
Effects of Extreme Light Cycle and Density on Melatonin, Appetite, and Energy Metabolism of the Soft-Shelled Turtle (Pelodiscus sinensis)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Acclimation
2.2. Experimental Protocol
2.3. Metabolic Rates Measurement
2.4. Sampling
2.5. Calculations
2.6. Analysis of Plasma Hormones
2.7. Total RNA Extraction, First-Strand cDNA Synthesis, and Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Plasma Melatonin Level
3.2. Growth and Food Intake
3.3. Appetite Regulation-Related Gene Expressions and Plasma Leptin and Ghrelin Levels
3.4. SMR
3.5. Plasma Growth Hormone Level
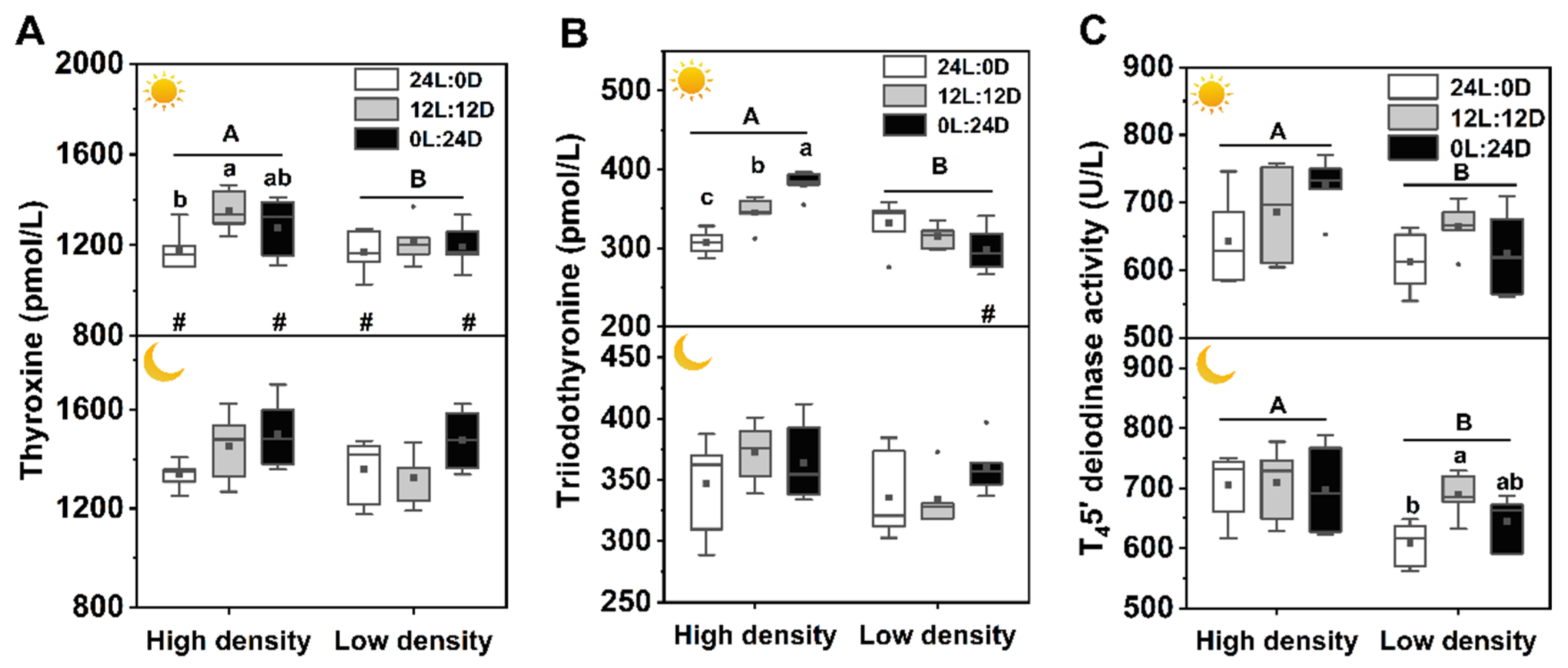
3.6. Thyroid Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumari, R.; Verma, V.; Kronfeld-Schor, N.; Singaravel, M. Differential response of diurnal and nocturnal mammals to prolonged altered light-dark cycle: A possible role of mood associated endocrine, inflammatory and antioxidant system. Chronobiol. Int. 2021, 38, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A. Chronic exposure to continuous brightness or darkness modulates immune responses and ameliorates the antioxidant enzyme system in male rats. Front. Vet. Sci. 2021, 8, 621188. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Zhao, J.; Ji, B.M.; Zhang, Y.D.; Ye, Z.Y.; Zhu, S.M. Effects of photoperiod on growth, digestive enzyme activity, stress, and oxidative status of juvenile Chinese soft-shelled turtles (Pelodiscus sinensis) in a greenhouse. Trans. ASABE 2020, 63, 1787–1793. [Google Scholar] [CrossRef]
- Burraco, P.; Valdés, A.E.; Orizaola, G.; Ardia, D. Metabolic costs of altered growth trajectories across life transitions in amphibians. J. Anim. Ecol. 2019, 89, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Q.; Niu, C.J.; Li, Q.F.; Ma, H.F. The effects of light intensity on daily food consumption and specific growth rate of the juvenile soft-shelled turtle, Trionyx sinensis. Acta Zool. Sin. 1998, 44, 157–161. [Google Scholar]
- Albarrán, M.T.; López-Burillo, S.; Pablos, M.I.; Reíter, R.J.; Agapito, M.T. Endogenous rhythms of melatonin, total antioxidant status and superoxide dismutase activity in several tissues of chick and their inhibition by light. J. Pineal Res. 2001, 30, 227–233. [Google Scholar] [CrossRef]
- Piccinetti, C.C.; Migliarini, B.; Olivotto, I.; Coletti, G.; Amici, A.; Carnevali, O. Appetite regulation: The central role of melatonin in Danio rerio. Horm. Behav. 2010, 58, 6. [Google Scholar] [CrossRef]
- Reiter, R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef] [Green Version]
- Prusik, M.; Lewczuk, B. Diurnal rhythm of plasma melatonin concentration in the domestic turkey and its regulation by light and endogenous oscillators. Animals 2020, 10, 678. [Google Scholar] [CrossRef]
- Firth, B.T.; Belan, I.; Kennaway, D.J. Persistence of a plasma melatonin rhythm in constant darkness and its inhibition by constant light in the sleepy lizard, Tiliqua rugosa. J. Pineal Res. 2006, 41, 15–20. [Google Scholar] [CrossRef]
- Murakami, N.; Nakamura, H.; Nishi, R.; Marumoto, N.; Nasu, T. Comparison of circadian oscillation of melatonin release in pineal cells of house sparrow, pigeon and Janpanese quail, using cell perfusion system. Brain Res. 1994, 651, 209–214. [Google Scholar] [CrossRef]
- Iigo, M.; Azuma, T.; Iwata, M. Lack of Circadian Regulation of Melatonin Rhythms in the Sockeye Salmon (Oncorhynchus nerka) in vivoandin vitro. Zool. Sci. 2007, 24, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Iigo, M.M.; Kitamurai, S.S.; Ikuta, K.K.; Sánchez-Vázquezi, F.J.F.J.; Ohtani-Kanekoi, R.R.; Hara, M.M.; Hirata, K.K.; Tabata, M.M.; Aida, K.K. Regulation by Light and Darkness of Melatonin Secretion from the Superfused Masu Salmon (Oncorhynchus masou) Pineal Organ. Biol. Rhythm Res. 1998, 29, 86–97. [Google Scholar] [CrossRef]
- De Pedro, N.; Martínez-Álvarez, R.M.; Delgado, M.J. Melatonin reduces body weight in goldfish (Carassius auratus) effects on metabolic resoures and some feeding regulators. J. Pineal Res. 2008, 45, 8. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.; Afeche, S.C.; Tan, D.X.; Reiter, R.J. Melatonin, energy metabolism, and obesity a review. J. Pineal Res. 2014, 56, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, A.; Topal, T.; Tan, D.X.; Reiter, R.J. Role of melatonin in metabolic regulation. Rev. Endocr. Metab. Disord. 2009, 10, 261–270. [Google Scholar] [CrossRef]
- Gandhi, A.V.; Mosser, E.A.; Oikonomou, G.; Prober, D.A. Melatonin is required for the circadian regulation of sleep. Neuron 2015, 85, 1193–1199. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martín, V.; Reiter, R.J. Regulation of antioxidant enzymes a significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Yang, C.H.; Xu, J.H.; Ren, Q.C.; Duan, T.; Mo, F.; Zhang, W. Melatonin promotes secondary hair follicle development of early postnatal cashmere goat and improves cashmere quantity and quality by enhancing antioxidant capacity and suppressing apoptosis. J. Pineal Res. 2019, 67, e12569. [Google Scholar] [CrossRef]
- Carrillo-Vico, A.; Guerrero, J.M.; Lardone, P.J.; Reiter, R.J. A review of the multiple actions of melatonin on the immune system. Endocrine 2005, 27, 189–200. [Google Scholar] [CrossRef]
- Giannetto, C.; Carcangiu, V.; Luridiana, S.; Parmeggiani, A.; Piccione, G. Twenty-four-hour rhythm patterns of plasma melatonin in short-day and long-day breeders maintained under natural environmental conditions. Chronobiol. Int. 2020, 37, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.W.; Johnson, C.E.; Bell, L.M.; Reilly, L.; Duncan, J.S.; Barrett, P.; Heideman, P.D.; Morgan, P.J. Divergent regulation of hypothalamic neuropeptide Y and agouti-related protein by photoperiod in F344 rats with differential food intake and growth. J. Neuroendocrinol. 2009, 21, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, H.; Niki, T.; Shiiya, T. Feeding behavior and gene expression of appetite-related neuropeptides in mice lacking for neuropeptide Y Y5 receptor subclass. World J. Gastroenterol. 2008, 14, 6312. [Google Scholar] [CrossRef] [PubMed]
- Boswell, T.; Dunn, I.C. Regulation of agouti-related protein and pro-opiomelanocortin gene expression in the avian arcuate nucleus. Front. Endocrinol. 2017, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schalla, M.A.; Stengel, A. Central mechanisms of kisspeptin-induced inhibition of food intake. Peptides 2021, 135, 170475. [Google Scholar] [CrossRef]
- Cázarez-Márquez, F.; Eliveld, J.; Ritsema, W.I.G.R.; Foppen, E.; Bossenbroek, Y.; Pelizzari, S.; Simonneaux, V.; Kalsbeek, A. Role of central kisspeptin and RFRP-3 in energy metabolism in the male Wistar rat. J. Neuroendocrinol. 2021, 33, e12973. [Google Scholar] [CrossRef]
- Ellacott, K.L.J.; Cone, R.D. The central melanocortin system and the integration of short- and long-term regulators of energy homeostasis. Recent Prog. Horm. Res. 2004, 59, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Wilding, J.P.H. Neuropeptides and appetite control. Diabetic Med. 2002, 19, 619–627. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Librán-Pérez, M.; López Patiño, M.A.; Soengas, J.L.; Míguez, J.M. Melatonin treatment alters glucosensing capacity and mRNA expression levels of peptides related to food intake control in rainbow trout hypothalamus. Gen. Comp. Endocrinol. 2012, 178, 131–138. [Google Scholar] [CrossRef]
- Genin, F.; Schilling, A.; Claustrat, B. Melatonin and methimazole mimic short-day-induced fattening in gray mouse lemurs. Physiol. Behav. 2003, 79, 553–559. [Google Scholar] [CrossRef]
- Mustonen, A.-M.; Nieminen, P.; Asikainen, J.; Saarela, S.; Kukkonen, J.V.K.; Hyvärinen, H. Continuous melatonin treatment and fasting in the Raccoon dog (Nyctereutes procyonoides)—Vernal body weight regulation and reproduction. Zool. Sci. 2004, 21, 163–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.L.; Li, X.; Shi, C.; Wu, F.C.; Song, C.B.; Liu, Y. Effects of stocking density on growth, metabolism, and energy budget of Haliotis discus hannai Ino. Aquaculture 2018, 483, 84–95. [Google Scholar] [CrossRef]
- Oyarzún, R.; Paredes, R.; Saravia, J.; Morera, F.J.; Muñoz, J.L.P.; Ruiz-Jarabo, I.; Mancera, J.M.; Vargas-Chacoff, L. Stocking density affects the growth performance, intermediary metabolism, osmoregulation, and response to stress in Patagonian blennie Eleginops maclovinus. Aquaculture 2020, 515, 734565. [Google Scholar] [CrossRef]
- Jing, R.Z. The Effects of Chronic Ammonia Exposure, Stocking Density and Hibernation on Energy Budget of Juvenile Soft-shelled Turtle, and the Function of Dietary Vitamin C on Hibernation. Ph.D. Thesis, Beijing Normal University, Beijing, China, 2007. [Google Scholar]
- Chen, X.R.; Niu, C.J.; Pu, L.J. Effects of stocking density on growth and non-specific immune responses in juvenile soft-shelled turtle, Pelodiscus sinensis. Aquac. Res. 2007, 38, 1380–1386. [Google Scholar] [CrossRef]
- Wideman, C.H.; Murphy, H.M. Constant light induces alterations in melatonin levels, food intake, feed efficiency, visceral adiposity, and circadian rhythms in rats. Nutr. Neurosci. 2013, 12, 233–240. [Google Scholar] [CrossRef]
- Simms, D.; Cizdziel, P.E.; Chomczynski, P. TRIzol: A new reagent for optimal single-step isolation of RNA. Focus 1993, 15, 99–103. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Vivien-Roels, B.; Arendt, J.; Breadtke, J. Circadian and circannual fluctuations of pineal indoleamines (serotonin and melatonin) in Testudo hermanni gmelin (Reptilia, Chelonia): Under natural conditions of photoperiod and temperature. Gen. Comp. Endocrinol. 1979, 37, 14. [Google Scholar] [CrossRef]
- Hou, Z.S.; Wen, H.S.; Li, J.F.; He, F.; Li, Y.; Qi, X.; Zhao, J.; Zhang, K.Q.; Tao, Y.X. Effects of photoperiod and light Spectrum on growth performance, digestive enzymes, hepatic biochemistry and peripheral hormones in spotted sea bass (Lateolabrax maculatus). Aquaculture 2019, 507, 419–427. [Google Scholar] [CrossRef]
- Wyse, C.A.; Zhang, X.; McLaughlin, M.; Biello, S.M.; Hough, D.; Bellingham, M.; Curtis, A.M.; Robinson, J.E.; Evans, N.P. Circadian rhythms of melatonin and behaviour in juvenile sheep in field conditions: Effects of photoperiod, environment and weaning. Physiol. Behav. 2018, 194, 362–370. [Google Scholar] [CrossRef]
- López-Patiño, M.A.; Gesto, M.; Conde-Sieira, M.; Soengas, J.L.; Míguez, J.M. Stress inhibition of melatonin synthesis in the pineal organ of rainbow trout (Oncorhynchus mykiss) is mediated by cortisol. J. Exp. Biol. 2014, 217, 1407–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikaido, Y.; Aluru, N.; McGuire, A.; Park, Y.-J.; Vijayan, M.M.; Takemura, A. Effect of cortisol on melatonin production by the pineal organ of tilapia, Oreochromis mossambicus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 155, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Sengupta, A.; Chaudhuri-Sengupta, S.; Maiti, B. Thyroid responses to altered photoperiod in the soft-shelled turtle Lissemys punctata punctata Bonnoterre. Acta Biol. Hung. 2007, 58, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Lugo, M.J.; Jiménez-Ortega, V.; Cano-Barquilla, P.; Mateos, P.F.; Spinedi, E.J.; Cardinali, D.P.; Esquifino, A.I. Melatonin counteracts changes in hypothalamic gene expression of signals regulating feeding behavior in high-fat fed rats. Horm. Mol. Biol. Clin. Investig. 2015, 21, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The role of kisspeptin neurons in reproduction and metabolism. J. Endocrinol. 2018, 238, R173–R183. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.W.; Elmquist, J.K.; Elias, C.F. Hypothalamic pathways linking energy balance and reproduction. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E827–E832. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Hosoda, H.; Matsuo, H.; Kangawa, K. Ghrelin: Discovery of the natural endogenous ligand for the growth hormone secretagogue receptor. Trends Endocrinol. Metab. 2001, 12, 118–122. [Google Scholar] [CrossRef]
- Blanton, M.L.; Specker, J.L. The hypothalamic-pituitary-thyroid (HPT) axis in fish and its role in fish development and reproduction. Crit. Rev. Toxicol. 2007, 37, 97–115. [Google Scholar] [CrossRef]
- Olatunji-bello, I.I.; Sofola, O.A. Effect of continous light and darkness exposures on the petuitary-gonadal axis and thyroid activity in male rats. Afr. J. Biomed. Res. 2001, 4, 119–122. [Google Scholar] [CrossRef]
- Singh, D.; Turner, C. Effect of light and darkness upon thyroid secretion rate and on the endocrine lgands of female rats. Proc. Soc. Exp. Biol. Med. 1969, 131, 4. [Google Scholar] [CrossRef]
- Creighton, J.A.; Rudeen, P.K. Effects of melatonin and thyroxine treatment on reproductive organs and thyroid hormone levels in male hamsters. J. Pineal Res. 1989, 6, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Varela, L.; Martínez-Sánchez, N.; Gallego, R.; Vázquez, M.J.; Roa, J.; Gándara, M.; Schoenmakers, E.; Nogueiras, R.; Chatterjee, K.; Tena-Sempere, M.; et al. Hypothalamic mTOR pathway mediates thyroid hormone-induced hyperphagia in hyperthyroidism. J. Pathol. 2012, 227, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Calvino, C.; Império, G.E.; Wilieman, M.; Costa-e-Sousa, R.H.; Souza, L.L.; Trevenzoli, I.H.; Pazos-Moura, C.C. Hypothyroidism induces hypophagia associated with alterations in protein expression of neuropeptide Y and proopiomelanocortin in the arcuate nucleus, independently of hypothalamic nuclei-specific changes in leptin signaling. Thyroid 2016, 26, 134–143. [Google Scholar] [CrossRef] [PubMed]
 and
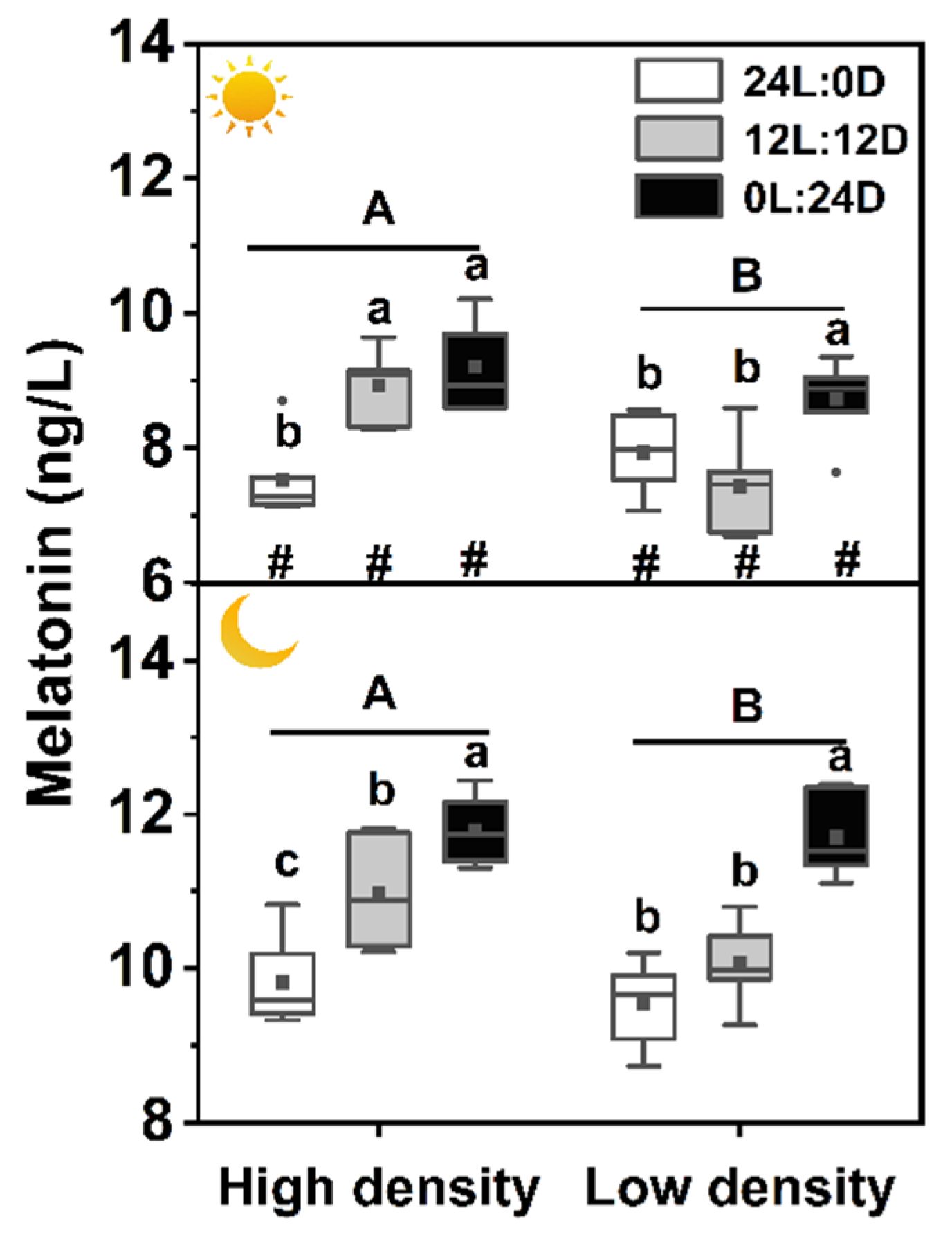
and  indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In the box plots, the solid horizontal indicates median, the filled square indicates the mean, and points indicate outliers. Different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In the box plots, the solid horizontal indicates median, the filled square indicates the mean, and points indicate outliers. Different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In the box plots, the solid horizontal indicates median, the filled square indicates the mean, and points indicate outliers. Different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In the box plots, the solid horizontal indicates median, the filled square indicates the mean, and points indicate outliers. Different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
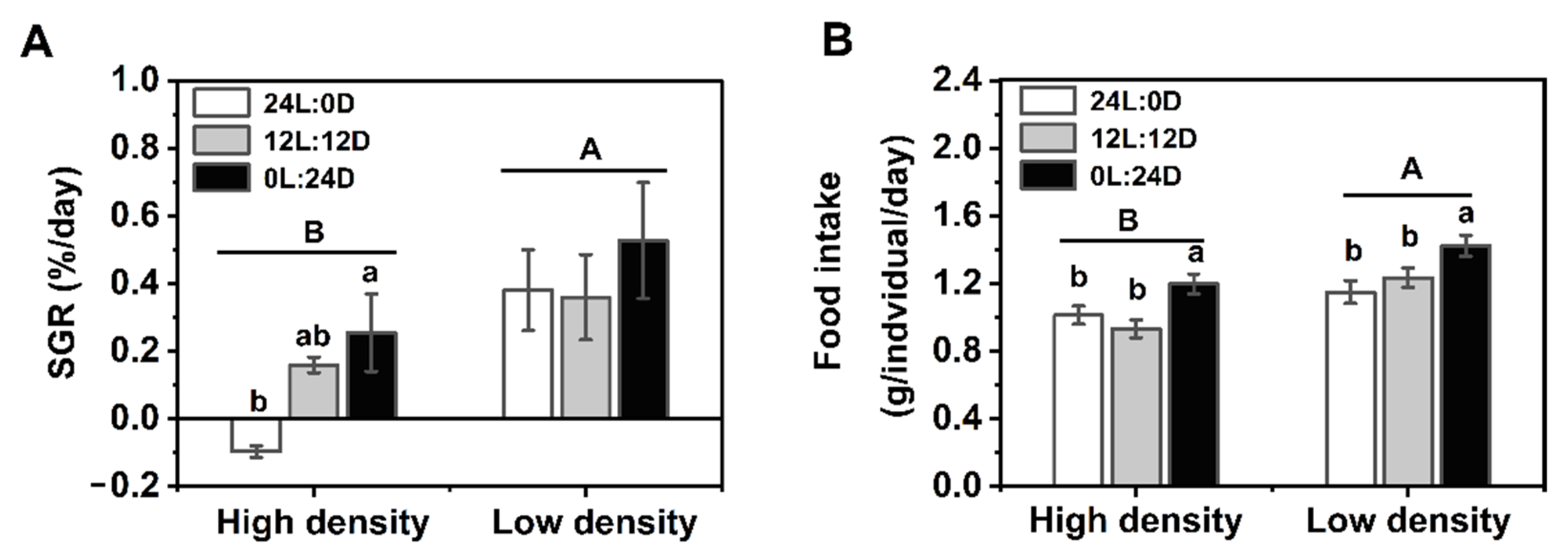
 and indicate daytime and nighttime, respectively. Dates are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In (A)–(C), different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities (p < 0.05). In (D,E), the description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Dates are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In (A)–(C), different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities (p < 0.05). In (D,E), the description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Dates are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In (A)–(C), different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities (p < 0.05). In (D,E), the description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Dates are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. In (A)–(C), different lowercase letters denote statistically significant differences among different photoperiods in each density; different capital letters indicate a significant difference between different densities (p < 0.05). In (D,E), the description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
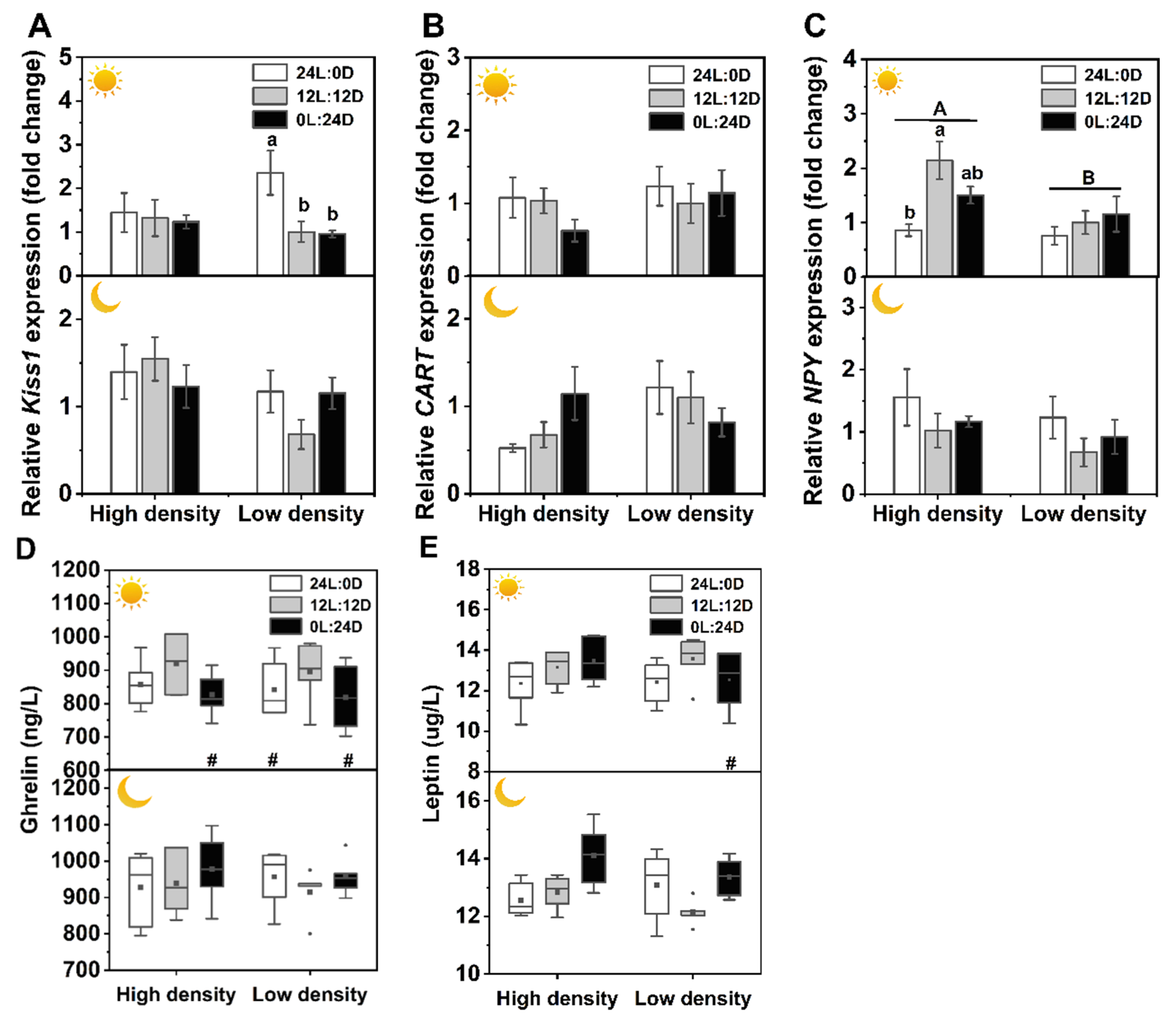
 and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. The description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. The description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. The description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. The description of the box plots is the same as that shown in Figure 1. # indicate a significant difference between the daytime and nighttime (p < 0.05). and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. The description of the box plots is the same as that shown in Figure 1. Different lowercase letters denote statistically significant differences among different photoperiods under each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
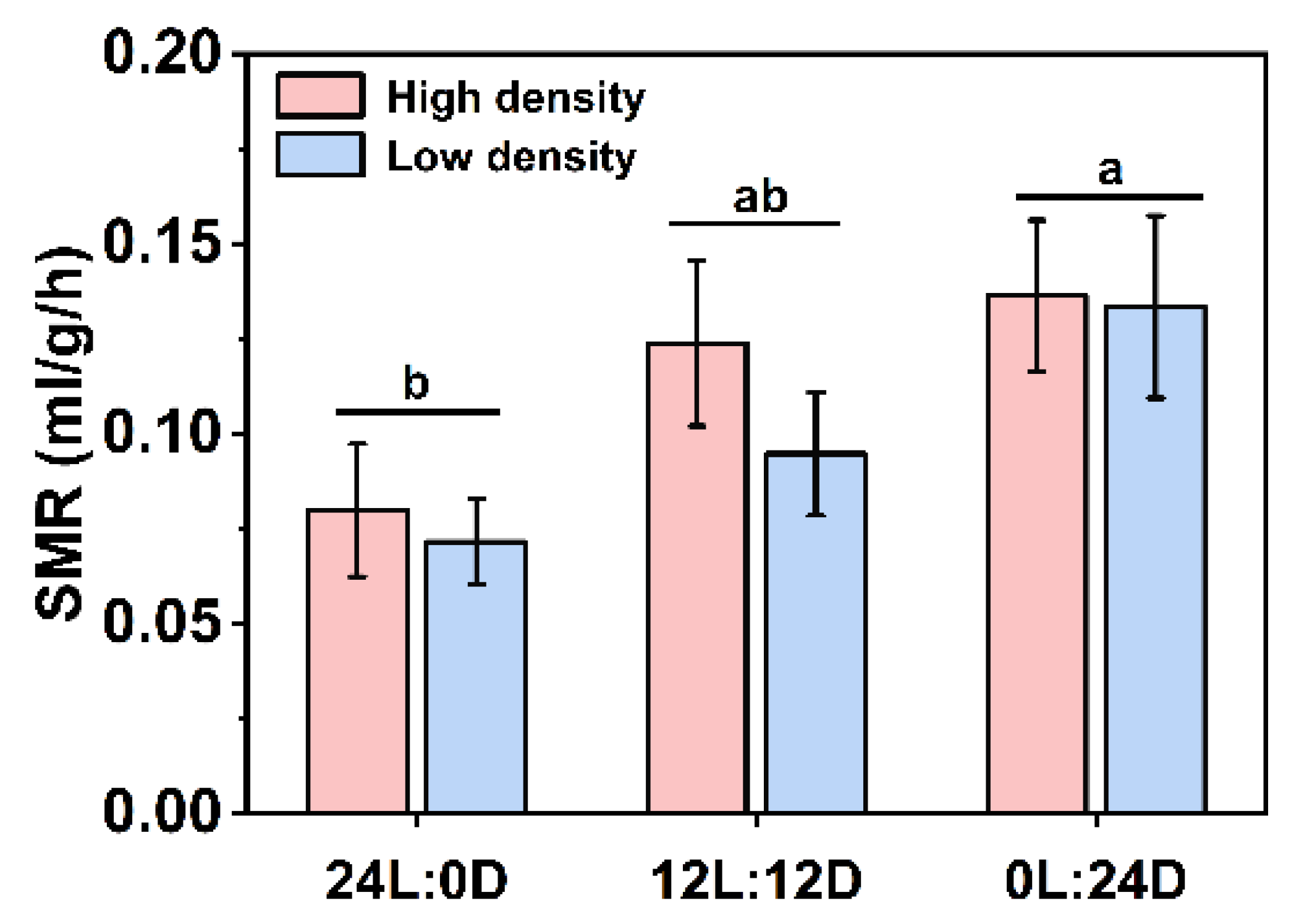
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. The description of the box plots is the same as that shown in Figure 1. Different lowercase letters denote statistically significant differences among different photoperiods under each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. The description of the box plots is the same as that shown in Figure 1. Different lowercase letters denote statistically significant differences among different photoperiods under each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).
and indicate daytime and nighttime, respectively. Data are n = 5 or 6 for all groups. Data are presented as the mean ± S.E. The description of the box plots is the same as that shown in Figure 1. Different lowercase letters denote statistically significant differences among different photoperiods under each density; different capital letters indicate a significant difference between different densities; # indicate a significant difference between the daytime and nighttime (p < 0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession No. | Forward Primer | Reverse Primer | PCR Efficiency |
|---|---|---|---|---|
| GAPDH | NM_001286927.1 | TTCATGGCACTGTCAAGGCT | GGTTGACGCCCATCACAAAC | 97.1% |
| Kiss1 | XM_025189675.1 | GAGTTCCAGTTGTAGGCGGA | GGGAGCCTGCAGATGGATAT | 90.7% |
| CART | XM_025185227.1 | CAAGCGGATCCCCATTTACG | TTCCCGATTCGAGCTCCTTT | 89.0% |
| NPY | XM_006138369.3 | TCATCACGCGGCAGAGATAT | GTCTTCAAACCTAGATCTTGGGA | 97.9% |
| Gene | Photoperiod | Time | Density | Density × Time | Photoperiod × Density | Photoperiod × Time | Density × Photoperiod × Time |
|---|---|---|---|---|---|---|---|
| Melatonin | F2,58 = 45.800 | F1,58 = 274.608 | F1,58 = 11.037 | F1,58 = 0.106 | F2,58 = 7.027 | F2,58 = 2.881 | F2,58 = 1.967 |
| p < 0.001 | p < 0.001 | p = 0.002 | p = 0.746 | p = 0.002 | p = 0.064 | p = 0.149 | |
| Kiss1 | F2,57= 2.440 | F1,57 = 1.319 | F1,57 = 0.461 | F1,57 = 1.583 | F2,57 = 2.555 | F2,57 = 1.727 | F2,57 = 0.961 |
| p = 0.096 | p = 0.256 | p = 0.500 | p = 0.213 | p = 0.087 | p = 0.187 | p = 0.388 | |
| CART | F2,53 = 0.115 | F1,58 = 0.534 | F1,53 = 2.806 | F1,53 = 0.031 | F2,53 = 0.475 | F2,53 = 0.623 | F2,53 = 2.485 |
| p = 0.892 | p = 0.468 | p = 0.100 | p = 0.862 | p = 0.625 | p = 0.540 | p = 0.093 | |
| NPY | F2,53 = 0.620 | F1,53 = 1.776 | F1,53 = 6.945 | F1,53 = 1.230 | F2,53 = 1.982 | F2,53 = 5.911 | F2,53 = 0.719 |
| p = 0.542 | p = 0.188 | p = 0.011 | p = 0.272 | p = 0.148 | p = 0.005 | p = 0.492 | |
| Leptin | F2,58 = 3.504 | F1,58 = 0.157 | F1,58 = 0.933 | F1,58 = 0.099 | F2,58 = 2.001 | F2,58 = 4.180 | F2,58 = 1.064 |
| p = 0.037 | p = 0.694 | p = 0.338 | p = 0.755 | p = 0.144 | p = 0.020 | p = 0.352 | |
| Ghrelin | F2,58 = 0.552 | F1,58 = 19.367 | F1,58 = 0.323 | F1,58 = 0.080 | F2,58 = 0.229 | F2,58 = 3.439 | F2,58 = 0.207 |
| p = 0.579 | p < 0.001 | p = 0.572 | p = 0.778 | p = 0.796 | p = 0.039 | p = 0.814 | |
| GH | F2,58 = 1.858 | F1,58 = 4.712 | F1,58 = 0.085 | F1,58 = 2.467 | F2,58 = 0.061 | F2,58 = 0.603 | F2,58 = 0.977 |
| p = 0.165 | p = 0.034 | p = 0.771 | p = 0.122 | p = 0.941 | p = 0.551 | p = 0.382 | |
| T4 | F2,58 = 5.769 | F1,58 = 48.356 | F1,58 = 5.553 | F1,58 = 0.444 | F2,58 = 2.597 | F2,58 = 2.716 | F2,58 = 0.090 |
| p = 0.005 | p < 0.001 | p = 0.022 | p = 0.508 | p = 0.083 | p = 0.075 | p = 0.914 | |
| T3 | F2,58 = 3.706 | F1,58 = 12.832 | F1,58 = 14.999 | F1,58 = 0.916 | F2,58 = 6.479 | F2,58 = 0.006 | F2,58 = 7.830 |
| p = 0.031 | p = 0.001 | p < 0.001 | p = 0.342 | p = 0.003 | p = 0.994 | p = 0.001 | |
| T45′-deiodinase | F2,58 = 4.303 | F1,58 = 1.617 | F1,58 = 17.497 | F1,58 = 0.065 | F2,58 = 1.703 | F2,58 = 0.607 | F2,58 = 1.660 |
| p = 0.018 | p = 0.209 | p < 0.001 | p = 0.799 | p = 0.191 | p = 0.548 | p = 0.199 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Z.; Liu, S.; Niu, C. Effects of Extreme Light Cycle and Density on Melatonin, Appetite, and Energy Metabolism of the Soft-Shelled Turtle (Pelodiscus sinensis). Biology 2022, 11, 965. https://doi.org/10.3390/biology11070965
Tang Z, Liu S, Niu C. Effects of Extreme Light Cycle and Density on Melatonin, Appetite, and Energy Metabolism of the Soft-Shelled Turtle (Pelodiscus sinensis). Biology. 2022; 11(7):965. https://doi.org/10.3390/biology11070965
Chicago/Turabian StyleTang, Zhonghua, Shifan Liu, and Cuijuan Niu. 2022. "Effects of Extreme Light Cycle and Density on Melatonin, Appetite, and Energy Metabolism of the Soft-Shelled Turtle (Pelodiscus sinensis)" Biology 11, no. 7: 965. https://doi.org/10.3390/biology11070965
APA StyleTang, Z., Liu, S., & Niu, C. (2022). Effects of Extreme Light Cycle and Density on Melatonin, Appetite, and Energy Metabolism of the Soft-Shelled Turtle (Pelodiscus sinensis). Biology, 11(7), 965. https://doi.org/10.3390/biology11070965