
Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants

 ,
, 
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Isolation, Identification, and Molecular Characterization
2.2. Bacterial Isolation, Characterization, and Molecular Identification
2.3. Greenhouse Experimental Design
2.4. Evaluation of the Total Soluble Carbohydrates and Proteins
2.5. Evaluation of Ascorbic Acid Content
2.6. Free Radical Scavenging Activity Evaluation
2.7. Antioxidant Enzyme Assays
2.8. The Effect of Foliar CF Application on Squash Plant Gene Expression under ZYMV Challenge
2.9. Identification of CF Bioactive Compounds through Gas Chromatography–Mass Spectrometry (GC–MS)
2.10. Statistical Analysis
3. Results and Discussion
3.1. Virus Isolation and Identification
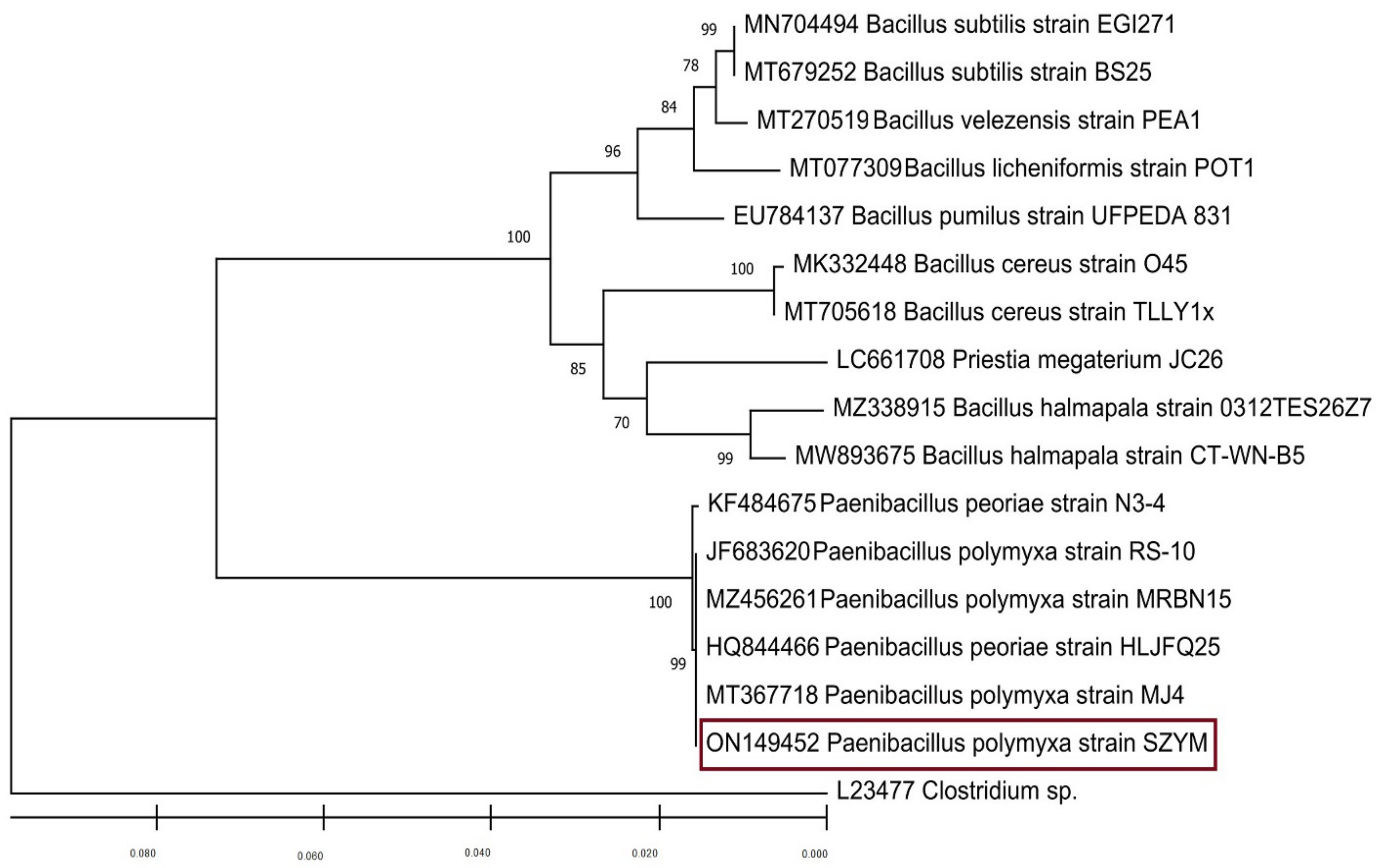
3.2. Bacterial Isolation and Identification
3.3. Efficacy of SZYM-CF on Symptom Development, Growth Parameters, and Viral Accumulation Level
3.4. Effect of SZYM-CF on Total Soluble Carbohydrates and Total Soluble Protein Content
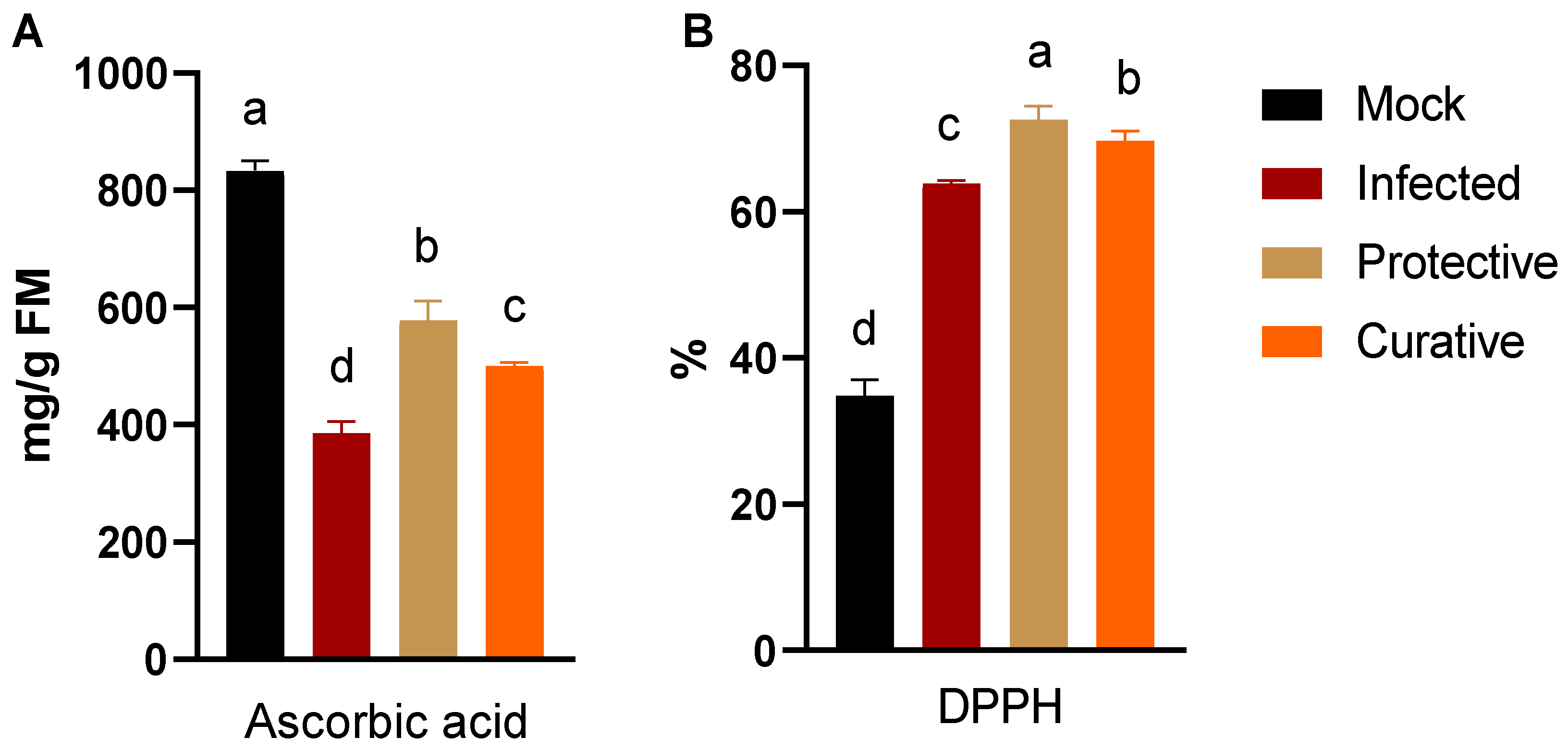
3.5. Alternation in Ascorbic acid Content and Free Radical Scavenging Activity
3.6. Antioxidant Enzymes Evaluation
3.7. Effect of SZYM-CF Foliar Application on Polyphenolic Pathway Synthesis Genesʹ Expression
3.8. Effect of Foliar Application of SZYM-CF on Pathogenesis-Related Genesʹ Expression

3.9. Phytochemical Analysis of the SZYM-CF Using GC–MS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Novello, G.; Cesaro, P.; Bona, E.; Massa, N.; Gosetti, F.; Scarafoni, A.; Todeschini, V.; Berta, G.; Lingua, G.; Gamalero, E. The effects of plant growth-promoting bacteria with biostimulant features on the growth of a local onion cultivar and a commercial zucchini variety. Agronomy 2021, 11, 888. [Google Scholar] [CrossRef]
- Al-Kubati, A.M.S.; Kang, B.; Liu, L.; Abbas, A.; Gu, Q. Development of bottle gourd lines resistant to zucchini yellow mosaic virus using ethyl methanesulfonate mutagenesis. HortScience 2021, 56, 909–914. [Google Scholar] [CrossRef]
- Fletcher, J.D.; Wallace, A.R.; Rogers, B.T. Potyviruses in New Zealand buttercup squash (Cucurbits maxima Duch.): Yield and quality effects of ZYMV and WMV 2 virus infections. N. Z. J. Crop Hortic. Sci. 2000, 28, 17–26. [Google Scholar] [CrossRef]
- Coutts, B.A.; Kehoe, M.A.; Jones, R.A.C. Minimising losses caused by Zucchini yellow mosaic virus in vegetable cucurbit crops in tropical, sub-tropical and Mediterranean environments through cultural methods and host resistance. Virus Res. 2011, 159, 141–160. [Google Scholar] [CrossRef]
- Bubici, G.; Navarro, B.; Carluccio, A.V.; Ciuffo, M.; Di Serio, F.; Cillo, F. Genomic sequence variability of an Italian Zucchini yellow mosaic virus isolate. Eur. J. Plant Pathol. 2020, 156, 325–332. [Google Scholar] [CrossRef]
- Coutts, B.A.; Kehoe, M.A.; Webster, C.G.; Wylie, S.J.; Jones, R.A.C. Zucchini yellow mosaic virus: Biological properties, detection procedures and comparison of coat protein gene sequences. Arch. Virol. 2011, 156, 2119–2131. [Google Scholar] [CrossRef]
- Maina, S.; Coutts, B.A.; Edwards, O.R.; De Almeida, L.; Kehoe, M.A.; Ximenes, A.; Jones, R.A.C. Zucchini yellow mosaic virus populations from East Timorese and Northern Australian cucurbit crops: Molecular properties, genetic connectivity, and biosecurity implications. Plant Dis. 2017, 101, 1236–1245. [Google Scholar] [CrossRef] [Green Version]
- Shongwe, L.T.; Masarirambi, M.T.; Oseni, T.O.; Wahome, P.K.; Nxumalo, K.A.; Gule, P.I. Effects of hydroponics systems on growth, yield and quality of zucchini (Cucurbita pepo L.). J. Plant Stud. 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Thanh, N.C.; Eed, E.M.; Elfasakhany, A.; Brindhadevi, K. Antioxidant, anti-inflammatory and anti-proliferative activities of green and yellow zucchini (Courgette). Appl. Nanosci. 2022, 3, 1–10. [Google Scholar] [CrossRef]
- Martínez-Valdivieso, D.; Font, R.; Fernández-Bedmar, Z.; Merinas-Amo, T.; Gómez, P.; Alonso-Moraga, Á.; Río-Celestino, D. Role of zucchini and its distinctive components in the modulation of degenerative processes: Genotoxicity, anti-genotoxicity, cytotoxicity and apoptotic effects. Nutrients 2017, 9, 755. [Google Scholar] [CrossRef] [Green Version]
- Bhan, C.; Kannaujia, P.K.; Shrivastava, A.K.; Bairwa, S.K.; Duhan, S.; Prasad, H.; Prasad, D. Antidiabetic potential of bioactive extracts from fruits and vegetables: A review. J. Progress. Agric. 2015, 6, 44–47. [Google Scholar]
- De Lima Souza, M.W.; Torres, S.B.; De Assis de Oliveira, F.; Da Silva Marques, I.C.; Pereira, K.T.O.; Guimarães, Í.T. Saline-water irrigation and plant growth regulator application on zucchini fruit yield and quality. Rev. Bras. Eng. Agríc. Ambient. 2020, 24, 679–684. [Google Scholar] [CrossRef]
- Formisano, L.; Miras-Moreno, B.; Ciriello, M.; El-Nakhel, C.; Corrado, G.; Lucini, L.; Colla, G.; Rouphael, Y. Trichoderma and phosphite elicited distinctive secondary metabolite signatures in zucchini squash plants. Agronomy 2021, 11, 1205. [Google Scholar] [CrossRef]
- Gal-On, A. A point mutation in the FRNK motif of the potyvirus helper component-protease gene alters symptom expression in cucurbits and elicits protection against the severe homologous virus. Phytopathology 2000, 90, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Kang, D.; Shi, Z.; Shen, H.; Wehner, T. Inheritance of resistance to zucchini yellow mosaic virus and watermelon mosaic virus in watermelon. J. Hered. 2004, 95, 498–502. [Google Scholar] [CrossRef]
- Essalimi, B.; Esserti, S.; Rifai, L.A.; Koussa, T.; Makroum, K.; Belfaiza, M.; Rifai, S.; Venisse, J.S.; Faize, L.; Alburquerque, N. Enhancement of plant growth, acclimatization, salt stress tolerance and verticillium wilt disease resistance using plant growth-promoting rhizobacteria (PGPR) associated with plum trees (Prunus domestica). Sci. Hortic. 2022, 291, 110621. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S.; El-Beltagi, H.S.; Umar, S.; Lee, J. Bioprospecting Plant Growth Promoting Rhizobacteria for Enhancing the Biological Properties and Phytochemical Composition of Medicinally Important Crops. Molecules 2022, 27, 1407. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef]
- Zhao, S.; Li, Y. Current understanding of the interplays between host hormones and plant viral infections. PLoS Pathog. 2021, 17, e1009242. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef] [PubMed]
- El-Gendi, H.; Al-Askar, A.A.; Király, L.; Samy, M.A.; Moawad, H.; Abdelkhalek, A. Foliar Applications of Bacillus subtilis HA1 Culture Filtrate Enhance Tomato Growth and Induce Systemic Resistance against Tobacco mosaic virus Infection. Horticulturae 2022, 8, 301. [Google Scholar] [CrossRef]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Antonie Van Leeuwenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Zhu, J.; Tan, T.; Xu, J.; Shen, A.; Yang, X.; Li, J.; Zeng, L.; Wei, L. Isolation and characterization of antagonistic Paenibacillus polymyxa HX-140 and its biocontrol potential against Fusarium wilt of cucumber seedlings. BMC Microbiol. 2021, 21, 75. [Google Scholar] [CrossRef]
- Chávez-Ramírez, B.; Rodríguez-Velázquez, N.D.; Mondragón-Talonia, C.M.; Avendaño-Arrazate, C.H.; Martínez-Bolaños, M.; Vásquez-Murrieta, M.S.; Estrada de los Santos, P. Paenibacillus polymyxa NMA1017 as a potential biocontrol agent of Phytophthora tropicalis, causal agent of cacao black pod rot in Chiapas, Mexico. Antonie Van Leeuwenhoek 2021, 114, 55–68. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Mahgoub, H.A.; Desbiez, C.; Wipf-Scheibel, C.; Dafalla, G.; Lecoq, H. Characterization and occurrence of zucchini yellow mosaic virus in Sudan. Plant Pathol. 1997, 46, 800–805. [Google Scholar] [CrossRef]
- Heflish, A.A.; Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Protective and Curative Effects of Trichoderma asperelloides Ta41 on Tomato Root Rot Caused by Rhizoctonia solani Rs33. Agronomy 2021, 11, 1162. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Salem, M.Z.M.; Hafez, E.; Behiry, S.I.; Qari, S.H. The Phytochemical, Antifungal, and First Report of the Antiviral Properties of Egyptian Haplophyllum tuberculatum Extract. Biology 2020, 9, 248. [Google Scholar] [CrossRef]
- Sambrook, J. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hafez, E.E.; El-Morsi, A.A.; El-Shahaby, O.A.; Abdelkhalek, A.A. Occurrence of iris yellow spot virus from onion crops in Egypt. VirusDisease 2014, 25, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; El-Gendi, H.; Al-Askar, A.A.; Maresca, V.; Moawad, H.; Elsharkawy, M.M.; Younes, H.A.; Behiry, S.I. Enhancing systemic resistance in faba bean (Vicia faba L.) to Bean yellow mosaic virus via soil application and foliar spray of nitrogen-fixing Rhizobium leguminosarum bv. viciae strain 33504-Alex1. Front. Plant Sci. 2022, 2679. [Google Scholar] [CrossRef]
- Islam, M.J.; Kim, J.W.; Begum, M.K.; Sohel, M.A.; Lim, Y.-S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants 2020, 9, 1511. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Oser, B.L.; Hawk, P.B. Hawk’s Physiological Chemistry, 14th ed.; McGraw-Hill: New York, NY, USA, 1965. [Google Scholar]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Cho, Y.K.; Ahn, H.Y.E.K. Purification and characterization of polyphenol oxidase from potato: II. Inhibition and catalytic mechanism. J. Food Biochem. 1999, 23, 593–605. [Google Scholar] [CrossRef]
- Angelini, R.; Manes, F.; Federico, R. Spatial and functional correlation between diamine-oxidase and peroxidase activities and their dependence upon de-etiolation and wounding in chick-pea stems. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef]
- Behiry, S.I.; Ashmawy, N.A.; Abdelkhalek, A.A.; Younes, H.A.; Khaled, A.E.; Hafez, E.E. Compatible- and incompatible-type interactions related to defense genes in potato elucidation by Pectobacterium carotovorum. J. Plant Dis. Prot. 2018, 125, 197–204. [Google Scholar] [CrossRef]
- ElMorsi, A.; Abdelkhalek, A.; Alshehaby, O.; Hafez, E.E. Pathogenesis-related genes as tools for discovering the response of onion defence system against iris yellow spot virus infection. Botany 2015, 93, 735–744. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Alsubaie, M.M.; Behiry, S.I. First Report of Protective Activity of Paronychia argentea Extract against Tobacco Mosaic Virus Infection. Plants 2021, 10, 2435. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kheder, A.; Sulaiman, T.; Ghanem, G.; Tohamy, M. Biological, serological and molecular characterization of Egyptian Zucchini yellow mosaic virus isolate infecting squash plants in Fayoum Governorate. Egypt. J. Phytopathol. 2017, 45, 1–14. [Google Scholar] [CrossRef]
- Derbalah, A.; Abdelsalam, I.; Behiry, S.; Abdelkhalek, A.; Abdelfatah, M.; Ismail, S.; Elsharkawy, M.M. Copper oxide nanostructures as a potential method for control zucchini yellow mosaic virus in Squash. Pest Manag. Sci. 2022, 78, 3587–3595. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef]
- Li, H.P.; Gan, Y.N.; Yue, L.J.; Han, Q.Q.; Chen, J.; Liu, Q.M.; Zhao, Q.; Zhang, J.L. Newly Isolated Paenibacillus monticola sp. nov., a Novel Plant Growth-Promoting Rhizobacteria Strain From High-Altitude Spruce Forests in the Qilian Mountains, China. Front. Microbiol. 2022, 13, 833313. [Google Scholar] [CrossRef]
- Yaoyao, E.; Yuan, J.; Yang, F.; Wang, L.; Ma, J.; Li, J.; Pu, X.; Raza, W.; Huang, Q.; Shen, Q. PGPR strain Paenibacillus polymyxa SQR-21 potentially benefits watermelon growth by re-shaping root protein expression. AMB Express 2017, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Langendries, S.; Goormachtig, S. Paenibacillus polymyxa, a Jack of all trades. Environ. Microbiol. 2021, 23, 5659–5669. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Hamad, A.; Lu, G. Physiological and metabolic changes of Cucurbita pepo leaves in response to zucchini yellow mosaic virus (ZYMV) infection and salicylic acid treatments. Plant Physiol. Biochem. PPB 2007, 45, 480–489. [Google Scholar] [CrossRef]
- Gallie, D.R. L-ascorbic Acid: A multifunctional molecule supporting plant growth and development. Scientifica 2013, 2013, 795964. [Google Scholar] [CrossRef] [Green Version]
- Sofy, A.R.; Dawoud, R.A.; Sofy, M.R.; Mohamed, H.I.; Hmed, A.A.; El-Dougdoug, N.K. Improving Regulation of Enzymatic and Non-Enzymatic Antioxidants and Stress-Related Gene Stimulation in Cucumber mosaic cucumovirus-Infected Cucumber Plants Treated with Glycine Betaine, Chitosan and Combination. Molecules 2020, 25, 2341. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613–630. [Google Scholar] [CrossRef]
- Fujiwara, A.; Shimura, H.; Masuta, C.; Sano, S.; Inukai, T. Exogenous ascorbic acid derivatives and dehydroascorbic acid are effective antiviral agents against Turnip mosaic virus in Brassica rapa. J. Gen. Plant Pathol. 2013, 79, 198–204. [Google Scholar] [CrossRef]
- Wang, S.D.; Zhu, F.; Yuan, S.; Yang, H.; Xu, F.; Shang, J.; Xu, M.Y.; Jia, S.D.; Zhang, Z.W.; Wang, J.H.; et al. The roles of ascorbic acid and glutathione in symptom alleviation to SA-deficient plants infected with RNA viruses. Planta 2011, 234, 171–181. [Google Scholar] [CrossRef]
- Fujiwara, A.; Togawa, S.; Hikawa, T.; Matsuura, H.; Masuta, C.; Inukai, T. Ascorbic acid accumulates as a defense response to Turnip mosaic virus in resistant Brassica rapa cultivars. J. Exp. Bot. 2016, 67, 4391–4402. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Zhang, Q.P.; Che, Y.P.; Zhu, P.X.; Zhang, Q.Q.; Ji, Z.L. Glutathione contributes to resistance responses to TMV through a differential modulation of salicylic acid and reactive oxygen species. Mol. Plant Pathol. 2021, 22, 1668–1687. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Alici, E.H.; Arabaci, G. Determination of SOD, POD, PPO and CAT Enzyme Activities in Rumex obtusifolius L. Annu. Res. Rev. Biol. 2016, 11, 1–7. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Anuradha, C.; Selvarajan, R.; Vasantha, S.; Suresha, G.S. Biochemical Characterization of Compatible Plant Virus Interaction: A Case Study with Bunchy Top Virus-Banana Host-Pathosystem. Plant Pathol. J. 2015, 14, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Al-Askar, A.A.; Arishi, A.A.; Behiry, S.I. Trichoderma hamatum Strain Th23 Promotes Tomato Growth and Induces Systemic Resistance against Tobacco Mosaic Virus. J. Fungi 2022, 8, 228. [Google Scholar] [CrossRef]
- Bakhtawar, F.; Wang, X.; Manan, A.; Iftikhar, Y.; Atta, S.; Bashir, M.A.; Mubeen, M.; Sajid, A.; Hannan, A.; Hashem, M.; et al. Biochemical characterization of citrus bent leaf viroid infecting citrus cultivars. J. King Saud Univ. Sci. 2022, 34, 101733. [Google Scholar] [CrossRef]
- Mohammadi, M.; Kazemi, H. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci. 2002, 162, 491–498. [Google Scholar] [CrossRef]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.Q.; Li, B.B.; Hong, Q.M.; Yan, Z.Y.; Yang, X.J.; Lu, K.C.; Chen, G.L.; Wang, L.; Chen, Y.H. A Glutathione Peroxidase Gene from Litopenaeus vannamei Is Involved in Oxidative Stress Responses and Pathogen Infection Resistance. Int. J. Mol. Sci. 2022, 23, 567. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhang, S.; Liu, X.; Shang, J.; Zhang, A.; Zhu, Z.; Zha, D. Chalcone synthase (CHS) family members analysis from eggplant (Solanum melongena L.) in the flavonoid biosynthetic pathway and expression patterns in response to heat stress. PLoS ONE 2020, 15, e0226537. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xu, X.; Kan, J.; Cheng, Z.M.; Chang, Y.; Lin, J.; Li, H. Genome-wide analysis of the C3H zinc finger family reveals its functions in salt stress responses of Pyrus betulaefolia. PeerJ 2020, 8, e9328. [Google Scholar] [CrossRef]
- Liu, H.; Huang, R.; Ma, J.; Sui, S.; Guo, Y.; Liu, D.; Li, Z.; Lin, Y.; Li, M. Two C3H Type Zinc Finger Protein Genes, CpCZF1 and CpCZF2, from Chimonanthus praecox Affect Stamen Development in Arabidopsis. Genes 2017, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Elsawy, M.M.; Ismail, I.A. Mechanism of resistance to Cucumber mosaic virus elicited by inoculation with Bacillus subtilis subsp. subtilis. Pest Manag. Sci. 2022, 78, 86–94. [Google Scholar] [CrossRef]
- Baebler, Š.; Stare, K.; Kovač, M.; Blejec, A.; Prezelj, N.; Stare, T.; Kogovšek, P.; Pompe-Novak, M.; Rosahl, S.; Ravnikar, M.; et al. Dynamics of responses in compatible potato—Potato virus y interaction are modulated by salicylic acid. PLoS ONE 2011, 6, e29009. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, F.; Tian, L.; Huang, M.; Deng, R.; Li, X.; Chen, W.; Wu, P.; Li, M.; Jiang, H.; et al. The Phenylalanine Ammonia Lyase Gene LjPAL1 Is Involved in Plant Defense Responses to Pathogens and Plays Diverse Roles in Lotus japonicus-Rhizobium Symbioses. Mol. Plant. Microbe. Interact. 2017, 30, 739–753. [Google Scholar] [CrossRef] [Green Version]
- Perrot, T.; Pauly, M.; Ram, V. Emerging Roles of β -Glucanases in Plant Development and Adaptative Responses. Plants 2022, 11, 1119. [Google Scholar] [CrossRef]
- Mélida, H.; Sopeña-Torres, S.; Bacete, L.; Garrido-Arandia, M.; Jordá, L.; López, G.; Muñoz-Barrios, A.; Pacios, L.F.; Molina, A. Non-branched β-1,3-glucan oligosaccharides trigger immune responses in Arabidopsis. Plant J. 2018, 93, 34–49. [Google Scholar] [CrossRef] [Green Version]
- Wanke, A.; Rovenich, H.; Schwanke, F.; Velte, S.; Becker, S.; Hehemann, J.H.; Wawra, S.; Zuccaro, A. Plant species-specific recognition of long and short β-1,3-linked glucans is mediated by different receptor systems. Plant J. 2020, 102, 1142–1156. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Hafez, E. Differential induction and suppression of the potato innate immune system in response to Alfalfa mosaic virus infection. Physiol. Mol. Plant Pathol. 2020, 110, 101485. [Google Scholar] [CrossRef]
- Beffa, R.S.; Hofer, R.M.; Thomas, M.; Meins, F.J. Decreased Susceptibility to Viral Disease of [beta]-1,3-Glucanase-Deficient Plants Generated by Antisense Transformation. Plant Cell 1996, 8, 1001–1011. [Google Scholar] [CrossRef]
- Iglesias, V.A.; Meins, F.; Meins, F., Jr. Movement of plant viruses is delayed in a β-1, 3-glucanase-deficient mutant showing a reduced plasmodesmatal size exclusion limit and enhanced callose deposition. Plant J. 2000, 21, 157–166. [Google Scholar] [CrossRef]
- Liu, H.; Carvalhais, L.C.; Kazan, K.; Schenk, P.M. Development of marker genes for jasmonic acid signaling in shoots and roots of wheat. Plant Signal. Behav. 2016, 11, e1176654. [Google Scholar] [CrossRef] [PubMed]
- Mathew, G.M.; Madhavan, A.; Arun, K.B.; Sindhu, R.; Binod, P.; Singhania, R.R.; Sukumaran, R.K.; Pandey, A. Thermophilic Chitinases: Structural, Functional and Engineering Attributes for Industrial Applications. Appl. Biochem. Biotechnol. 2021, 193, 142–164. [Google Scholar] [CrossRef] [PubMed]
- Oka, K.; Kobayashi, M.; Mitsuhara, I.; Seo, S. Jasmonic acid negatively regulates resistance to Tobacco mosaic virus in tobacco. Plant Cell Physiol. 2013, 54, 1999–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Xi, D.-H.; Yuan, S.; Xu, F.; Zhang, D.-W.; Lin, H.-H. Salicylic acid and jasmonic acid are essential for systemic resistance against tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant Microbe Interact. 2014, 27, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current overview of allergens of plant pathogenesis related protein families. Sci. World J. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Agoramoorthy, G.; Chandrasekaran, M.; Venkatesalu, V.; Hsu, M.J. Antibacterial and antifungal activities of fatty acid methyl esters of the blind-your-eye mangrove from India. Brazilian J. Microbiol. 2007, 38, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Waheed, A.; Chohan, M.M.; Ahmed, D.; Ullah, N. The first report on the in vitro antimicrobial activities of extracts of leaves of Ehretia serrata. Saudi J. Biol. Sci. 2019, 26, 1253–1261. [Google Scholar] [CrossRef]
- Krishnan, K.; Mani, A.; Jasmine, S. Cytotoxic Activity of Bioactive Compound 1, 2- Benzene Dicarboxylic Acid, Mono 2- Ethylhexyl Ester Extracted from a Marine Derived Streptomyces sp. VITSJK8. Int. J. Mol. Cell. Med. 2014, 3, 246–254. [Google Scholar]
- Yogeswari, S.; Ramalakshmi, S.; Neelavathy, R.; Muthumary, J. Identification and comparative studies of different volatile fractions from Monochaetia kansensis by GCMS. Glob. J. Pharmacol. 2012, 6, 65–71. [Google Scholar]
- Ugbogu, E.A.; Akubugwo, I.E.; Ude, V.C.; Gilbert, J.; Ekeanyanwu, B. Toxicological Evaluation of Phytochemical Characterized Aqueous Extract of Wild Dried Lentinus squarrosulus (Mont.) Mushroom in Rats. Toxicol. Res. 2019, 35, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Adeyemo, O.M.; Onilude, A.A.; Babatola, L.J. Effect of production parameters and inhibitory activity of antimicrobial compounds produced by co-cultured strains of Streptomyces xinghaiensis-OY62 and S. rimosus-OG95. J. King Saud Univ. Sci. 2020, 32, 294–301. [Google Scholar] [CrossRef]
- Faridha Begum, I.; Mohankumar, R.; Jeevan, M.; Ramani, K. GC–MS analysis of bio-active molecules derived from Paracoccus pantotrophus FMR19 and the antimicrobial activity against bacterial pathogens and MDROs. Indian J. Microbiol. 2016, 56, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Ashour, M.L.; El-Readi, M.; Youns, M.; Mulyaningsih, S.; Sporer, F.; Efferth, T.; Wink, M. Chemical composition and biological activity of the essential oil obtained from Bupleurum marginatum (Apiaceae). J. Pharm. Pharmacol. 2009, 61, 1079–1087. [Google Scholar] [CrossRef]
- Oepen, K.; Özbek, H.; Schüffler, A.; Liermann, J.C.; Thines, E.; Schneider, D. Myristic acid inhibits the activity of the bacterial ABC transporter BmrA. Int. J. Mol. Sci. 2021, 22, 13565. [Google Scholar] [CrossRef]
- Ren, J.; Wang, J.; Karthikeyan, S.; Liu, H.; Cai, J. Natural anti-phytopathogenic fungi compound phenol, 2, 4-bis (1, 1-dimethylethyl) from Pseudomonas fluorescens TL-1. Indian J. Biochem. Biophys. 2019, 56, 162–168. [Google Scholar]
- Kemung, H.M.; Tan, L.T.H.; Chan, K.G.; Ser, H.L.; Law, J.W.F.; Lee, L.H.; Goh, B.H. Antioxidant Activities of Streptomyces sp. strain MUSC 14 from Mangrove Forest Soil in Malaysia. BioMed Res. Int. 2020, 2020, 6402607. [Google Scholar] [CrossRef] [Green Version]
- Ser, H.-L.; Palanisamy, U.D.; Yin, W.-F.; Abd Malek, S.N.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Presence of antioxidative agent, Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro- in newly isolated Streptomyces mangrovisoli sp. nov. Front. Microbiol. 2015, 6, 854. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Direction | Nucleotide Sequence (5′ to 3′) |
|---|---|---|---|
| Zucchini yellow mosaic virus-coat protein | ZYMV-CP | Forward | GGACAGTGCGACTATAGCTTCAA |
| Reverse | TTTAACCGCGAATTGCGTATC | ||
| 16S ribosomal RNA | 16S rRNA | Forward | AGAGTTTGATCCTGGCTCAG GGTTACCTTGTTACGACTT |
| Reverse | |||
| Pathogenesis related protein-1 | PR-1 | Forward | CCAAGACTATCTTGCGGTTC |
| Reverse | GAACCTAAGCCACGATACCA | ||
| Endoglucanase | PR-2 | Forward | TCAATTATCAAAACTTGTTC |
| Reverse | AACCGGTCTCGGATACAAC | ||
| Chitinase | PR-3 | Forward | GGAGGAGTTCTTCAACGGCA |
| Reverse | ACGATTGGAGGGCTTCAAGG | ||
| Phenylalanine Ammonia-Lyase | PAL | Forward | ATGGAGGCAACTTCCAAGGA |
| Reverse | CCATGGCAATCTCAGCACCT | ||
| Chalcone Synthase | CHS | Forward | CACCGTGGAGGAGTATCGTAAGGC |
| Reverse | TGATCAACACAGTTGGAAGGCG | ||
| p-coumarate 3-hydroxylase | C3H | Forward | TTGGTGGCTACGACATTCCTAAGG |
| Reverse | GGTCTGAACTCCAATGGGTTATTCC | ||
| Elongation factor 1-alpha | EF1a | Forward | ATTCGAGAAGGAAGCTGCTG |
| Reverse | TTGGTGGTCTAAACTTCCAC |
| Treatment | DAS-ELISA Values * | Fresh Weight (g) | Dry Weight (g) | Total Soluble Carbohydrates mg/g DW | Total Soluble Proteins mg/g DW |
|---|---|---|---|---|---|
| Mock | 0.09 ± 0.02 d | 8.09 ± 0.52 a | 0.975 ± 0.25 a | 11.2 ± 0.6 a | 2.5 ± 0.2 a |
| Infected | 1.69 ± 0.06 a | 5.78 ± 0.45 d | 0.630 ± 0.23 d | 0.7 ± 0.02 c | 1.9 ± 0.01 b |
| Protective | 0.34 ± 0.03 c | 6.64 ± 0.36 b | 0.863 ± 0.24 b | 1.8 ± 0.1 b | 2.3 ± 0.4 a |
| Curative | 0.43 ± 0.03 b | 6.42 ± 0.39 c | 0.775 ± 0.21 c | 1.5 ± 0.1 d | 2.1 ± 0.1 ab |
| Peak No | Retention Time (RT) | Compound Name | Area | Molecular Formula | Molecular Weight |
|---|---|---|---|---|---|
| 1 | 3.43 | Propanoic acid, 2-oxo-, ethyl ester (ethyl pyruvate) | 1.143.99 | C5H8O3 | 116 |
| 2 | 11.03 | 1-Tetradecene | 253.40 | C14H28 | 196 |
| 3 | 11.09 | Hexadecane | 460.90 | C16H34 | 226.44 |
| 4 | 11.96 | Nonane, 1-iodo- | 245.18 | C9H19I | 254 |
| 5 | 12.18 | Phenol, 2,4-bis(1,1-dimethylethyl)- | 1.478.91 | C14H22O | 206 |
| 6 | 12.74 | 1-Hexadecene | 154.66 | C16H32 | 224.42 |
| 7 | 12.79 | Tridecane | 318.84 | CH3(CH2)11CH3 | 184.37 |
| 8 | 13.69 | Nonane, 5-(2-methylpropyl)- | 205.97 | C13H28 | 184 |
| 9 | 14.06 | Tetradecanoic acid | 803.98 | CH3(CH2)12COOH | 228.37 |
| 10 | 14.75 | Pyrrolo [1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)- | 214.59 | C11H18N2O2 | 210.27 |
| 11 | 15.43 | 1-Butanamine, N-(1-propylbutylidene)- | 610.87 | C11H23N | 169.31 |
| 12 | 15.48 | Pentadecanoic acid | 318.68 | C15H30O2 | 242.40 |
| 13 | 15.54 | L-Proline, N-valeryl-, heptadecyl ester | 994.52 | C27H51NO3 | 437.7 |
| 14 | 23.40 | 1,2-Benzenedicarboxylic acid, mono(2-ethylhexyl) ester | 31.739.38 | C16H22O4 | 278.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelkhalek, A.; Al-Askar, A.A.; Elbeaino, T.; Moawad, H.; El-Gendi, H. Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants. Biology 2022, 11, 1150. https://doi.org/10.3390/biology11081150
Abdelkhalek A, Al-Askar AA, Elbeaino T, Moawad H, El-Gendi H. Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants. Biology. 2022; 11(8):1150. https://doi.org/10.3390/biology11081150
Chicago/Turabian StyleAbdelkhalek, Ahmed, Abdulaziz A. Al-Askar, Toufic Elbeaino, Hassan Moawad, and Hamada El-Gendi. 2022. "Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants" Biology 11, no. 8: 1150. https://doi.org/10.3390/biology11081150
APA StyleAbdelkhalek, A., Al-Askar, A. A., Elbeaino, T., Moawad, H., & El-Gendi, H. (2022). Protective and Curative Activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus Infestation in Squash Plants. Biology, 11(8), 1150. https://doi.org/10.3390/biology11081150