
Phytoplasma Taxonomy: Nomenclature, Classification, and Identification


Abstract
:Simple Summary
Abstract
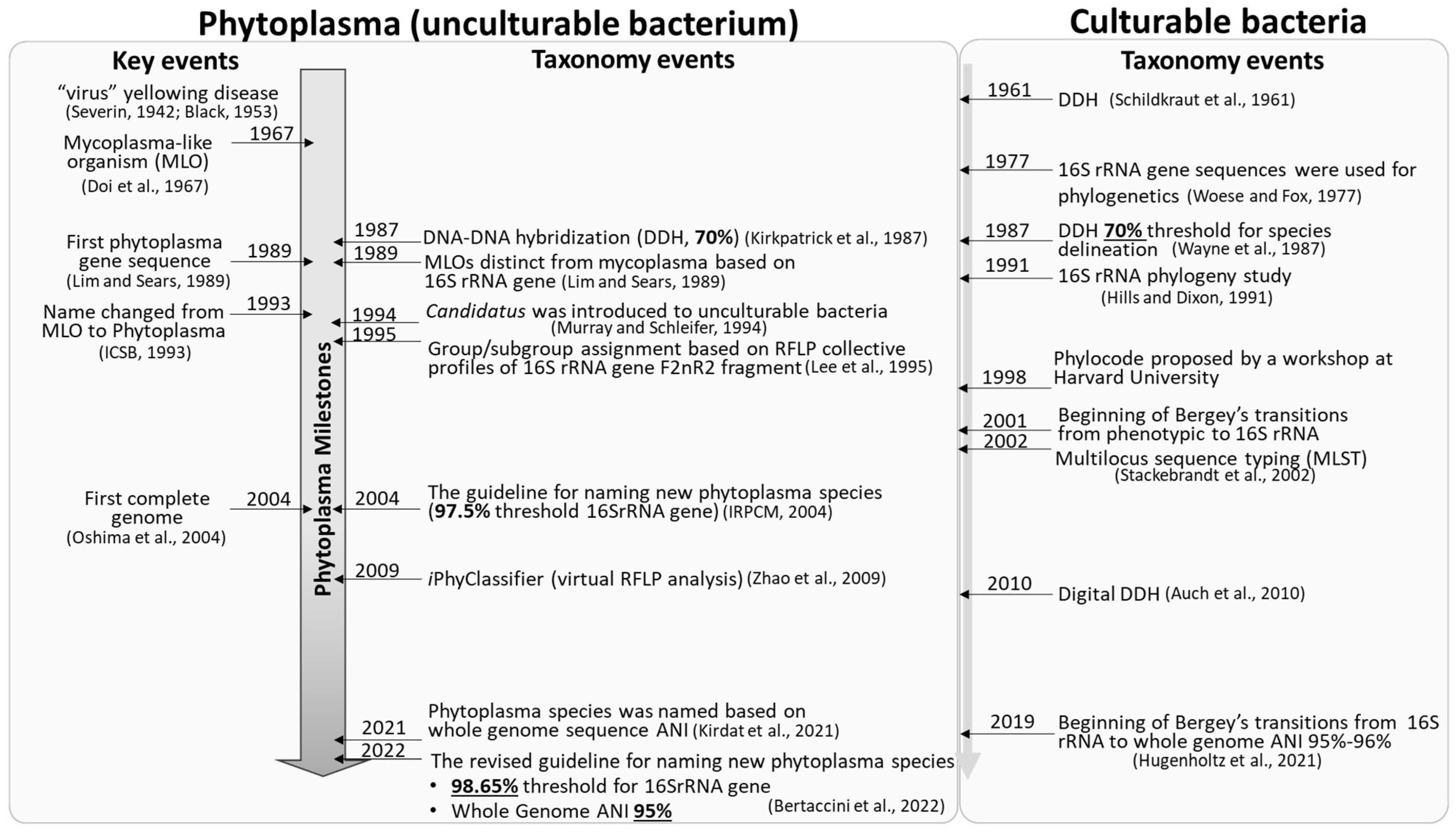
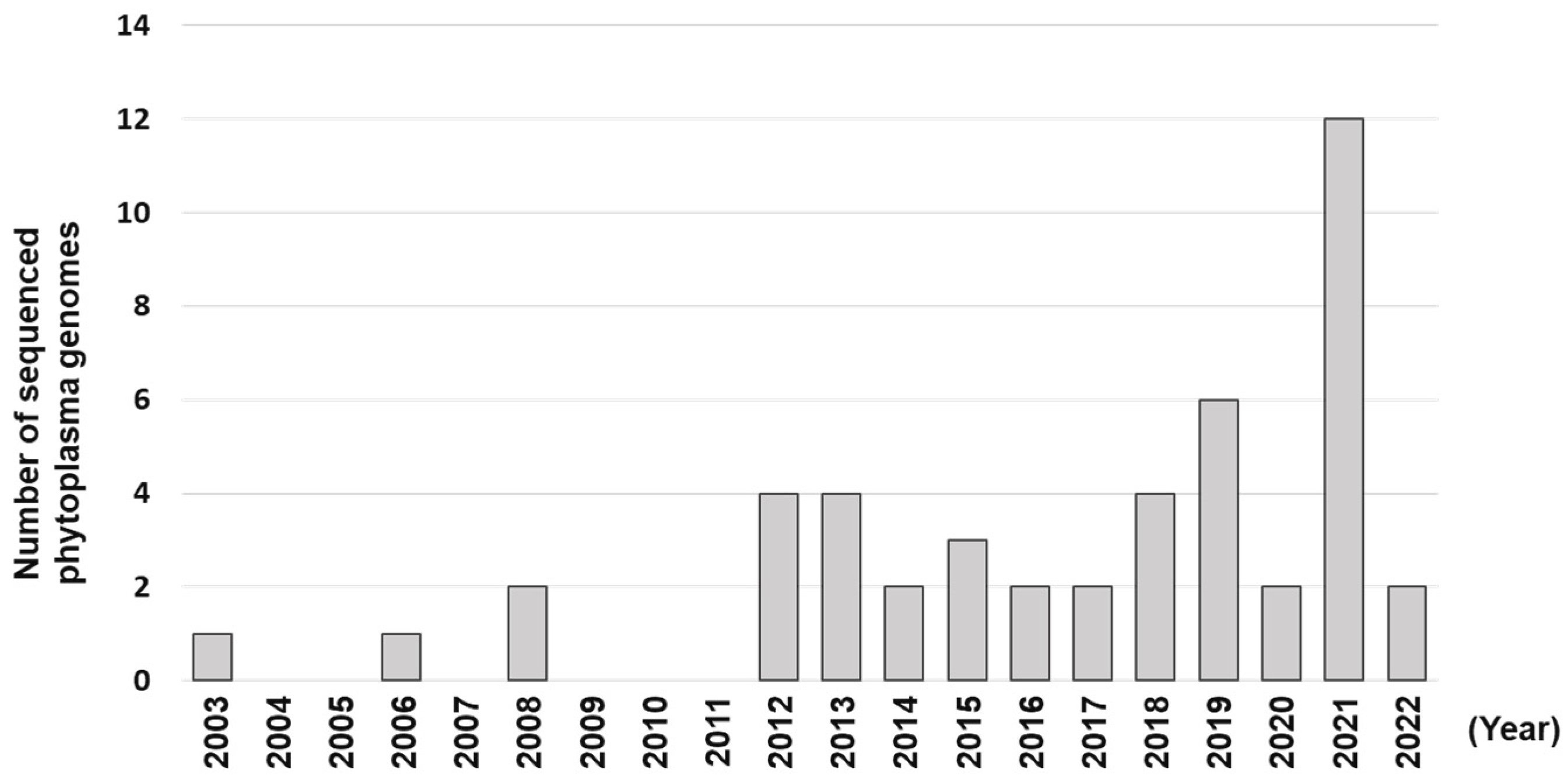
1. Introduction
2. Phytoplasma Nomenclature: Delineation of Candidatus Phytoplasma Species
2.1. Transition from 16S rRNA Gene to Whole Genome-Based Nomenclature of Candidatus Phytoplasma Species?
2.2. The Newly Revised 2022 Guidelines and Proposed Amendments
- (a)
- The ‘Ca. Phytoplasma’ species description should refer to a single, unique 16S rRNA gene sequence (full length or nearly full length, >1500 bp) or whole genome sequence with at least 60% coverage. The strain from which this sequence was obtained should be named the ‘reference strain’ and not the ‘type strain’. Strains in which even minimal differences in the 16S rRNA gene sequence from the reference strain are detected are referred as ‘related’ to the Candidatus species.
- (b)
- In general, a strain can be described as a novel ‘Ca. Phytoplasma’ species if its 16S rRNA gene sequence shares <98.65% identity or its whole genome shares an ANI score <95–96% to that of any previously described ‘Ca. Phytoplasma’ species.
- (c)
- There are, however, cases of phytoplasmas that share >98.65% identity of their 16S rRNA gene sequences or >95–96% ANI of their genomes, but clearly represent ecologically separated populations and, therefore, may deserve description as separate species. For such cases, description of two different species is recommended only when all three of the following conditions apply:
- (i)
- the two phytoplasmas are transmitted by different vectors;
- (ii)
- the two phytoplasmas have a different natural plant host (or, at least, their behavior is significantly different in the same plant host);
- (iii)
- there is evidence of significant molecular diversity, achieved by either hybridization to cloned DNA probes, serological reaction or multilocus sequence analysis (MLSA).
- (d)
- The rank of subspecies should not be used.
- (e)
- Due to strict international regulations, collection of micropropagation is no longer feasible or realistic; the gene clones of the reference strain should be deposited to the authorized scientific organizations in America, Europe, and Asia, etc. (to be determined by phytoplasma scientists).
- (f)
- Manuscripts that describe a novel ‘Ca. Phytoplasma’ species should preferably be submitted to the Int. J. Syst. Evol. Microbiol (IJSEM).
- (g)
- The abbreviation for Candidatus is Ca. (e.g., ‘Ca. Phytoplasma japonicum’ stands for ‘Candidatus Phytoplasma japonicum’).
3. Phytoplasma Classification: 16Sr Group/Subgroup Classification System Based on Collective RFLP Profiles
4. Phytoplasma Identification: Detection, Diagnostics and Characterization
5. Challenges and Perspectives
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Severin, H.H. Infection of perennial delphiniums by California-aster-yellows virus. Hilgardia 1942, 14, 411–440. [Google Scholar] [CrossRef] [Green Version]
- Black, L.M. Transmission of plant viruses by cicadellids. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 1953; Volume 1, pp. 69–89. [Google Scholar]
- Doi, Y.; Teranaka, M.; Yora, K.; Asuyama, H. Mycoplasma- or PLT group-like microorganisms found in the phloem elements of plants infected with mulberry dwarf, potato witches’ broom, aster yellows or paulownia witches’ broom. Ann. Phytopathol. Soc. Jpn. 1967, 33, 259–266. [Google Scholar] [CrossRef]
- Lim, P.O.; Sears, B.B. 16S rRNA sequence indicates that plant-pathogenic mycoplasmalike organisms are evolutionarily distinct from animal mycoplasmas. J. Bacteriol. 1989, 171, 5901–5906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Committee on Systematic Bacteriology Subcommittee on the Taxonomy of Mollicutes. Minutes of the Interim Meetings, 1 and 2 August, 1992, Ames, Iowa. Int. J. Syst. Bacteriol. 1993, 43, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Oshima, K.; Kakizawa, S.; Nishigawa, H.; Jung, H.Y.; Wei, W.; Suzuki, S.; Arashida, R.; Nakata, D.; Miyata, S.; Ugaki, M.; et al. Reductive evolution suggested from the complete genome sequence of a plant-pathogenic phytoplasma. Nat. Genet. 2004, 36, 27–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Zhang, J.; Ewing, A.; Miller, S.A.; Jancso Radek, A.; Shevchenko, D.V.; Tsukerman, K.; Walunas, T.; Lapidus, A.; Campbell, J.W.; et al. Living with genome instability: The adaptation of phytoplasmas to diverse environments of their insect and plant hosts. J. Bacteriol. 2006, 188, 3682–3696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Li, Q.; Zhao, Y.; Wei, H.; Wang, J.; Baker, C.J.; Liu, Q.; Wei, W. Integration of metabolomics and existing omics data reveals new insights into phytoplasma-induced metabolic reprogramming in host plants. PLoS ONE 2021, 16, e0246203. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.C.; Stenger, D.C.; Morris, T.J.; Purcell, A.H. Cloning and detection of DNA from a nonculturable plant pathogenic mycoplasma-like organism. Science 1987, 238, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.G.E.; Schleifer, K.H. Taxonomic notes: A proposal for recording the properties of putative taxa of procaryotes. Int. J. Syst. Evol. Microbiol. 1994, 44, 174–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.M.; Bertaccini, A.; Vibio, M.; Gundersen, D.E. Detection of multiple phytoplasmas in perennial fruit trees with decline symptoms in Italy. Phytopathology 1995, 85, 728–735. [Google Scholar] [CrossRef]
- IRPCM Phytoplasma/Spiroplasma Working Team–Phytoplasma Taxonomy Group. ‘Candidatus Phytoplasma’, a taxon for the wall-less, non-helical prokaryotes that colonize plant phloem and insects. Int. J. Syst. Evol. Microbiol. 2004, 54, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wei, W.; Lee, M.; Shao, J.; Suo, X.; Davis, R.E. Construction of an interactive online phytoplasma classification tool, iPhyClassifier, and its application in analysis of the peach X-disease phytoplasma group (16SrIII). Int. J. Syst. Evol. Microbiol. 2009, 59 Pt 10, 2582. [Google Scholar] [CrossRef] [PubMed]
- Kirdat, K.; Tiwarekar, B.; Thorat, V.; Sathe, S.; Shouche, Y.; Yadav, A. ‘Candidatus Phytoplasma sacchari’, a novel taxon-associated with Sugarcane Grassy Shoot (SCGS) disease. Int. J. Syst. Evol. Microbiol. 2021, 71, 004591. [Google Scholar] [CrossRef] [PubMed]
- Bertaccini, A.; Arocha-Rosete, Y.; Contaldo, N.; Duduk, B.; Fiore, N.; Montano, H.G.; Kube, M.; Kuo, C.H.; Martini, M.; Oshima, K.; et al. Revision of the ‘Candidatus Phytoplasma’species description guidelines. Int. J. Syst. Evol. Microbiol. 2022, 72, 005353. [Google Scholar] [CrossRef]
- Schildkraut, C.L.; Marmur, J.; Doty, P. The formation of hybrid DNA molecules and their use in studies of DNA homologies. J. Mol. Biol. 1961, 3, 595–617, IN15–IN16. [Google Scholar] [CrossRef]
- Woese, C.R.; Fox, G.E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef] [Green Version]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, L.H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.S.M.P.; et al. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Bacteriol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, P.; Chuvochina, M.; Oren, A.; Parks, D.H.; Soo, R.M. Prokaryotic taxonomy and nomenclature in the age of big sequence data. ISME J. 2021, 15, 1879–1892. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Frederiksen, W.; Garrity, G.M.; Grimont, P.A.; Kämpfer, P.; Maiden, M.C.; Nesme, X.; Rosselló-Mora, R.; Swings, J.; Trüper, H.G.; et al. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 2002, 52, 1043–1047. [Google Scholar]
- Auch, A.F.; von Jan, M.; Klenk, H.P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genomic. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, K.E.; Pettersson, B. Taxonomy of mollicutes. In Molecular Biology and Pathogenicity of Mycoplasmas (1–29); Springer: Boston, MA, USA, 2002. [Google Scholar]
- Baron, E.J. Classification. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 3. Available online: https://www.ncbi.nlm.nih.gov/books/NBK8406/ (accessed on 6 June 2022).
- Kämpfer, P.; Glaeser, S.P. Prokaryotic taxonomy in the sequencing era–the polyphasic approach revisited. Environ. Microbiol. 2012, 14, 291–317. [Google Scholar] [CrossRef] [PubMed]
- Rosselló-Mora, R.; Amann, R. The species concept for prokaryotes. FEMS Microbiol. Rev. 2001, 25, 39–67. [Google Scholar] [CrossRef]
- Chiykowski, L.N. Clover phyllody virus in Canada and its transmission. Can. J. Bot. 1962, 40, 397–404. [Google Scholar] [CrossRef]
- Freitag, J.H. Interaction and mutual suppression among three strains of aster yellows virus. Virology 1964, 24, 401–413. [Google Scholar] [CrossRef]
- Granados, R.R.; Chapman, R.K. Identification of some new aster yellows virus strains and their transmission by aster leafhopper Macrosteles fascifrons. Phytopathology 1968, 58, 1685. [Google Scholar]
- Chiykowski, L.N.; Sinha, R.C. Differentiation of MLO diseases by means of symptomatology and vector transmission. In Recent advances in mycoplasmology. In Proceedings of the 7th congress of the International Organization for Mycoplasmology, Baden near Vienna, Austria, 2–9 June 1988; Gustav Fischer Verlag: Baden near Vienna, Austria, 1990; pp. 280–287. [Google Scholar]
- McCoy, R.E.; Caudwell, A.; Chang, C.G.; Chen, T.A.; Chiykowski, L.N.; Cousin, M.T.; De Leeuw, G.D.; Golino, D.A.; Hacke, K.J.; Kirkpatrick, B.C.; et al. Mycoplasmalike organisms. Mycoplasmas 1989, 5, 545–568. [Google Scholar]
- Stackebrandt, E.; GOEBEL, B.M. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Stephen, J.R.; McCaig, A.E.; Smith, Z.; Prosser, J.I.; Embley, T.M. Molecular diversity of soil and marine 16S rRNA gene sequences related to beta-subgroup ammonia-oxidizing bacteria. Appl. Environ. Microbiol. 1996, 62, 4147–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drancourt, M.; Bollet, C.; Carlioz, A.; Martelin, R.; Gayral, J.P.; Raoult, D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 2000, 38, 3623–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, E.R.; Krüger, A.S.; Hauben, L.; Seal, S.E.; De Baere, R.; De Wachter, R.; Timmis, K.N.; Swings, J. 16S rRNA gene sequence analyses and inter-and intrageneric relationships of Xanthomonas species and Stenotrophomonas maltophilia. FEMS Microbiol. Lett. 1997, 151, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tse, H.; Yuen, K.Y. Then and now: Use of 16S rDNA gene sequencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Murray, R.G.E.; Sneath, P.H.A.; Mair, N.S.; Sharpe, M.E. Kingdom Procaryotae. Bergey’s Man. Syst. Bacteriol. 1984, 1, 34–36. [Google Scholar]
- Gasparich, G.E.; Bertaccini, A.; Zhao, Y. Candidatus Phytoplasma. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Oren, A. A plea for linguistic accuracy–also for Candidatus taxa. Int. J. Syst. Evol. Microbiol. 2017, 67, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Zreik, L.; Carle, P.; Bove, J.M.; Garnier, M. Characterization of the mycoplasmalike organism associated with witches’-broom disease of lime and proposition of a “Candidatus” taxon for the organism, “Candidatus Phytoplasma aurantifolia”. Int. J. Syst. Bacteriol. 1995, 45, 449–453. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Fox, G.E.; Wisotzkey, J.D.; Jurtshuk, P., Jr. How close is close: 16S rRNA sequence identity may not be sufficient to guarantee species identity. Int. J. Syst. Evol. Microbiol. 1992, 42, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Zhang, W. Comparative phylogenies of ribosomal proteins and the 16S rRNA gene at higher ranks of the class Actinobacteria. Curr. Microbiol. 2012, 65, 1–6. [Google Scholar] [CrossRef]
- Maiden, M.C. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 2006, 60, 561–588. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Wareing, D.R.A.; Ure, R.; Fox, A.J.; Bolton, F.E.; Bootsma, H.J.; Willems, R.J.L.; Urwin, R.; Maiden, M.C.J. Multilocus sequence typing system for Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 2, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kakizawa, S.; Makino, A.; Ishii, Y.; Tamaki, H.; Kamagata, Y. Draft genome sequence of “Candidatus Phytoplasma asteris” strain OY-V, an unculturable plant-pathogenic bacterium. Genome Announc. 2014, 2, e00944-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirdat, K.; Tiwarekar, B.; Thorat, V.; Narawade, N.; Dhotre, D.; Sathe, S.; Shouche, Y.; Yadav, A. Draft genome sequences of two phytoplasma strains associated with sugarcane grassy shoot (SCGS) and bermuda grass white leaf (BGWL) diseases. Mol. Plant-Microbe Interact 2020, 33, 715–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.H.; Cho, S.T.; Chen, C.L.; Yang, J.Y.; Kuo, C.H. Draft genome sequence of a 16SrII-A subgroup phytoplasma associated with purple coneflower (Echinacea purpurea) witches’ broom disease in Taiwan. Genome Announc. 2015, 3, e01398-15. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Quiroga, N.; Curkovic, S.T.; Zamorano, A.; Fiore, N. Detection and identification of 16SrXIII-F and a novel 16SrXIII phytoplasma subgroups associated with strawberry phyllody in Chile. Eur. J. Plant Pathol. 2019, 155, 1039–1046. [Google Scholar] [CrossRef]
- Huang, C.T.; Cho, S.T.; Lin, Y.C.; Tan, C.M.; Chiu, Y.C.; Yang, J.Y.; Kuo, C.H. Comparative Genome Analysis of ‘Candidatus Phytoplasma luffae’Reveals the Influential Roles of Potential Mobile Units in Phytoplasma Evolution. Front. Microbiol. 2022, 13, 773608. [Google Scholar] [CrossRef]
- Kube, M.; Schneider, B.; Kuhl, H.; Dandekar, T.; Heitmann, K.; Migdoll, A.M.; Reinhardt, R.; Seemüller, E. The linear chromosome of the plant-pathogenic mycoplasma ‘Candidatus Phytoplasma mali’. BMC Genet. 2008, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Fernández, F.D.; Conci, L.R. Genome characterization of ‘Candidatus Phytoplasma meliae’ (isolate ChTYXIII). bioRxiv 2021. [Google Scholar] [CrossRef]
- Fischer, A.; Santana-Cruz, I.; Wambua, L.; Olds, C.; Midega, C.; Dickinson, M.; Kawicha, P.; Khan, Z.; Masiga, D.; Jores, J.; et al. Draft genome sequence of “Candidatus Phytoplasma oryzae” strain Mbita1, the causative agent of Napier grass stunt disease in Kenya. Genome Announc. 2016, 4, e00297-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaglino, F.; Kube, M.; Jawhari, M.; Abou-Jawdah, Y.; Siewert, C.; Choueiri, E.; Sobh, H.; Casati, P.; Tedeschi, R.; Lova, M.M.; et al. ‘Candidatus Phytoplasma phoenicium’associated with almond witches’-broom disease: From draft genome to genetic diversity among strain populations. BMC Microbiol. 2015, 15, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Shao, J.; Zhao, Y.; Davis, R.E.; Costanzo, S. Draft genome sequence of ‘Candidatus Phytoplasma pini’-related strain MDPP: A resource for comparative genomics of gymnosperm-infecting phytoplasmas. Plant Dis. 2020, 104, 1009–1010. [Google Scholar] [CrossRef] [Green Version]
- Fernández, F.D.; Zübert, C.; Huettel, B.; Kube, M.; Conci, L.R. Draft Genome Sequence of “Candidatus Phytoplasma pruni” (X-disease group, subgroup 16SrIII-B) strain ChTDIII from Argentina. Microbiol. Resour. Announc. 2020, 9, e00792-20. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.M.; Shao, J.; Bottner-Parker, K.D.; Gundersen-Rindal, D.E.; Zhao, Y.; Davis, R.E. Draft genome sequence of “Candidatus Phytoplasma pruni” strain CX, a plant-pathogenic bacterium. Genome Announc. 2015, 3, e01117-15. [Google Scholar] [CrossRef] [Green Version]
- Music, M.S.; Samarzija, I.; Hogenhout, S.A.; Haryono, M.; Cho, S.T.; Kuo, C.H. The genome of ‘Candidatus Phytoplasma solani’strain SA-1 is highly dynamic and prone to adopting foreign sequences. Syst. Appl. Microbiol. 2019, 42, 117–127. [Google Scholar] [CrossRef]
- Mitrović, J.; Siewert, C.; Duduk, B.; Hecht, J.; Mölling, K.; Broecker, F.; Beyerlein, P.; Büttner, C.; Bertaccini, A.; Kube, M. Generation and Analysis of Draft Sequences of ‘Stolbur’Phytoplasma from Multiple Displacement Amplification Templates. Microb. Physiol. 2014, 24, 1–11. [Google Scholar] [CrossRef]
- Cai, W.; Nunziata, S.O.; Srivastava, S.K.; Wilson, T.; Chambers, N.; Rivera, Y.; Nakhla, M.; Costanzo, S. Draft Genome Sequence Resource of AldY-WA1, a Phytoplasma Strain Associated with Alder Yellows of Alnus rubra in Washington, U.S.A. Plant Dis. 2022, 106, 1971–1973. [Google Scholar] [CrossRef]
- Chen, W.; Li, Y.; Wang, Q.; Wang, N.; Wu, Y. Comparative genome analysis of wheat blue dwarf phytoplasma, an obligate pathogen that causes wheat blue dwarf disease in China. PLoS ONE 2014, 9, e96436. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Jiao, Q.; Yang, S.; Gao, R.; Lu, X.; Zhou, G. Comparative genome analysis of jujube witches’-broom Phytoplasma, an obligate pathogen that causes jujube witches’-broom disease. BMC Genom. 2018, 19, 689. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, D.; Galetto, L.; Rashidi, M.; Abbà, S.; Palmano, S.; Firrao, G.; Bosco, D.; Marzachì, C. Decreasing global transcript levels over time suggest that phytoplasma cells enter stationary phase during plant and insect colonization. Appl. Environ. Microbiol. 2015, 81, 2591–2602. [Google Scholar] [CrossRef] [Green Version]
- Nijo, T.; Iwabuchi, N.; Tokuda, R.; Suzuki, T.; Matsumoto, O.; Miyazaki, A.; Maejima, K.; Oshima, K.; Namba, S.; Yamaji, Y. Enrichment of phytoplasma genome DNA through a methyl-CpG binding domain-mediated method for efficient genome sequencing. J. Gen. Plant Pathol. 2021, 87, 154–163. [Google Scholar] [CrossRef]
- Saccardo, F.; Martini, M.; Palmano, S.; Ermacora, P.; Scortichini, M.; Loi, N.; Firrao, G. Genome drafts of four phytoplasma strains of the ribosomal group 16SrIII. Microbiology 2012, 158, 2805–2814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlovskis, Z.; Canale, M.C.; Haryono, M.; Lopes, J.R.S.; Kuo, C.H.; Hogenhout, S.A. A few sequence polymorphisms among isolates of Maize bushy stunt phytoplasma associate with organ proliferation symptoms of infected maize plants. Ann. Bot. 2017, 119, 869–884. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Zhang, X.; Meng, F.; Wang, Y.; Zhou, Y.; Liu, J. Draft Genome Sequences Resources of Mulberry Dwarf Phytoplasma Strain MDGZ-01 Associated with Mulberry Yellow Dwarf (MYD) Diseases. Plant Dis. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Sparks, M.E.; Bottner-Parker, K.D.; Gundersen-Rindal, D.E.; Lee, I.M. Draft genome sequence of the New Jersey aster yellows strain of ‘Candidatus Phytoplasma asteris’. PLoS ONE 2018, 13, e0192379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.T.; Lin, C.P.; Kuo, C.H. Genomic characterization of the periwinkle leaf yellowing (PLY) phytoplasmas in Tawan. Front. Microbiol. 2019, 10, 2194. [Google Scholar] [CrossRef] [PubMed]
- Town, J.R.; Wist, T.; Perez-Lopez, E.; Olivier, C.Y.; Dumonceaux, T.J. Genome sequence of a plant-pathogenic bacterium, “Candidatus Phytoplasma asteris” strain TW1. Microbiol. Resour. Announc. 2018, 7, e01109-18. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Sun, G.; Zhai, X.; Xu, P.; Ma, L.; Deng, M.; Zhao, Z.; Yang, H.; Dong, Y.; Shang, Z.; et al. Genomic insights into the fast growth of paulownias and the formation of Paulownia witches’ broom. Mol. Plant 2021, 14, 1668–1682. [Google Scholar] [CrossRef]
- Chung, W.C.; Chen, L.L.; Lo, W.S.; Lin, C.P.; Kuo, C.H. Comparative analysis of the peanut witches’-broom phytoplasma genome reveals horizontal transfer of potential mobile units and effectors. PLoS ONE 2013, 8, e62770. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; He, Y.; Zheng, Z.; Chen, J.; Wang, Z.; Zhou, G. Draft genome sequence of rice orange leaf phytoplasma from Guangdong, China. Genome Announc. 2017, 5, e00430-17. [Google Scholar] [CrossRef] [Green Version]
- Ranebennur, H.; Kirdat, K.; Tiwarekar, B.; Rawat, K.; Chalam, V.C.; Solanke, A.U.; Yadav, R.; Singh, K.; Sathe, S.; Yadav, A.; et al. Draft genome sequence of ‘Candidatus Phytoplasma australasia’, strain SS02 associated with sesame phyllody disease. 3 Biotech 2022, 12, 107. [Google Scholar] [CrossRef]
- Tran-Nguyen, L.T.; Kube, M.; Schneider, B.; Reinhardt, R.; Gibb, K.S. Comparative genome analysis of “Candidatus Phytoplasma australiense” (subgroup tuf-Australia I; rp-A) and “Ca. Phytoplasma asteris” strains OY-M and AY-WB. J. Bacteriol. 2008, 190, 3979–3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, M.T.; Liefting, L.W.; Havukkala, I.; Beever, R.E. Comparison of the complete genome sequence of two closely related isolates of ‘Candidatus Phytoplasma australiense’reveals genome plasticity. BMC Genom. 2013, 14, 529. [Google Scholar] [CrossRef] [Green Version]
- Debonneville, C.; Mandelli, L.; Brodard, J.; Groux, R.; Roquis, D.; Schumpp, O. The Complete Genome of the “Flavescence Dorée” Phytoplasma Reveals Characteristics of Low Genome Plasticity. Biology 2022, 11, 953. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Davis, R.E.; Lee, M.; Bottner-Parker, K.D. The agent associated with blue dwarf disease in wheat represents a new phytoplasma taxon, ‘Candidatus Phytoplasma tritici’. Int. J. Syst. Evol. Microbiol. 2021, 71, 004604. [Google Scholar] [CrossRef]
- Stackebrandt, E. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Kirkpatrick, B.C. Strategies for characterizing plant pathogenic mycoplasma-like organisms and their effects on plants. Plant-Microbe Interact. USA 1989. [Google Scholar]
- Davis, R.E.; Lee, I.; Dally, E.L.; Dewitt, N.; Douglas, S.M. Cloned nucleic acid hybridization probes in detection and classification of mycoplasmalike organisms (MLOs). In Proceedings of the VII International Symposium on Virus Diseases of Ornamental Plants, Sanremo, Italy, 29 May–2 June 1988; Volume 234, pp. 115–122. [Google Scholar]
- Zhao, Y.; Wei, W.; Davis, R.E.; Lee, I.-M. Recent advances in 16S rRNA gene-based phytoplasma differ-entiation, classification and taxonomy. In Phytoplasmas: Genomes, Plant Hosts and Vector; Weintraub, P., Jones, P., Eds.; CABI Publishing: Wallingford, UK, 2010; pp. 64–92. [Google Scholar]
- Lee, I.M.; Gundersen-Rindal, D.E.; Davis, R.E.; Bartoszyk, I.M. Revised classification scheme of phytoplasmas based on RFLP analyses of 16S rRNA and ribosomal protein gene sequences. Int. J. Syst. Evol. Microbiol. 2010, 48, 1153–1169. [Google Scholar] [CrossRef] [Green Version]
- Marcone, C.; Ragozzino, A.; Seemüller, E. Detection of Bermuda grass white leaf disease in Italy and characterization of the associated phytoplasma by RFLP analysis. Plant Dis. 1997, 81, 862–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-López, E.; Luna-Rodríguez, M.; Olivier, C.Y.; Dumonceaux, T.J. The underestimated diversity of phytoplasmas in Latin America. Int. J. Syst. Evol. Microbiol. 2016, 66, 492–513. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, F.D.; Meneguzzi, N.G.; Guzman, F.A.; Kirschbaum, D.S.; Conci, V.C.; Nome, C.F.; Conci, L.R. Detection and identification of a novel 16SrXIII subgroup phytoplasma associated with strawberry red leaf disease in Argentina. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 8, 2741–2747. [Google Scholar] [CrossRef] [Green Version]
- Nejat, N.; Vadamalai, G. Diagnostic techniques for detection of phytoplasma diseases: Past and present. J. Plant Dis. Prot. 2013, 120, 16–25. [Google Scholar] [CrossRef]
- Lee, I.-M.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Davis, R.E.; Lee, M.; Zhao, Y. Computer-simulated RFLP analysis of 16S rRNA genes: Identification of ten new phytoplasma groups. Int. J. Syst. Evol. Microbiol. 2007, 57, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Lee, M.; Davis, R.E.; Suo, X.; Zhao, Y. Automated RFLP pattern comparison and similarity coefficient calculation for rapid delineation of new and distinct phytoplasma 16Sr subgroup lineages. Int. J. Syst. Evol. Microbiol. 2008, 58, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Edel, V. Use of PCR and RFLP in fungal systematics. In Chemical Fungal Taxonomy; CRC Press: Boca Raton, FL, USA, 2020; pp. 51–76. [Google Scholar]
- Claussen, M.; Schmidt, S. Differentiation of Basidiobolus s isolates: RFLP of a diagnostic PCR amplicon matches sequence-based classification and growth temperature preferences. J. Fungi 2021, 7, 110. [Google Scholar] [CrossRef]
- Silvester, R.; Alexander, D.; Antony, A.C.; Hatha, M. GroEL PCR-RFLP–an efficient tool to discriminate closely related pathogenic Vibrio species. Microb. Pathog. 2017, 105, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.M.; Gundersen-Rindal, D.E.; Davis, R.E.; Bottner, K.D.; Marcone, C.; Seemüller, E. ‘Candidatus Phytoplasma asteris’, a novel phytoplasma taxon associated with aster yellows and related diseases. Int. J. Syst. Evol. Microbiol. 2004, 54, 1037–1048. [Google Scholar] [CrossRef] [Green Version]
- Arocha, Y.; Antesana, O.; Montellano, E.; Franco, P.; Plata, G.; Jones, P. ‘Candidatus Phytoplasma lycopersici’, a phytoplasma associated with ‘hoja de perejil’ disease in Bolivia. Int. J. Syst. Evol. Microbiol. 2007, 57, 1704–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, D.T.; Blackall, L.L.; Scott, P.T.; Walsh, K.B. Phylogenetic positions of phytoplasmas associated with dieback, yellow crinkle and mosaic diseases of papaya, and their proposed inclusion in ‘Candidatus Phytoplasma australiense’ and a new taxon, ‘Candidatus Phytoplasma australasia’. Int. J. Syst. Evol. Microbiol. 1998, 48, 941–951. [Google Scholar] [CrossRef]
- Davis, R.E.; Zhao, Y.; Dally, E.L.; Lee, M.; Jomantiene, R.; Douglas, S.M. ‘Candidatus Phytoplasma pruni’, a novel taxon associated with X-disease of stone fruits, Prunus s: Multilocus characterization based on 16S rRNA, secY, and ribosomal protein genes. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 2, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Martini, M.; Marcone, C.; Zhu, S.F. Classification of phytoplasma strains in the elm yellows group (16SrV) and proposal of ‘Candidatus Phytoplasma ulmi’ for the phytoplasma associated with elm yellows. Int. J. Syst. Evol. Microbiol. 2004, 54, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.Y.; Sawayanagi, T.; Kakizawa, S.; Nishigawa, H.; Wei, W.; Oshima, K.; Miyata, S.I.; Ugaki, M.; Hibi, T.; Namba, S. ‘Candidatus Phytoplasma ziziphi’, a novel phytoplasma taxon associated with jujube witches’-broom disease. Int. J. Syst. Evol. Microbiol. 2003, 53, 1037–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malembic-Maher, S.; Salar, P.; Filippin, L.; Carle, P.; Angelini, E.; Foissac, X. Genetic diversity of European phytoplasmas of the 16SrV taxonomic group and proposal of ‘Candidatus Phytoplasma rubi’. Int. J. Syst. Evol. Microbiol. 2011, 61, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Win, N.K.K.; Lee, S.Y.; Bertaccini, A.; Namba, S.; Jung, H.Y. ‘Candidatus Phytoplasma balanitae’ associated with witches’ broom disease of Balanites triflora. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 2, 636–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiruki, C.; Wang, K. Clover proliferation phytoplasma: ‘Candidatus Phytoplasma trifolii’. Int. J. Syst. Evol. Microbiol. 2004, 54, 1349–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, R.E.; Zhao, Y.; Dally, E.L.; Jomantiene, R.; Lee, M.; Wei, W.; Kitajima, E.W. ‘Candidatus Phytoplasma sudamericanum’, a novel taxon, and strain PassWB-Br4, a new subgroup 16SrIII-V phytoplasma, from diseased passion fruit (Passiflora edulis f. flavicarpa Deg.). Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 4, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.M.; Sinclair, W.A.; Smart, C.D.; Davis, R.E. The phytoplasma associated with ash yellows and lilac witches’-broom: ‘Candidatus Phytoplasma fraxini’. Int. J. Syst. Evol. Microbiol. 1999, 49, 1605–1614. [Google Scholar] [CrossRef]
- Davis, R.E.; Zhao, Y.; Wei, W.; Dally, E.L.; Lee, M. ‘Candidatus Phytoplasma luffae’, a novel taxon associated with witches’ broom disease of loofah, Luffa aegyptica Mill. Int. J. Syst. Evol. Microbiol. 1999, 67, 3127–3133. [Google Scholar] [CrossRef] [PubMed]
- Verdin, E.; Salar, P.; Danet, J.L.; Choueiri, E.; Jreijiri, F.; El Zammar, S.; Gelie, B.; Bove, J.M.; Garnier, M. ‘Candidatus Phytoplasma phoenicium’sp. nov., a novel phytoplasma associated with an emerging lethal disease of almond trees in Lebanon and Iran. Int. J. Syst. Evol. Microbiol. 2003, 53, 833–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemüller, E.; Schneider, B. ‘Candidatus Phytoplasma mali’, ‘Candidatus Phytoplasma pyri’and ‘Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline and European stone fruit yellows, respectively. Int. J. Syst. Evol. Microbiol. 2004, 54, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Marcone, C.; Gibb, K.S.; Streten, C.; Schneider, B. ‘Candidatus Phytoplasma spartii’, ‘Candidatus Phytoplasma rhamni’ and ‘Candidatus Phytoplasma allocasuarinae’, respectively associated with spartium witches’-broom, buckthorn witches’-broom and allocasuarina yellows diseases. Int. J. Syst. Evol. Microbiol. 2004, 54, 1025–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.Y.; Sawayanagi, T.; Wongkaew, P.; Kakizawa, S.; Nishigawa, H.; Wei, W.; Oshima, K.; Miyata, S.I.; Ugaki, M.; Hibi, T.; et al. ‘Candidatus Phytoplasma oryzae’, a novel phytoplasma taxon associated with wheat yellow dwarf disease. Int. J. Syst. Evol. Microbiol. 2003, 53, 1925–1929. [Google Scholar] [CrossRef]
- Šafárŏvá, D.; Zemanek, T.; Valova, P.; Navratil, M. ‘Candidatus Phytoplasma cirsii’, a novel taxon from creeping thistle [Cirsium arvense (L.) Scop.]. Int. J. Syst. Evol. Microbiol. 2016, 66, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.E.; Dally, E.L.; Gundersen, D.E.; Lee, I.-M.; Habili, N. “Candidatus Phytoplasma australiense,” a new phytoplasma taxon associated with Australian grapevine yellows. Int. J. Syst. Bacteriol. 1997, 47, 262–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawayanagi, T.; Horikoshi, N.; Kanehira, T.; Shinohara, M.; Bertaccini, A.; Cousin, M.T.; Hiruki, C.; Namba, S. ‘Candidatus Phytoplasma japonicum’, a new phytoplasma taxon associated with Japanese Hydrangea phyllody. Int. J. Syst. Evol. Microbiol. 1999, 49, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Valiunas, D.; Staniulis, J.; Davis, R.E. ‘Candidatus Phytoplasma fragariae’, a novel phytoplasma taxon discovered in yellows diseased strawberry, Fragaria× ananassa. Int. J. Syst. Evol. Microbiol. 1999, 56, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Quaglino, F.; Zhao, Y.; Casati, P.; Bulgari, D.; Bianco, P.A.; Wei, W.; Davis, R.E. ‘Candidatus Phytoplasma solani’, a novel taxon associated with stolbur-and bois noir-related diseases of plants. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 8, 2879–2894. [Google Scholar] [CrossRef] [Green Version]
- Martini, M.; Marcone, C.; Mitrović, J.; Maixner, M.; Delić, D.; Myrta, A.; Ermacora, P.; Bertaccini, A.; Duduk, B. ‘Candidatus Phytoplasma convolvuli’, a new phytoplasma taxon associated with bindweed yellows in four European countries. Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 12, 2910–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, R.E.; Harrison, N.A.; Zhao, Y.; Wei, W.; Dally, E.L. ‘Candidatus Phytoplasma hispanicum’, a novel taxon associated with Mexican periwinkle virescence disease of Catharanthus roseus. Int. J. Syst. Evol. Microbiol. 2016, 66, 3463–3467. [Google Scholar] [CrossRef]
- Fernández, F.D.; Galdeano, E.; Kornowski, M.V.; Arneodo, J.D.; Conci, L.R. Description of ‘Candidatus Phytoplasma meliae’, a phytoplasma associated with Chinaberry (Melia azedarach L.) yellowing in South America. Int. J. Syst. Evol. Microbiol. 2016, 66, 5244–5251. [Google Scholar] [CrossRef] [PubMed]
- Marcone, C.; Schneider, B.; Seemüller, E. ‘Candidatus Phytoplasma cynodontis’, the phytoplasma associated with Bermuda grass white leaf disease. Int. J. Syst. Evol. Microbiol. 2004, 54, 1077–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montano, H.G.; Davis, R.E.; Dally, E.L.; Hogenhout, S.; Pimentel, J.P.; Brioso, P.S. ‘Candidatus Phytoplasma brasiliense’, a new phytoplasma taxon associated with hibiscus witches’ broom disease. Int. J. Syst. Evol. Microbiol. 2001, 51, 1109–1118. [Google Scholar] [CrossRef] [Green Version]
- Arocha, Y.; Lopez, M.; Pinol, B.; Fernandez, M.; Picornell, B.; Almeida, R.; Palenzuela, I.; Wilson, M.R.; Jones, P. ‘Candidatus Phytoplasma graminis’ and ‘Candidatus Phytoplasma caricae’, two novel phytoplasmas associated with diseases of sugarcane, weeds and papaya in Cuba. Int. J. Syst. Evol. Microbiol. 2005, 55, 2451–2463. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Bottner, K.D.; Secor, G.; Rivera-Varas, V. ‘Candidatus Phytoplasma americanum’, a phytoplasma associated with a potato purple top wilt disease complex. Int. J. Syst. Evol. Microbiol. 2005, 56, 1593–1597. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.Y.; Sawayanagi, T.; Kakizawa, S.; Nishigawa, H.; Miyata, S.I.; Oshima, K.; Ugaki, M.; Lee, J.T.; Hibi, T.; Namba, S. ‘Candidatus Phytoplasma castaneae’, a novel phytoplasma taxon associated with chestnut witches’ broom disease. Int. J. Syst. Evol. Microbiol. 2002, 52, 1543–1549. [Google Scholar]
- Schneider, B.; Torres, E.; Martín, M.P.; Schröder, M.; Behnke, H.D.; Seemüller, E. ‘Candidatus Phytoplasma pini’, a novel taxon from Pinus silvestris and Pinus halepensis. Int. J. Syst. Evol. Microbiol. 2005, 55, 303–307. [Google Scholar] [CrossRef]
- Harrison, N.A.; Davis, R.E.; Oropeza, C.; Helmick, E.E.; Narvaez, M.; Eden-Green, S.; Dollet, M.; Dickinson, M. ‘Candidatus Phytoplasma palmicola’, associated with a lethal yellowing-type disease of coconut (Cocos nucifera L.) in Mozambique. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 6, 1890–1899. [Google Scholar] [CrossRef] [Green Version]
- Al-Saady, N.A.; Khan, A.J.; Calari, A.; Al-Subhi, A.M.; Bertaccini, A. ‘Candidatus Phytoplasma omanense’, associated with witches’-broom of Cassia italica (Mill.) Spreng. in Oman. Int. J. Syst. Evol. Microbiol. 2008, 58, 461–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Sun, Q.; Wei, W.; Davis, R.E.; Wu, W.; Liu, Q. ‘Candidatus Phytoplasma tamaricis’, a novel taxon discovered in witches’-broom-diseased salt cedar (Tamarix chinensis Lour.). Int. J. Syst. Evol. Microbiol. 2009, 59, 2496–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.M.; Bottner-Parker, K.D.; Zhao, Y.; Villalobos, W.; Moreira, L. ‘Candidatus Phytoplasma costaricanum’, a novel phytoplasma associated with an emerging disease in soybean (Glycine max). Int. J. Syst. Evol. Microbiol. 2011, 61, 2822–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejat, N.; Vadamalai, G.; Davis, R.E.; Harrison, N.A.; Sijam, K.; Dickinson, M.; Abdullah, S.N.A.; Zhao, Y. ‘Candidatus Phytoplasma malaysianum’, a novel taxon associated with virescence and phyllody of Madagascar periwinkle (Catharanthus roseus). Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 2, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Naderali, N.; Nejat, N.; Vadamalai, G.; Davis, R.E.; Wei, W.; Harrison, N.A.; Kong, L.; Kadir, J.; Tan, Y.H.; Zhao, Y. ‘Candidatus Phytoplasma wodyetiae’, a new taxon associated with yellow decline disease of foxtail palm (Wodyetia bifurcata) in Malaysia. Int. J. Syst. Evol. Microbiol. 2017, 67, 3765–3772. [Google Scholar] [CrossRef]
- Jardim, B.R.; Kinoti, W.M.; Tran-Nguyen, L.T.; Gambley, C.; Rodoni, B.; Constable, F.E. ‘Candidatus Phytoplasma stylosanthis’, a novel taxon with a diverse host range in Australia, characterised using multilocus sequence analysis of 16S rRNA, secA, tuf, and rp genes. Int. J. Syst. Evol. Microbiol. 2021, 71, ijsem004589. [Google Scholar]
- Miyazaki, A.; Shigaki, T.; Koinuma, H.; Iwabuchi, N.; Rauka, G.B.; Kembu, A.; Saul, J.; Watanabe, K.; Nijo, T.; Maejima, K.; et al. ‘Candidatus Phytoplasma noviguineense’, a novel taxon associated with Bogia coconut syndrome and banana wilt disease on the island of New Guinea. Int. J. Syst. Evol. Microbiol. 2018, 68, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.M.; Pease, B.; Perkins, S.L.; Constable, F.E.; Kinoti, W.M.; Warmington, D.; Allgood, B.; Powell, S.; Taylor, P.; Pearce, C.; et al. ‘Candidatus Phytoplasma dypsidis’, a novel taxon associated with a lethal wilt disease of palms in Australia. Int. J. Syst. Evol. Microbiol. 2021, 71, 004818. [Google Scholar] [CrossRef]
- Zhao, Y.; Davis, R.E. Criteria for phytoplasma 16Sr group/subgroup delineation and the need of a platform for proper registration of new groups and subgroups. Int. J. Syst. Evol. Microbiol. 2021, 66, 2121–2123. [Google Scholar] [CrossRef]
- MacLean, A.M.; Sugio, A.; Makarova, O.V.; Findlay, K.C.; Grieve, V.M.; Tóth, R.; Nicolaisen, M.; Hogenhout, S.A. Phytoplasma effector SAP54 induces indeterminate leaf-like flower development in Arabidopsis plants. Plant Physiol. 2011, 157, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Davis, R.E.; Nuss, D.L.; Zhao, Y. Phytoplasmal infection derails genetically preprogrammed meristem fate and alters plant architecture. Proc. Natl. Acad. Sci. USA 2013, 110, 19149–19154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Cai, H.; Wei, W.; Davis, R.E.; Lee, I.M.; Chen, H.; Zhao, Y. Identification of two new phylogenetically distant phytoplasmas from S enna surattensis plants exhibiting stem fasciation and shoot proliferation symptoms. Ann. Appl. Biol. 2012, 160, 25–34. [Google Scholar] [CrossRef]
- Zwolińska, A.; Krawczyk, K.; Borodynko-Filas, N.; Pospieszny, H. Non-crop sources of Rapeseed Phyllody phytoplasma (‘Candidatus Phytoplasma asteris’: 16SrI-B and 16SrI-(B/L)L), and closely related strains. Crop Prot. 2019, 119, 59–68. [Google Scholar] [CrossRef]
- Lee, I.M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasmalike organisms. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Gundersen, D.E.; Lee, I.-M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Mediterr. 1996, 35, 114–151. [Google Scholar]
- Christensen, N.M.; Nicolaisen, M.; Hansen, M.; Schulz, A. Distribution of phytoplasmas in infected plants as revealed by real-time PCR and bioimaging. Mol. Plant-Microbe Interact. 1993, 17, 1175–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Kakizawa, S.; Suzuki, S.; Jung, H.Y.; Nishigawa, H.; Miyata, S.; Oshima, K.; Ugaki, M.; Hibi, T.; Namba, S. In planta dynamic analysis of onion yellows phytoplasma using localized inoculation by insect transmission. Phytopathology 2004, 94, 244–250. [Google Scholar] [CrossRef] [Green Version]
- Mehle, N.; Dreo, T.; Ravnikar, M. Quantitative analysis of “flavescence doreé” phytoplasma with droplet digital PCR. Phytopathogenic Mollicutes 2014, 4, 9–15. [Google Scholar] [CrossRef]
- Dickinson, M. Loop-mediated isothermal amplification (LAMP) for detection of phytoplasmas in the field. In Plant Pathology; Humana Press: New York, NY, USA, 2015; pp. 99–111. [Google Scholar]
- Wheatley, M.S.; Wang, Q.; Wei, W.; Bottner-Parker, K.D.; Zhao, Y.; Yang, Y. Cas12a-based diagnostics for potato purple top disease complex associated with infection by ‘Candidatus Phytoplasma trifolii’-related strains. Plant Dis. 2022, PDIS09212119RE. [Google Scholar] [CrossRef]
- Deng, S.J.; Hiruki, C. Amplification of 16S ribosomal-RNA genes from culturable and nonculturable Mollicutes. J. Microbiol. Methods 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenic classification of plant pathogenic mycoplasmalike organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, I.R., Tully, J.G., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 369–380. [Google Scholar] [CrossRef]
- Casati, P.; Quaglino, F.; Stern, A.R.; Tedeschi, R.; Alma, A.; Bianco, P.A. Multiple gene analyses reveal extensive genetic diversity among ‘Candidatus Phytoplasma mali’populations. Ann. Appl. Biol. 2011, 158, 257–266. [Google Scholar] [CrossRef]
- Wei, W.; Cai, H.; Jiang, Y.; Lee, I.M.; Davis, R.E.; Ding, Y.; Yuan, E.; Chen, H.; Zhao, Y. A new phytoplasma associated with little leaf disease in azalea: Multilocus sequence characterization reveals a distinct lineage within the aster yellows phytoplasma group. Ann. Appl. Biol. 2011, 158, 318–330. [Google Scholar] [CrossRef]
- Varghese, N.J.; Mukherjee, S.; Ivanova, N.; Konstantinidis, K.T.; Mavrommatis, K.; Kyrpides, N.C.; Pati, A. Microbial species delineation using whole genome sequences. Nucleic Acids Res. 2015, 43, 6761–6771. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Organism Name | Organism Infraspecific Names Strain | 16Sr Group Classification | Host Symptoms | Country | References or GenBank Deposition | Assembly Accession | Assembly Stats Total Sequence Length | Assembly Level | Assembly Submission Date |
|---|---|---|---|---|---|---|---|---|---|
| ‘Catharanthus roseus’ aster yellows phytoplasma | De Villa | I-B | Maize Bushy Stunt-like | South Africa | Coetzee et al. deposited | GCF_004214875.1 | 603,949 | Complete Genome | 20 February 2019 |
| ‘Chrysanthemum coronarium’ phytoplasma | OY-V | I-B | onion yellows | Japan | [50] | GCF_000744065.1 | 739,592 | Contig | 14 August 2014 |
| ‘Cynodon dactylon’ phytoplasma | LW01 | XIV-A | Bermuda grass white leaf | India | [51] | GCF_009268075.1 | 483,935 | Scaffold | 22 October 2019 |
| ‘Echinacea purpurea’ witches’-broom phytoplasma | NCHU2014 | II-A | purple coneflower witches’ broom | Taiwan | [52] | GCF_001307505.1 | 545,427 | Contig | 7 October 2015 |
| ‘Fragaria x ananassa’ phyllody phytoplasma | StrPh-Cl | XIII-F | strawberry phyllody | Chile | [53] | GCF_018274325.1 | 627,584 | Contig | 4 May 2021 |
| ‘Parthenium hysterophorus’ phyllody phytoplasma | PR34 | II-new subgroup | Santa-Maria phyllody | India | Kirdat deposited | GCF_015100165.1 | 740,170 | Contig | 29 October 2020 |
| ‘Parthenium sp.’ Phyllody phytoplasma | PR08 | II-D | Santa-Maria phyllody | India | Kirdat deposited | GCF_015239935.1 | 586,816 | Contig | 10 May 2021 |
| ‘Santalum album’ aster yellows phytoplasma | SW86 | I-B | Sandalwood Spike | India | Tiwarekar deposited | GCF_018283495.1 | 554,025 | Contig | 5 May 2021 |
| Aster yellows witches’-broom phytoplasma AYWB | AYWB | I-A | Aster yellows witches’-broom in lettuce | USA | [7] | GCF_000012225.1 | 723,970 | Complete Genome | 1 November 2006 |
| Ca. Phytoplasma aurantifolia | WBDL | II-C | Lime witches’ broom phytoplasma in periwinkle | Oman | Foissac and Carle deposited | GCF_002009625.1 | 474,669 | Contig | 2 March 2017 |
| Ca. Phytoplasma luffae | NCHU2019 | VIII-A | Loofah witches’ broom | Taiwan | [54] | GCF_018024475.1 | 769,143 | Complete Genome | 16 April 2021 |
| Ca. Phytoplasma mali | AT | X-A | apple proliferation | NA | [55] | GCF_000026205.1 | 601,943 | Complete Genome | 4 July 2008 |
| Ca. Phytoplasma | ChTYXIII-Mo | XIII-G | Chinaberry yellowing | Argentina | [56] | GCF_016876135.2 | 751,949 | Contig | 14 April 2021 |
| Ca. Phytoplasma oryzae | NGS-S10 | XI-A | Napier Grass Stunt | Kenya | [57] | GCF_003263355.1 | 484,488 | Contig | 25 June 2018 |
| Ca. Phytoplasma phoenicium | SA213 | XI-D | almond witches’-broom | Lebanon | [58] | GCF_001189415.1 | 345,965 | Contig | 30 July 2015 |
| Ca. Phytoplasma pini | MDPP | XXI-B | pine phytoplasma | USA | [59] | GCF_007821455.1 | 474,136 | Contig | 1 August 2019 |
| Ca. Phytoplasma pruni | ChTDIII | III-B | China-tree decline | Argentina | [60] | GCF_013391955.1 | 790,517 | Contig | 8 July 2020 |
| Ca. Phytoplasma pruni | CX | III-A | Stone fruit tree decline | NA | [61] | GCF_001277135.1 | 598,511 | Contig | 1 September 2015 |
| Ca. Phytoplasma sacchari | SCGS | XI-B | Sugarcane Grassy Shoot | India | [51] | GCF_009268105.1 | 505,173 | Contig | 4 November 2019 |
| Ca. Phytoplasma solani | SA-1 | XII-A | Bois noir in Periwinkle | NA | [62] | GCF_003698095.1 | 821,322 | Contig | 30 October 2018 |
| Ca. Phytoplasma solani | 284/09 | XII-A | Stolbur phytoplasma (in tobacco and parsley) | NA | [63] | GCF_000970375.1 | 570,238 | Chromosome | 22 October 2013 |
| Ca. Phytoplasma sp. AldY-WA1 | AldY-WA1 | V-A | Alder yellows | USA | [64] | GCF_020312115.1 | 457,625 | Scaffold | 6 October 2021 |
| Ca. Phytoplasma tritici | WBD | I-C | 00420042pe blue dwarf | China | [65] | GCF_000495255.1 | 611,462 | Contig | 1 November 2013 |
| Ca. Phytoplasma ziziphi | Jwb-nky | V-B | jujube witches’ broom | China | [66] | GCF_003640545.1 | 750,803 | Complete Genome | 12 October 2018 |
| Chrysanthemum yellows phytoplasma | CYP | I-B | Chrysanthemum yellows | Italy | [67] | GCF_000803325.1 | 659,699 | Contig | 18 December 2014 |
| Hydrangea phyllody phytoplasma | HP | I-D | Hydrangea phyllody | Japan | [68] | GCF_018327665.1 | 597,775 | Contig | 28 April 2021 |
| Italian clover phyllody phytoplasma str. MA1 | MA1 | III-B | Italian clover phyllody (in periwinkle) | Italy | [69] | GCF_000300695.1 | 597,245 | Contig | 1 October 2012 |
| Maize bushy stunt phytoplasma | M3 | I-B | Maize bushy stunt | Brazil | [70] | GCF_001712875.1 | 576,118 | Complete Genome | 25 August 2016 |
| Milkweed yellows phytoplasma str. MW1 | MW1 | III-F | Milkweed yellows (in periwinkle) | Italy | [69] | GCF_000309485.1 | 583,806 | Contig | 1 October 2012 |
| Mulberry dwarf phytoplasma | MDGZ-01 | I-B | Mulberry dwarf | China | [71] | GCF_020714625.1 | 622,358 | Complete Genome | 2 November 2021 |
| New Jersey aster yellows phytoplasma | NJAY | I-A | New Jersey aster yellows (in periwinkle) | USA | [72] | GCA_002554195.1 | 652,092 | Contig | 16 October 2017 |
| Periwinkle leaf yellowing phytoplasma | DY2014 | I-B | Periwinkle leaf yellowing | Taiwan | [73] | GCA_005093185.1 | 824,596 | Contig | 2 May 2019 |
| ‘Brassica napus’ phytoplasma | TW1 | I-new subgroup | Rapeseed stunting and virescence | Canada | [74] | GCA_003181115.1 | 743,598 | Contig | 31 May 2018 |
| ‘Elaeagnus angustifolia’ witches’-broom phytoplasma | TBZ1 | I-new subgroup | Russian olive tree witches’-broom | Iran | Azizpour et al. deposited | GCA_018598675.1 | 833,199 | Contig | 30 May 2021 |
| Onion yellows phytoplasma | OY | I-B | Onion yellows (in chrysanthemum) | Japan | [6] | GCA_000009845.1 | 853,092 | Complete Genome | 9 December 2003 |
| Paulownia witches’-broom phytoplasma | Zhengzhou | I-D | Paulownia witches’-broom | China | [75] | GCF_019396865.1 | 891,641 | Complete Genome | 29 July 2021 |
| Peanut witches’-broom phytoplasma NTU2011 | NTU2011 | II-A | Peanut witches’-broom (in periwinkle) | Taiwan | [76] | GCF_000364425.1 | 566,694 | Contig | 26 March 2013 |
| Poinsettia branch-inducing phytoplasma str. JR1 | JR1 | III-H | Poinsettia branch-inducing (in periwinkle) | Italy | [69] | GCF_000309465.1 | 631,440 | Contig | 1 October 2012 |
| Rice orange leaf phytoplasma | LD1 | IX-A | Rice orange leaf | China | [77] | GCF_001866375.1 | 599,264 | Contig | 4 November 2016 |
| Sesame phyllody phytoplasma | SS02 | II-A or II-D | Sesame phyllody | India | [78] | GCF_018390775.1 | 536,153 | Contig | 17 May 2021 |
| Ca. Phytoplasma australiense | XII-B | Maintained in periwinkle | Australia | [79] | GCA_000069925.1 | 879,959 | Complete Genome | 2 April 2008 | |
| Strawberry lethal yellows phytoplasma (CPA) | NZSb11 | XII-B variant | Strawberry lethal yellows | Australia and New Zealand | [80] | GCF_000397185.1 | 959,779 | Complete Genome | 16 May 2013 |
| Texas Phoenix palm phytoplasma | Flo-TPPD | IV-D | Texas Phoenix Palm decline | USA | Bao et al. deposited | GCF_005774685.1 | 744,506 | Contig | 23 May 2019 |
| Vaccinium witches’-broom phytoplasma str. VAC | VAC | III-F | Vaccinium witches’-broom (in periwinkle) | Italy | [69] | GCF_000309405.1 | 647,754 | Contig | 1 October 2012 |
| Ca. Phytoplasma sp. | Tabriz.2 | I-B | Elaeagnus sp. (symptoms not described) | Iran | Zirak et al. deposited | GCA_019841745.1 | 762,261 | Contig | 24 August 2021 |
| Ca. Phytoplasma trifolii-related | CBPPT1 | VI-A | Potato purple top (in periwinkle) | USA | Wei et al. deposited | PRJNA839414 | 514,536 | Contig | 18 May 2022 |
| Florescence dorée (FD) phytoplasma | CH | V-A | Florescence dorée (in insect vector Scaphoideus titanus | Switzerland | [81] | PRJNA838420 | 654,223 | Complete Genome | 27 June 2022 |
| 2004 Guidelines (IRPCM [12]) | 2022 Revised Guidelines [15] | Suggested Clarification and Amendments to the 2022 Revised Guidelines |
|---|---|---|
| (a) The ‘Ca. Phytoplasma’ species description should refer to a single, unique 16S rRNA gene sequence (>1200 bp). The strain from which this sequence was obtained should be named the ‘reference strain’ and not the ‘type strain’. Strains in which even minimal differences in the 16S rRNA gene sequence from the reference strain are detected do not ‘belong’ to the Candidatus species, but are ‘related’ to it. | Extended the required length of 16S rRNA gene sequence from >1200 bp to full length or nearly full length. Introduced the term “member strains”. Comment: The new term may lead to erroneous assignment of a single given strain to more than one species. See Section 2.2 for details) | (a) The ‘Ca. Phytoplasma’ species description should refer to a single, unique 16S rRNA gene sequence (full length or nearly full length, >1500 bp) or whole genome sequence with at least 60% coverage (see Section 4 and Section 5). The strain from which this sequence was obtained should be named the ‘reference strain’ and not the ‘type strain’. Strains in which even minimal differences in the 16S rRNA gene sequence from the reference strain are detected are referred as ‘related’ to the Candidatus species. |
| (b) In general, a strain can be described as a novel ‘Ca. Phytoplasma’ species if its 16S rRNA gene sequence has <97.5% similarity to that of any previously described ‘Ca. Phytoplasma’ species. | Revised the threshold value for 16S rRNA gene sequence identity-based Ca. Phytoplasma species delineation to 98.65%. Proposed whole genome ANI-based criterum (95%) for Ca. Phytoplasma species delineation. | (b) In general, a strain can be described as a novel ‘Ca. Phytoplasma’ species if its 16S rRNA gene sequence shares <98.65% identity or its whole genome shares an ANI score <95–96% to that of any previously described ‘Ca. Phytoplasma’ species. |
| (c) There are, however, cases of phytoplasmas that share >97.5% of their 16S rRNA gene sequence, but clearly represent ecologically separated populations and, therefore, may deserve description as separate species. For such cases, description of two different species is recommended only when all three of the following conditions apply: (i) the two phytoplasmas are transmitted by different vectors; (ii) the two phytoplasmas have a different natural plant host (or, at least, their behaviour is significantly different in the same plant host); (iii) there is evidence of significant molecular diversity, achieved by either hybridization to cloned DNA probes, serological reaction or PCR-based assay. | If a strain shares >98.65% similarity in 16S rRNA gene sequence and >95% genome ANI with previously established species, two out of five housekeeping genes (groEL, tuf, rp, secA and secY) with suggested criteria can be used for delineating new species. Comment: All taxonomic frameworks attempt to reflect the ecological nature of organisms. Therefore, Rule (c) of the 2004 guidelines should be retained with necessary modifications. Due to the lack of universal primers and sufficient comparative analysis of housekeeping gene sequence data, it is still difficult to establish objective criteria for species delineation based on MLSA. But if the strain under study clearly represents ecologically separated populations, MLSA could be used to demonstrate significant molecular diversity in addition to fulfilling the unique vectorship and host specificity requirement (see Section 2.2 for details). | (c) There are, however, cases of phytoplasmas that share >98.65% identity in their 16S rRNA gene sequences or >95–96% ANI in their genomes, but clearly represent ecologically separated populations and, therefore, may deserve description as separate species. For such cases, description of two different species is recommended only when all three of the following conditions apply: (i) the two phytoplasmas are transmitted by different vectors; (ii) the two phytoplasmas have a different natural plant host (or, at least, their behaviour is significantly different in the same plant host); (iii) there is evidence of significant molecular diversity, achieved by either hybridization to cloned DNA probes, serological reaction or MLSA assay on at least two housekeeping genes. |
| (d) The rank of subspecies should not be used. | (d) The rank of subspecies should not be used. | |
| (e) The reference strain should be made available to the scientific community from the authors of the Candidatus species description paper and it should be deposited (unless in vitro micropropagation proves impossible) in the micropropagated collection of Dr Assunta Bertaccini, DiSTA, Patologia Vegetale, Universita‘ di Bologna, Italy. | (e) Due to strict international regulations, the rule e may no longer feasible; The gene clones of the reference strain should be deposited to the scientific committee or the authorized organizations in different countries in America, Europe, and Asia, etc (to be determined by phytoplasma scientists). | |
| (f) Manuscripts that describe a novel ‘Ca. Phytoplasma’ species should preferably be submitted to the Int. J. Syst. Evol. Microbiol (IJSEM). | (f) Manuscripts that describe a novel ‘Ca. Phytoplasma’ species should preferably be submitted to the Int. J. Syst. Evol. Microbiol (IJSEM). | |
| (g) The abbreviation for Candidatus is Ca. (e.g., ‘Ca. Phytoplasma japonicum’ stands for ‘Candidatus Phytoplasma japonicum’). | (g) The abbreviation for Candidatus is Ca. (e.g., ‘Ca. Phytoplasma japonicum’ stands for ‘Candidatus Phytoplasma japonicum’). |
| Group | Number of ‘Ca. Phytoplasma’ Species | Accession Number of Reference Strain | Subgroup | Reference | |
|---|---|---|---|---|---|
| 16SrI: Aster yellows group | 3 | ‘Ca. Phytoplasma asteris’ | M30790 | 16SI-B | [99] |
| ‘Ca. Phytoplasma lycopersici’ | EF199549 | 16SrI-Y | [100] | ||
| ‘Ca. Phytoplasma tritici’ | NZ AVAO01000003 | 16SrI-C | [82] | ||
| 16SrII: Peanut witches’ broom group | 1 | ‘Ca. Phytoplasma aurantifolia’ | U15442 | 16SrII-B | [41] |
| * Abolished | ‘Ca. Phytoplasma australasia’ | Y10096 | 16SrII-D | [101] | |
| 16SrIII: X-disease group | 1 | ‘Ca. Phytoplasma pruni’ | JQ044393 | 16SrIII-A | [102] |
| 16SrIV: Coconut lethal yellows group | 2 | ‘Ca. Phytoplasma palmae’ | U18747 | 16SrIV-A | [12,15] |
| ‘Ca. Phytoplasma cocostanzaniae’ | X80117 | 16SrIV-C | [12,15] | ||
| 16SrV: Elm yellows group | 4 | ‘Ca. Phytoplasma ulmi’ | AY197655 | 16SrV-A | [103] |
| ‘Ca. Phytoplasma ziziphi’ | AB052876 | 16SrV-B | [104] | ||
| ‘Ca. Phytoplasma rubi’ | AY197648 | 16SrV-E | [105] | ||
| ‘Ca. Phytoplasma balanitae’ | AB689678 | 16SrV-new subgroup | [106] | ||
| 16SrVI: Clover proliferation group | 2 | ‘Ca. Phytoplasma trifolii’ | AY390261 | 16SrVI-A | [107] |
| ‘Ca. Phytoplasma sudamericanum’ | GU292081 | 16SrVI-I | [108] | ||
| 16SrVII: Ash yellows group | 1 | ‘Ca. Phytoplasma fraxini’ | AF092209 | 16SrVII-A | [109] |
| 16SrVIII: Loofah witches’ broom group | 1 | ‘Ca. Phytoplasma luffae’ | AF248956 | 16SrVIII-A | [110] |
| 16SrIX: Pigeon pea witches’ broom group | 1 | ‘Ca. Phytoplasma phoenicium’ | AF248956 | 16SrIX-D | [111] |
| 16SrX: Apple proliferation group | 4 | ‘Ca. Phytoplasma mali’ | AJ542541 | 16SrX-A | [112] |
| ‘Ca. Phytoplasma pyri’ | AJ542543 | 16SrX-C | [112] | ||
| ‘Ca. Phytoplasma prunorum’ | AJ542544 | 16SrX-F | [112] | ||
| ‘Ca. Phytoplasma spartii’ | X92869 | 16SrX-D | [113] | ||
| 16SrXI: Rice yellow dwarf group | 3 | ‘Ca. Phytoplasma oryzae’ | AB052873 | 16SrXI-A | [114] |
| ‘Ca. Phytoplasma cirsii’ | KR869146 | 16SrXI-D | [115] | ||
| ‘Ca. Phytoplasma sacchari’ | VWXM00000000 | 16SrXI-B | [14] | ||
| 16SrXII: Stolbur group | 5 | ‘Ca. Phytoplasma australiense’ | L76865 | 16SrXII-B | [116] |
| ‘Ca. Phytoplasma japonicum’ | AB010425 | 16SrXII-D | [117] | ||
| ‘Ca. Phytoplasma fragariae’ | DQ086423 | 16SrXII-E | [118] | ||
| ‘Ca. Phytoplasma solani’ | AF248959 | 16SrXII-A | [119] | ||
| ‘Ca. Phytoplasma convolvuli’ | JN833705 | 16SrXII-H | [120] | ||
| 16SrXIII: Mexican periwinkle virescence group | 2 | ‘Ca. Phytoplasma hispanicum’ | AF248960 | 16SrXIII-A | [121] |
| ‘Ca. Phytcoplasma meliae | KU850940 | 16SrXIII-G | [122] | ||
| 16SrXIV: Bermudagrass white leaf group | 1 | ‘Ca. Phytoplasma cynodontis’ | AJ550984 | 16SrXIV-A | [123] |
| 16SrXV: Hibiscus witches’ broom group | 1 | ‘Ca. Phytoplasma brasiliense’ | AF147708 | 16SrXV-A | [124] |
| 16SrXVI: Sugar cane yellow leaf syndrome group | 1 | ‘Ca. Phytoplasma graminis’ | AY725228 | 16SrXVI-A | [125] |
| 16SrXVII: Papaya bunchy top group | 1 | ‘Ca. Phytoplasma caricae’ | AY725234 | 16SrXVII-A | [125] |
| 16SrXVIII: American potato purple top wilt group | 1 | ‘Ca. Phytoplasma americanum’ | DQ174122 | 16SrXVIII-A | [126] |
| 16SrXIX: Japanese chestnut witches’ broom group | 1 | ‘Ca. Phytoplasma castaneae’ | AB054986 | 16SrXIX-A | [127] |
| 16SrXX: Buckthorn witches’ broom group | 1 | ‘Ca. Phytoplasma rhamni’ | X76431 | 16SrXX-A | [113] |
| 16SrXXI: Pine shoot proliferation group | 1 | ‘Ca. Phytoplasma pini’ | AJ632155 | 16SrXXI-A | [128] |
| 16SrXXII: Nigerian coconut lethal decline group | 1 | ‘Ca. Phytoplasma palmicola’ | KF751387 | 16SrXXII-A | [129] |
| 16SrXXIII: Buckland Valley grapevine yellows group | 1 unnamed species identified | AY083605 | 16SrXXIII-A | [94] | |
| 16SrXXIV: Sorghum bunchy shoot group | 1 unnamed new species identified | AF509322 | 16SrXXIV-A | ||
| 16SrXXV: Weeping tea tree witches’ broom group | 1 unnamed new species identified | AF521672 | 16SrXXV-A | ||
| 16SrXXVI: Mauritius sugar cane yellows D3T1 group | 1 unnamed new species identified | AJ539179 | 16SrXXVI-A | ||
| 16SrXXVII: Mauritius sugar cane yellows D3T2 group | 1 unnamed new species identified | AJ539180 | 16SrXXVII-A | ||
| 16SrXXVIII: Havana derbid group | 1 unnamed new species identified | AY744945 | 16SrXXVII-A | ||
| 16SrXXIX: Cassia witches’ broom group | 1 | ‘Ca. Phytoplasma omanense’ | EF666051 | 16SrXXIX-A | [130] |
| 16SrXXX: Salt cedar witches’ broom group | 1 | ‘Ca. Phytoplasma tamaricis’ | FJ432664 | 16SrXXX-A | [131] |
| 16SrXXXI: Soybean stunt phytoplasma group | 1 | ‘Ca. Phytoplasma costaricanum’ | HQ225630 | 16SrXXXI-A | [132] |
| 16SrXXXII: Malaysian periwinkle virescence group | 1 | ‘Ca. Phytoplasma malaysianum’ | EU371934 | 16SrXXXII-A | [133] |
| 16SrXXXIII: Allocasuarina group | 1 | ‘Ca. Phytoplasma allocasuarinae’ | AY135523 | 16SrXXXIII-A | [12] |
| 16SrXXXIV: grapevine yellows | No new species identified, abolished | DQ232752 | |||
| 16SrXXXV: Pepper witches’-broom | No new species identified, abolished | EU125184 | |||
| 16SrXXXVI: foxtail palm yellow decline group | 1 | ‘Ca. Phytoplasma wodyetiae’ | KC844879 | 16SrXXXVI-A | [134] |
| 16SrXXXVII: Stylosanthes little leaf group | 1 | ‘Ca. Phytoplasma stylosanthis’ | MT431550 | 16SrXXXVII-A | [135] |
| 16SrXXXVIII: Bogia coconut syndrome group | 1 | ‘Ca. Phytoplasma noviguineense’ | LC228755 | 16SrXXXVIII-A | [136] |
| 16SrXXXIX: Palm lethal wilt group | 1 | ‘Ca. Phytoplasma dypsidis’ | MT536195 | 16SrXXXIX-A | [137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, W.; Zhao, Y. Phytoplasma Taxonomy: Nomenclature, Classification, and Identification. Biology 2022, 11, 1119. https://doi.org/10.3390/biology11081119
Wei W, Zhao Y. Phytoplasma Taxonomy: Nomenclature, Classification, and Identification. Biology. 2022; 11(8):1119. https://doi.org/10.3390/biology11081119
Chicago/Turabian StyleWei, Wei, and Yan Zhao. 2022. "Phytoplasma Taxonomy: Nomenclature, Classification, and Identification" Biology 11, no. 8: 1119. https://doi.org/10.3390/biology11081119
APA StyleWei, W., & Zhao, Y. (2022). Phytoplasma Taxonomy: Nomenclature, Classification, and Identification. Biology, 11(8), 1119. https://doi.org/10.3390/biology11081119