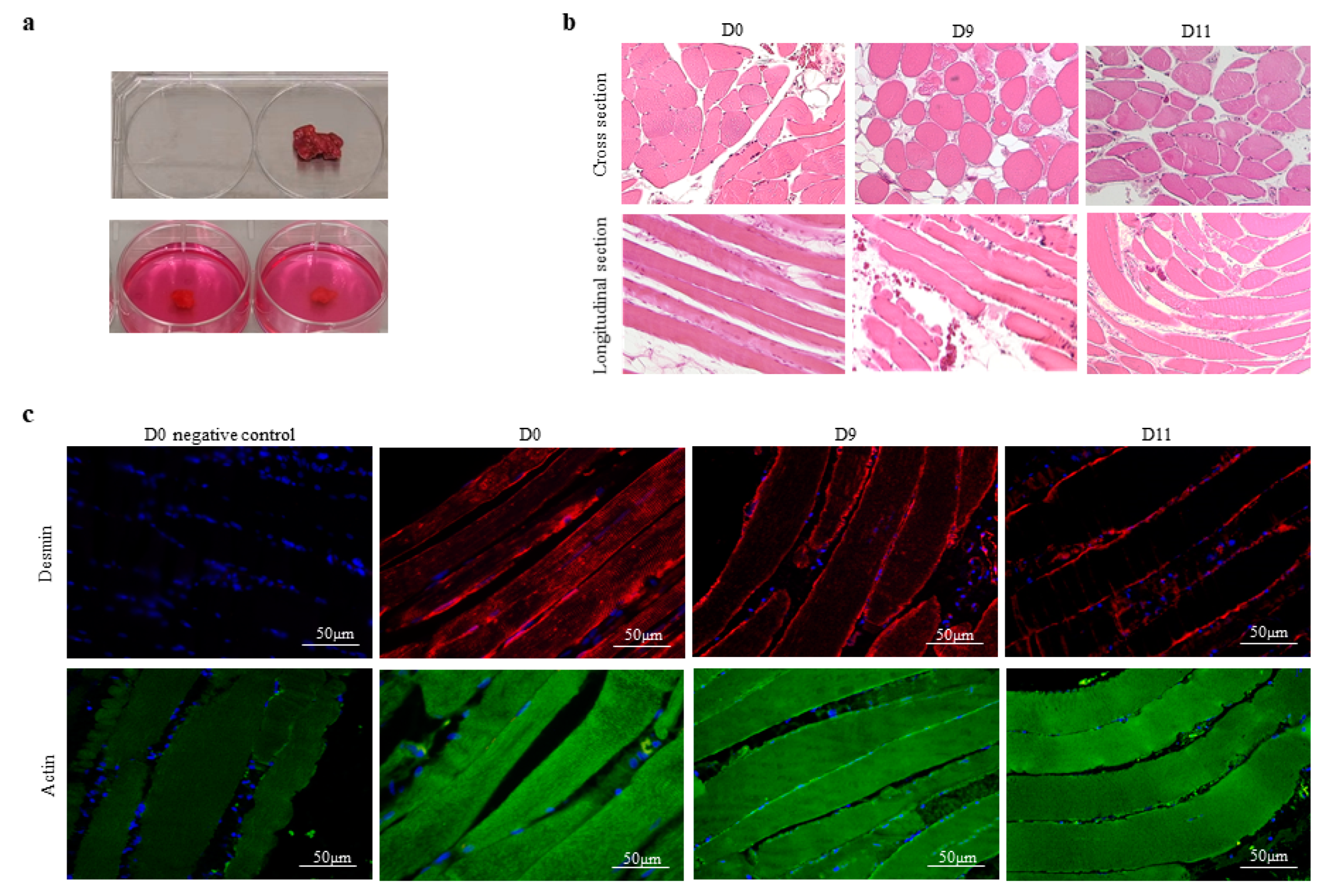
3.1. The Integrity of Human Skeletal Muscle Tissue under Experimental Conditions In Vitro
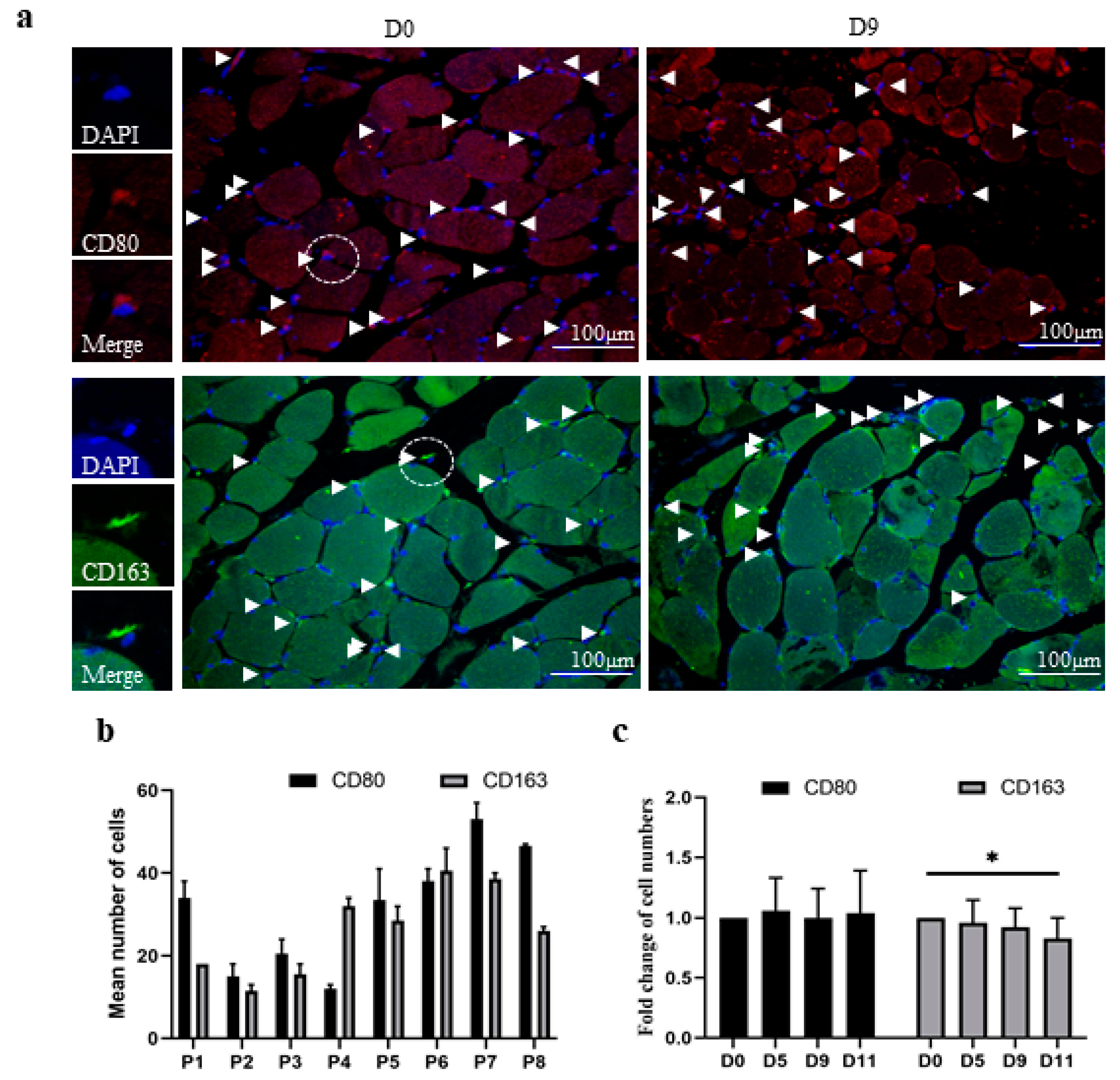
3.2. The In Vitro Model of Human Skeletal Muscle Maintains the Polarization Profile of Resident Macrophages
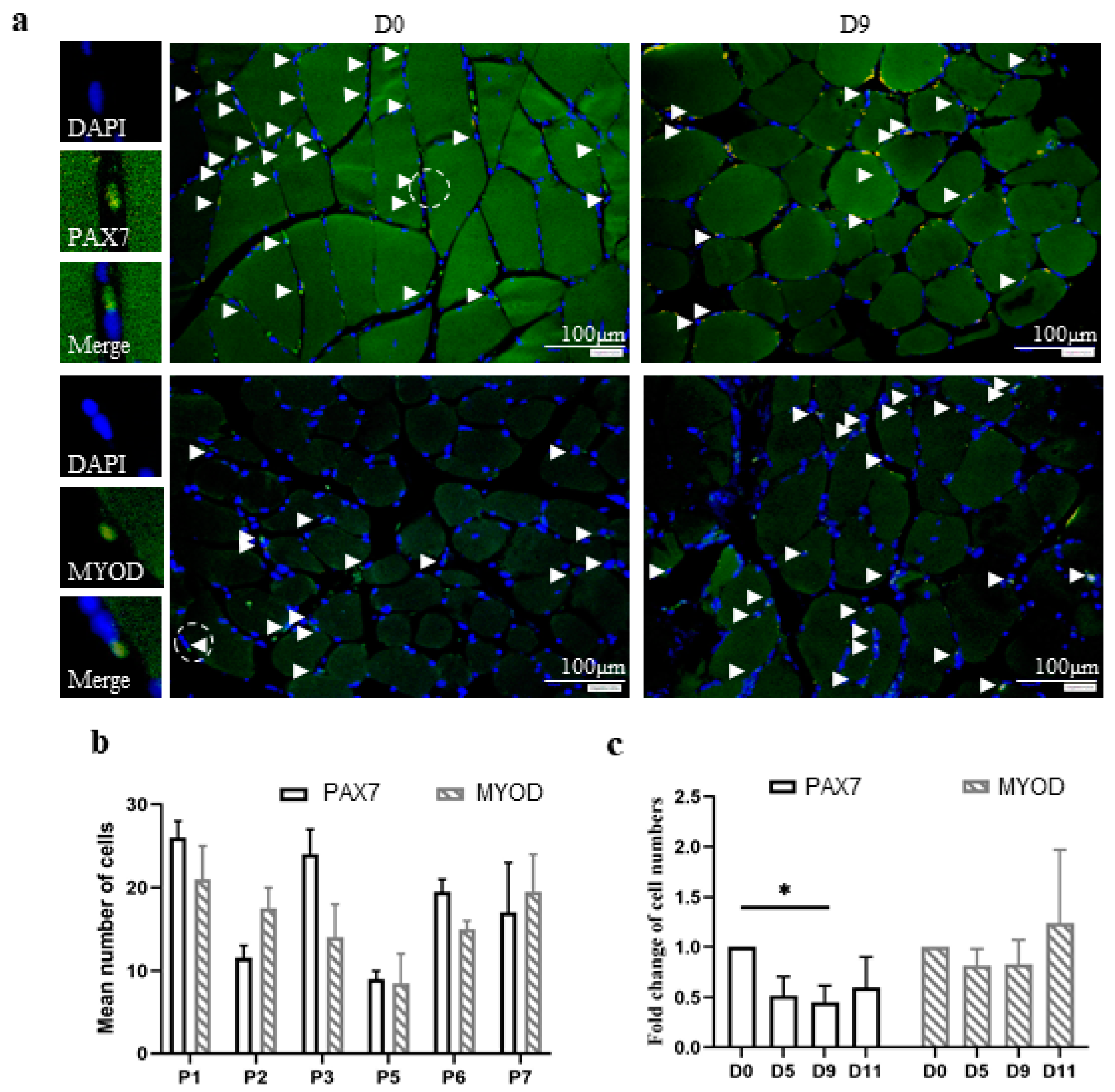
3.3. The In Vitro Model of Human Skeletal Muscle Maintains the Number and Differentiation of Skeletal Muscle Stem Cells
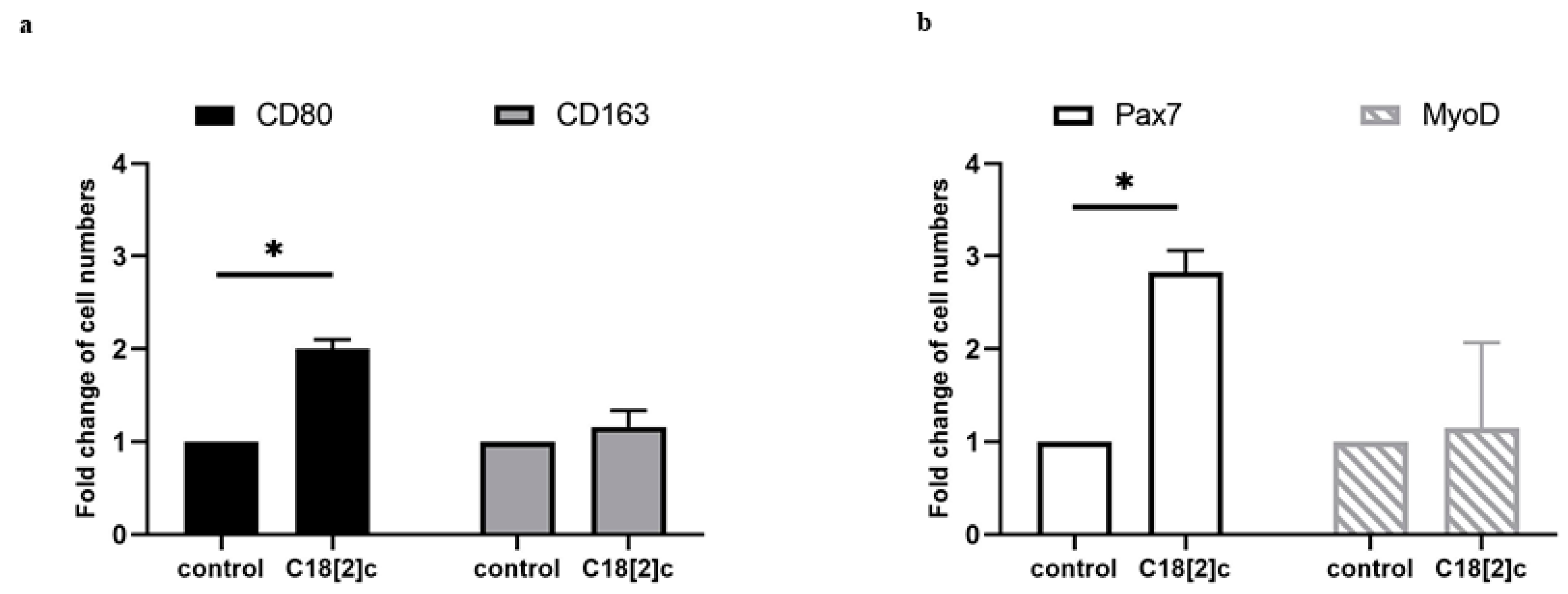
3.4. Unsaturated C18 Fatty Acid Activates M1 Macrophages and Stem Cells in the Human Skeletal Muscle Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, M.; Cappellesso, F.; Amorim, R.; Serneels, J.; Virga, F.; Eelen, G.; Carobbio, S.; Rincon, M.Y.; Maechler, P.; De Bock, K.; et al. Macrophage-derived glutamine boosts satellite cells and muscle regeneration. Nature 2020, 587, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Chazaud, B. Inflammation and Skeletal Muscle Regeneration: Leave It to the Macrophages! Trends Immunol. 2020, 41, 481–492. [Google Scholar] [CrossRef] [PubMed]
- van Doorslaer de Ten Ryen, S.; Francaux, M.; Deldicque, L. Regulation of satellite cells by exercise in hypoxic conditions: A narrative review. Eur. J. Appl. Physiol. 2021, 121, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, C.L.; Fealy, C.E.; Erickson, M.L.; Davuluri, G.; Fujioka, H.; Dantas, W.S.; Huang, E.; Pergola, K.; Mey, J.T.; King, W.T.; et al. Lipids activate skeletal muscle mitochondrial fission and quality control networks to induce insulin resistance in humans. Metabolism 2021, 121, 154803. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liao, C.; Chen, X.; Xu, Y.; Tan, J.; Han, F.; Ye, X. Identification and Characterization of Skeletal Muscle Stem Cells from Human Orbicularis Oculi Muscle. Tissue Eng. Part C Methods 2018, 24, 486–493. [Google Scholar] [CrossRef]
- Parker, L.; Caldow, M.K.; Watts, R.; Levinger, P.; Cameron-Smith, D.; Levinger, I. Age and sex differences in human skeletal muscle fibrosis markers and transforming growth factor-beta signaling. Eur. J. Appl. Physiol. 2017, 117, 1463–1472. [Google Scholar] [CrossRef]
- Sardon Puig, L.; Pillon, N.J.; Naslund, E.; Krook, A.; Zierath, J.R. Influence of obesity, weight loss, and free fatty acids on skeletal muscle clock gene expression. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E1–E10. [Google Scholar] [CrossRef]
- Laurentius, T.; Raffetseder, U.; Fellner, C.; Kob, R.; Nourbakhsh, M.; Floege, J.; Bertsch, T.; Bollheimer, L.C.; Ostendorf, T. High-fat diet-induced obesity causes an inflammatory microenvironment in the kidneys of aging Long-Evans rats. J. Inflamm. 2019, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Kob, R.; Fellner, C.; Bertsch, T.; Wittmann, A.; Mishura, D.; Sieber, C.C.; Fischer, B.E.; Stroszczynski, C.; Bollheimer, C.L. Gender-specific differences in the development of sarcopenia in the rodent model of the ageing high-fat rat. J. Cachexia Sarcopenia Muscle 2015, 6, 181–191. [Google Scholar] [CrossRef]
- Rauen, M.; Hao, D.; Muller, A.; Muckter, E.; Bollheimer, L.C.; Nourbakhsh, M. Free Fatty Acid Species Differentially Modulate the Inflammatory Gene Response in Primary Human Skeletal Myoblasts. Biology 2021, 10, 1318. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.Y.; Driscoll, R.K.; Piao, Y.; Chia, C.W.; Gorospe, M.; Ferrucci, L. Skewed macrophage polarization in aging skeletal muscle. Aging Cell 2019, 18, e13032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Dick, S.A.; Wong, A.; Hamidzada, H.; Nejat, S.; Nechanitzky, R.; Vohra, S.; Mueller, B.; Zaman, R.; Kantores, C.; Aronoff, L.; et al. Three tissue resident macrophage subsets coexist across organs with conserved origins and life cycles. Sci. Immunol. 2022, 7, eabf7777. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Hirschi, K.K. Tissue-Resident Macrophage Development and Function. Front Cell Dev. Biol. 2020, 8, 617879. [Google Scholar] [CrossRef]
- Ishida, J.; Saitoh, M.; Doehner, W.; von Haehling, S.; Anker, M.; Anker, S.D.; Springer, J. Animal models of cachexia and sarcopenia in chronic illness: Cardiac function, body composition changes and therapeutic results. Int. J. Cardiol. 2017, 238, 12–18. [Google Scholar] [CrossRef]
- Christian, C.J.; Benian, G.M. Animal models of sarcopenia. Aging Cell 2020, 19, e13223. [Google Scholar] [CrossRef]
- Nagashima, T.; Hadiwidjaja, S.; Ohsumi, S.; Murata, A.; Hisada, T.; Kato, R.; Okada, Y.; Honda, H.; Shimizu, K. In Vitro Model of Human Skeletal Muscle Tissues with Contractility Fabricated by Immortalized Human Myogenic Cells. Adv. Biosyst. 2020, 4, e2000121. [Google Scholar] [CrossRef]
- Urciuolo, A.; Serena, E.; Ghua, R.; Zatti, S.; Giomo, M.; Mattei, N.; Vetralla, M.; Selmin, G.; Luni, C.; Vitulo, N.; et al. Engineering a 3D in vitro model of human skeletal muscle at the single fiber scale. PLoS ONE 2020, 15, e0232081. [Google Scholar] [CrossRef] [PubMed]
- Wibberley, A.; Staunton, C.A.; Feetham, C.H.; Vereninov, A.A.; Barrett-Jolley, R. An in vitro model of skeletal muscle volume regulation. PLoS ONE 2015, 10, e0127889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squire, J. Special Issue: The Actin-Myosin Interaction in Muscle: Background and Overview. Int. J. Mol. Sci. 2019, 20, 5715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulin, D.; Li, Z. Desmin: A major intermediate filament protein essential for the structural integrity and function of muscle. Exp. Cell Res. 2004, 301, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Atale, N.; Chakraborty, M.; Mohanty, S.; Bhattacharya, S.; Nigam, D.; Sharma, M.; Rani, V. Cardioprotective role of Syzygium cumini against glucose-induced oxidative stress in H9C2 cardiac myocytes. Cardiovasc. Toxicol. 2013, 13, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, K.; Monkawa, T.; Akiyama, T.; Nakatake, Y.; Oda, M.; Goparaju, S.K.; Kimura, H.; Chikazawa-Nohtomi, N.; Sato, S.; Ishiguro, K.; et al. Induction of human pluripotent stem cells into kidney tissues by synthetic mRNAs encoding transcription factors. Sci. Rep. 2019, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, K.M.; Clark, N.M.; Olivares-Navarrete, R. Macrophage response to hydrophilic biomaterials regulates MSC recruitment and T-helper cell populations. Biomaterials 2018, 182, 202–215. [Google Scholar] [CrossRef]
- Vogelpoel, L.T.; Hansen, I.S.; Rispens, T.; Muller, F.J.; van Capel, T.M.; Turina, M.C.; Vos, J.B.; Baeten, D.L.; Kapsenberg, M.L.; de Jong, E.C.; et al. Fc gamma receptor-TLR cross-talk elicits pro-inflammatory cytokine production by human M2 macrophages. Nat. Commun. 2014, 5, 5444. [Google Scholar] [CrossRef] [Green Version]
- Kosmac, K.; Peck, B.D.; Walton, R.G.; Mula, J.; Kern, P.A.; Bamman, M.M.; Dennis, R.A.; Jacobs, C.A.; Lattermann, C.; Johnson, D.L.; et al. Immunohistochemical Identification of Human Skeletal Muscle Macrophages. Bio-protocol 2018, 8, e2883. [Google Scholar] [CrossRef] [Green Version]
- Varga, T.; Mounier, R.; Horvath, A.; Cuvellier, S.; Dumont, F.; Poliska, S.; Ardjoune, H.; Juban, G.; Nagy, L.; Chazaud, B. Highly Dynamic Transcriptional Signature of Distinct Macrophage Subsets during Sterile Inflammation, Resolution, and Tissue Repair. J. Immunol. 2016, 196, 4771–4782. [Google Scholar] [CrossRef] [Green Version]
- Patsouris, D.; Cao, J.J.; Vial, G.; Bravard, A.; Lefai, E.; Durand, A.; Durand, C.; Chauvin, M.A.; Laugerette, F.; Debard, C.; et al. Insulin resistance is associated with MCP1-mediated macrophage accumulation in skeletal muscle in mice and humans. PLoS ONE 2014, 9, e110653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Ballantyne, C.M. Skeletal muscle inflammation and insulin resistance in obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Morales, F.E.; IglayReger, H.B.; Treutelaar, M.K.; Rothberg, A.E.; Hubal, M.J.; Nadler, E.P.; Robidoux, J.; Barakat, H.; Horowitz, J.F.; et al. Expression of macrophage genes within skeletal muscle correlates inversely with adiposity and insulin resistance in humans. Appl. Physiol. Nutr. Metab. 2018, 43, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amouzou, C.; Breuker, C.; Fabre, O.; Bourret, A.; Lambert, K.; Birot, O.; Fédou, C.; Dupuy, A.M.; Cristol, J.P.; Sutra, T.; et al. Skeletal Muscle Insulin Resistance and Absence of Inflammation Characterize Insulin-Resistant Grade I Obese Women. PLoS ONE 2016, 11, e0154119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, C.S.; Sparks, L.M.; Johannsen, D.L.; Covington, J.D.; Church, T.S.; Ravussin, E. Low macrophage accumulation in skeletal muscle of obese type 2 diabetics and elderly subjects. Obesity 2012, 20, 1530–1533. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, J.R.; Kaluhiokalani, J.P.; Hafen, P.S.; Deyhle, M.R.; Parcell, A.C.; Hyldahl, R.D. An altered response in macrophage phenotype following damage in aged human skeletal muscle: Implications for skeletal muscle repair. FASEB J. 2019, 33, 10353–10368. [Google Scholar] [CrossRef] [Green Version]
- Haizlip, K.M.; Harrison, B.C.; Leinwand, L.A. Sex-based differences in skeletal muscle kinetics and fiber-type composition. Physiology 2015, 30, 30–39. [Google Scholar] [CrossRef]
- Marg, A.; Escobar, H.; Gloy, S.; Kufeld, M.; Zacher, J.; Spuler, A.; Birchmeier, C.; Izsvák, Z.; Spuler, S. Human satellite cells have regenerative capacity and are genetically manipulable. J. Clin. Investig. 2014, 124, 4257–4265. [Google Scholar] [CrossRef] [Green Version]
- Snijders, T.; Nederveen, J.P.; McKay, B.R.; Joanisse, S.; Verdijk, L.B.; van Loon, L.J.C.; Parise, G. Satellite cells in human skeletal muscle plasticity. Front. Physiol. 2015, 6, 283. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Wang, H.; Hu, P. Stem cell activation in skeletal muscle regeneration. Cell. Mol. Life Sci. 2015, 72, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; Geles, K.G.; Paik, J.H.; DePinho, R.A.; Tjian, R. Codependent activators direct myoblast-specific MyoD transcription. Dev. Cell 2008, 15, 534–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riuzzi, F.; Sorci, G.; Sagheddu, R.; Sidoni, A.; Alaggio, R.; Ninfo, V.; Donato, R. RAGE signaling deficiency in rhabdomyosarcoma cells causes upregulation of PAX7 and uncontrolled proliferation. J. Cell Sci. 2014, 127 Pt 8, 1699–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Sex | Age (Years) | BMI (kg/m2) | Type 2 Diabetes | Source |
|---|---|---|---|---|---|
| P1 | female | 73 | 26.6 | Yes | gluteus medius muscle |
| P2 | female | 82 | 26 | Yes | gluteus medius muscle |
| P3 | female | 68 | >40 | Yes | deltoid muscle |
| P4 | female | 76 | 27.3 | No | multifidus muscle |
| P5 | female | 82 | 26.1 | No | pronator quadratus muscle |
| P6 | male | 72 | 37.5 | No | vastus lateralis muscle |
| P7 | female | 81 | 25.6 | No | pronator quadratus muscle |
| P8 | female | 68 | 24.8 | No | vastus lateralis muscle |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, D.; Becker, N.; Mückter, E.; Müller, A.; Pishnamaz, M.; Bollheimer, L.C.; Hildebrand, F.; Nourbakhsh, M. In Vitro Model of Human Skeletal Muscle Tissue for the Study of Resident Macrophages and Stem Cells. Biology 2022, 11, 936. https://doi.org/10.3390/biology11060936
Hao D, Becker N, Mückter E, Müller A, Pishnamaz M, Bollheimer LC, Hildebrand F, Nourbakhsh M. In Vitro Model of Human Skeletal Muscle Tissue for the Study of Resident Macrophages and Stem Cells. Biology. 2022; 11(6):936. https://doi.org/10.3390/biology11060936
Chicago/Turabian StyleHao, Dandan, Nils Becker, Eva Mückter, Aline Müller, Miguel Pishnamaz, Leo Cornelis Bollheimer, Frank Hildebrand, and Mahtab Nourbakhsh. 2022. "In Vitro Model of Human Skeletal Muscle Tissue for the Study of Resident Macrophages and Stem Cells" Biology 11, no. 6: 936. https://doi.org/10.3390/biology11060936
APA StyleHao, D., Becker, N., Mückter, E., Müller, A., Pishnamaz, M., Bollheimer, L. C., Hildebrand, F., & Nourbakhsh, M. (2022). In Vitro Model of Human Skeletal Muscle Tissue for the Study of Resident Macrophages and Stem Cells. Biology, 11(6), 936. https://doi.org/10.3390/biology11060936